
Abstract
Big science initiatives are trying to reconstruct and model the brain by attempting to simulate brain tissue at larger scales and with increasingly more biological detail than previously thought possible. The exponential growth of parallel computer performance has been supporting these developments, and at the same time maintainers of neuroscientific simulation code have strived to optimally and efficiently exploit new hardware features. Current state-of-the-art software for the simulation of biological networks has so far been developed using performance engineering practices, but a thorough analysis and modeling of the computational and performance characteristics, especially in the case of morphologically detailed neuron simulations, is lacking. Other computational sciences have successfully used analytic performance engineering, which is based on “white-box,” that is, first-principles performance models, to gain insight on the computational properties of simulation kernels, aid developers in performance optimizations and eventually drive codesign efforts, but to our knowledge a model-based performance analysis of neuron simulations has not yet been conducted. We present a detailed study of the shared-memory performance of morphologically detailed neuron simulations based on the Execution-Cache-Memory performance model. We demonstrate that this model can deliver accurate predictions of the runtime of almost all the kernels that constitute the neuron models under investigation. The gained insight is used to identify the main governing mechanisms underlying performance bottlenecks in the simulation. The implications of this analysis on the optimization of neural simulation software and eventually codesign of future hardware architectures are discussed. In this sense, our work represents a valuable conceptual and quantitative contribution to understanding the performance properties of biological networks simulations.
Keywords
1. Introduction and related work
1.1. Neuron simulations
Understanding the biological and theoretical principles underlying the brain’s physiological and cognitive functions is a great challenge for modern science. Exploiting the greater availability of data and resources, new computational approaches based on mathematical modeling and simulations have been developed to bridge the gap between the observed structural and functional complexity of the brain and the rather sparse experimental data, such as the works of Izhikevich and Edelman (2008), Potjans and Diesmann (2012), Markram et al. (2015), and Schuecker et al. (2017).
Simulations of biological neurons are characterized by demanding performance and memory requirements: a neuron can have up to 10,000 connections and must track separate states and events for each one; the model for a single neuron can be very detailed itself and contain up to 20,000 differential equations; neurons are very dense, and a small piece of tissue of roughly 1 mm3 can contain up to 100,000 neurons; finally, very fast axonal connections and current transients can limit the simulation timestep to 0.1 ms or even lower. Therefore, developers have gone to great lengths optimizing the memory requirements of the connectivity infrastructure in Jordan et al. (2018), the efficiency of the parallel communication algorithm in Hines et al. (2011) and Ananthanarayanan and Modha (2007), the scalability of data distribution in Kozloski and Wagner (2011) and even the parallel assembly of the neural network in Ippen et al. (2017). While these efforts improve the performance of the distributed simulations, little is still known about the intrinsic single-core and shared-memory performance properties of neuron simulations. On the other hand, the work of Zenke and Gerstner (2014) studied the performance of shared-memory simulations of biological neurons. However, their analysis is mainly based on empirical performance analysis and is centered on current-based point neuron simulations, a formalism that discards information about a neuron’s arborization.
The assumptions underlying brain simulations are very diverse, leading to a wide range of models across several orders of magnitude of spatial and time scales and thus to a complex landscape of simulation strategies, as summarized in the reviews by Brette et al. (2007) and Tikidji-Hamburyan et al. (2017). In this work, we focus on the simulation of morphologically detailed neurons based on the popular neuroscientific software NEURON presented in Carnevale and Hines (2006), which implements a modeling paradigm that includes details about a neuron’s individual morphology as well as its connectivity and allows to easily introduce custom models in the system. Our purpose is to extract the fundamental computational properties of the simulations of detailed biological networks and understand their relationship with modern microprocessor architectures.
1.2. The need for analytic performance modeling
An analytic performance model is a simplified description of the interactions between software and hardware together with a recipe for generating predictions of execution time. Such a model must be simple to be tractable but also elaborate enough to produce useful predictions.
Purely analytic (aka first-principles or white-box) models are based on known technical details of the hardware and some assumptions about how the software executes. The textbook example of a white-box model is the Roofline model by Williams et al. (2009) for loop performance prediction. The accuracy of such predictions depends crucially on the reliability of low-level details. A lack of predictive power challenges the underlying assumptions and, once corrected, often leads to better insight.
Black-box models, on the other hand, are ideally unaware of code and hardware specifics; measured data are used to identify crucial influence factors for the metrics to be modeled (see e.g. Calotoiu et al., 2013). One can then predict properties of arbitrary code or play with parameters to explore design spaces. Black-box models have a wider range of applicability: Even if low-level hardware information is lacking, they still provide predictive power. Wrong predictions, however, may be rooted in inappropriate fitting procedures and do not directly lead to better insight.
In this work, we choose the analytic approach combined with some phenomenological input, which makes the model a gray-box model. Analytic modeling has several decisive advantages that make it more suitable for delivering the insight we are looking for. First, it allows for universality identification, which means that some behavior in hardware–software interaction is valid for a wide range of microarchitectures of some kind. Second, it enables the identification of governing mechanisms: Since the model pinpoints the actual performance bottlenecks in the hardware, classes of codes with similar behavior are readily identified. This insight directly leads to possible codesign approaches. And third, analytic models provide insight via model failure, as described above.
1.3. The ECM performance model for multicore processors
The Execution-Cache-Memory (ECM) model takes into account predictions of single-threaded in-core execution time and data transfers through the complete cache hierarchy for steady-state loops. These predictions can be put together in different ways, depending on the CPU architecture. The following steps must be taken to set up the model for a given sequential loop code on a given architecture: Calculate data transfer volumes per iteration across the memory hierarchy, that is, for all data paths. This requires knowledge about how the data flows through the system and how the caches are organized (i.e. inclusive vs. exclusive). It is simple to do for pure streaming kernels with only spatial data locality but could become involved for kernels with data reuse. See, for example, Hager et al. (2018). Using the known data path bandwidths

Here, Vi is the data volume transferred per iteration over data path i. Latency effects are ignored in this step, so these times are “optimistic.” For convenience, we use a compact notation for such predictions, for example:
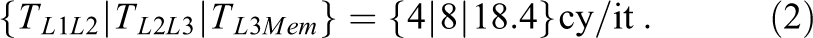
3. Set up a model for execution time of the loop code, assuming that all data reside in the L1 cache. This is based on the compiler-generated machine code. It may be as simple as assuming maximum throughput for all instructions (relying on the out-of-order execution capabilities of the core to ensure maximum overlap among successive loop body executions) or as complex as considering the full critical path (CP). Tools such as IACA, the Architecture Code Analyzer by Intel (2017),
1
can help with this task. It turns out that splitting the predicted execution time into data transfer cycles, specifically cycles in which loads retire (called “nonoverlapping time” 4. The calculated time contributions must now be combined according to a machine model appropriate for the architecture at hand. As shown by Hofmann et al. (2018), different CPUs can have different machine models, but the crucial feature of a machine is its ability to overlap data transfers in the memory hierarchy. On all recent Intel server microarchitectures, it turns out that the single-core model yields the best predictions if one assumes no temporal overlap of data transfers through the cache hierarchy and between the L1 cache and registers (i.e.
The necessary information about the processor to construct the ECM model, for example, data path bandwidths, the number, throughput, and latency of execution units, the organization of the caches, and so on can ideally be obtained from vendor documentation such as Intel (2018). If such information is unavailable or incomplete, as is usually the case for the overlapping characteristics of the memory hierarchy, microbenchmarks can be used. See Hofmann et al. (2019) for a full account of the necessary procedures.
For a data set in main memory on one core of an Intel server CPU with an inclusive cache hierarchy, the model thus predicts the following per-iteration loop runtime:
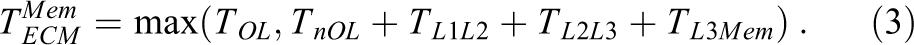
Here,
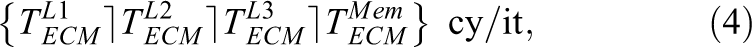
where
Scalability across cores is assumed to be perfect until a bandwidth bottleneck is hit. Since the memory interface is the only multicore bandwidth bottleneck on Intel processors, the predicted execution time is for n cores is
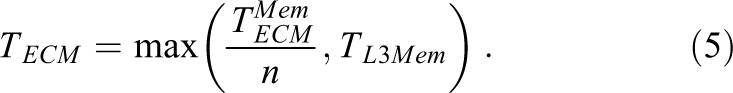
The bandwidth saturation point, that is, the number of cores required for saturation, is readily obtained from this expression:

A full account of the ECM model would exceed the scope of this article, so we refer to Stengel et al. (2015) and Hofmann et al. (2019) for a recent discussion. Note that IACA was used for the in-core analysis part of all the loops investigated in this article. The data transfer part was straightforward to set up since no significant temporal data reuse occurs. Difficulties encountered with the modeling procedures will be pointed out where relevant. An example of how the ECM model can be constructed for a simple streaming kernel will be given in Section 2.1.
Up to now, the ECM model has been shown to work well for standard multicore CPUs. Extension of the model to accelerators, specifically GPGPUs, is under investigation. The main problem with these kinds of architectures is that latency effects play a major role for single-core/thread execution, but they are ignored by the model in its current form.
1.4. Contributions and organization of this article
In this work, we make the following contributions: We demonstrate that the analytic ECM performance model can be applied successfully to nontrivial loop kernels with a wide range of different performance features. Although there are considerable error margins in some cases, a very good qualitative understanding can be achieved. We identify cases where the model needs corrections or cannot be applied sensibly: strong latency components in the data transfers and long CPs in the core execution. While latency-bound data access is beyond the applicability of the model in its current form, a long CP does not hinder the derivation of useful qualitative conclusions. We apply the ECM model for the first time to the Intel Skylake-X processor architecture, whose cache hierarchy is different from earlier Intel designs. We give clear guidelines for codesigning an “ideal” processor architecture for neuron simulations. In particular, we spot wide SIMD capabilities as a crucial ingredient in achieving memory bandwidth saturation. A low core count part with a high clock speed and wide SIMD units (such as AVX-512) will present the most cost-effective hardware platform. Cache size is inconsequential for most kernels.
This article is organized as follows: In section 2, we give details on the software and hardware environment under investigation, including preliminary performance observations. In section 3, we construct and validate ECM performance models for the important kernel classes in NEURON. In section 4, we summarize and discuss the findings in order to pinpoint the pivotal components of processor architectures in terms of neuron simulation performance and give an outlook to future work.
We provide a reproducibility appendix as a downloadable release file at Cremonesi et al. (2019), which should enable the interested reader to rerun our experiments and reproduce the relevant performance data.
2. Application and simulation environment
2.1. Target architectures and programming environment
We apply the ECM model introduced by Treibig and Hager (2010) and refined by Stengel et al. (2015) on two Intel processors with different micro-architectures: the Ivy Bridge (IVB) Intel(R) Xeon(R) E5-2660v2 and the Skylake (SKX) Intel(R) Xeon(R) Gold 6140 (with Sub-NUMA clustering turned off). The ECM model for the IVB architecture has been extensively studied by Hofmann et al. (2017) and Hammer et al. (2017). The ECM model for the SKX architecture has not been fully developed to date, but a preliminary formulation based on (5) that takes into account the victim cache architecture of the L3 was published in Hager et al. (2018). The heuristics governing cache replacement policies are not disclosed by Intel, but we have found that the following assumptions usually lead to good model predictions: Read traffic from main memory goes straight to L2. All evicted cache lines from L2, both clean and dirty, are moved to L3. The data path between the L2 and the L3 cache can be assumed to provide a bandwidth of 16 B/cy in both directions (i.e. full duplex).
The most relevant hardware features for the modeling of both architectures are presented in Tables 1 and 2. The Intel IACA tool was used for estimating in-core execution times of loop kernels. Although IACA supports both architectures, its support for CP prediction was recently dropped. The IACA outputs for all kernels are available in the reproducibility appendix.
Hardware characteristics of the target CPU architectures.
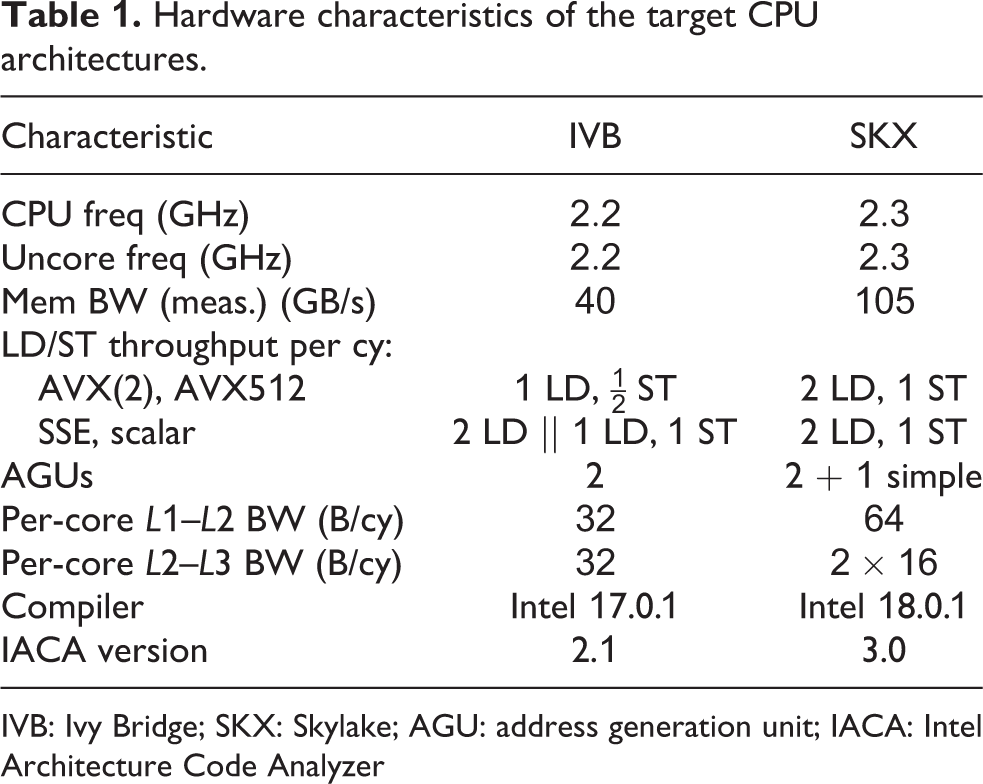
IVB: Ivy Bridge; SKX: Skylake; AGU: address generation unit; IACA: Intel Architecture Code Analyzer
Useful benchmark values (double precision).a
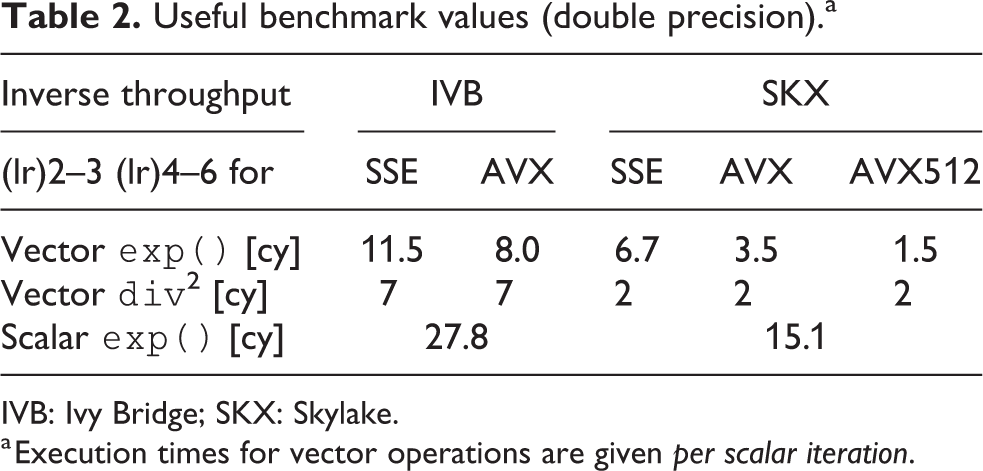
IVB: Ivy Bridge; SKX: Skylake.
a Execution times for vector operations are given per scalar iteration.
We illustrate the application of the ECM model to SKX with the STREAM triad kernel developed by McCalpin (1995):
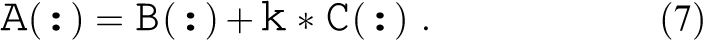
Considering only AVX vectorization as an example, this kernel has the following properties per scalar iteration: Inverse throughput prediction of Two loads and one store, so Due to the victim L3 cache, we have to distinguish in-memory and in-L3 data sets. L3: Memory: The transfer times between L2 and L3 are the same in this particular case because reads and writes to L3 can overlap: L3: Memory:
So the ECM model contributions for the STREAM triad kernel in (7) on SKX-AVX would be:
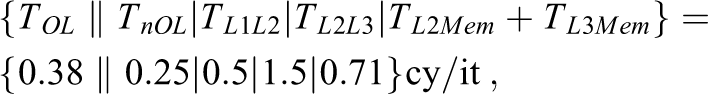
with corresponding predictions according to the nonoverlapping machine model of
To roughly compare the two architectures, a common approach is to use the peak performance as a metric, measured in single-precision or double-precision Gflop/s. The IVB chip supports AVX vectorization and can retire one multiply and one add instruction per cycle, while the SKX chip supports AVX512 vectorization and can retire two fused multiply-add instructions per cycle. This leads to the peak performance numbers as shown in Table 3. The naive Roofline model uses the peak performance of the chip as the core-bound limit, but often other limitations apply, such as the load or store throughput between registers and the L1 cache, or pipeline stalls due to a long CP or loop-carried dependencies. The ECM model takes this into account via the
Peak performance for the target architectures.

IVB: Ivy Bridge; SKX: Skylake.
On IVB, we used the Intel 17.0.1 compiler with options
To measure relevant performance metrics such as data transfer through the memory hierarchy, we used the
2.2. Simulation of morphologically detailed neurons
A common approach to modeling morphologically detailed neurons is the so-called conductance-based (COBA) compartmental model as formalized in the reviews by Brette et al. (2007), Bhalla (2012), and Gerstner et al. (2014). In this abstraction, the arborization of dendrites and axons is represented as a tree of sections, where a section corresponds to an unbranched portion of the neuron. Each section is then divided into compartments that represent discretization units for the numerical approximation. Quantities of interest such as membrane potential or channel gating variables are typically only well defined at compartment centers and branching points.
In the compartmental model, each compartment is considered analogous to an RC circuit where the resistance (or rather, the conductance) term can be nonlinearly dependent on the membrane potential itself. Due to their stability, implicit methods are a common choice for time integration of the differential equations arising from this representation, thus the solution of a linear system of equations is required at each timestep. In the presence of branching points, this leads to a quasi-tridiagonal system that can be solved in linear time using the algorithm proposed in Hines (1984).
In the COBA model, the membrane conductance is determined by aggregating several contributions from ion channels, which are special cross-membrane proteins that allow an ionic current to flow into or out of the cell. Thus in the COBA compartmental model, when using an implicit time integrator, three algorithmic phases are required to advance a neuron in time: first one must compute the contributions to the linear system (the ion channel and synapses currents); then one must solve the linear system; finally, one must update the states of individual ion channel and synapse instances based on the recently computed compartment potentials (see Figure 1). Note that in this model each ion channel or synapse type will have two kernels associated with it: a current kernel that defines how to compute the contributions to the linear system for that family of ion channels or synapses, and a state kernel that defines the numerical time integration of that ion channel’s or synapse’s state variables, typically based on the exponential Euler method (see Oh and French, 2006).
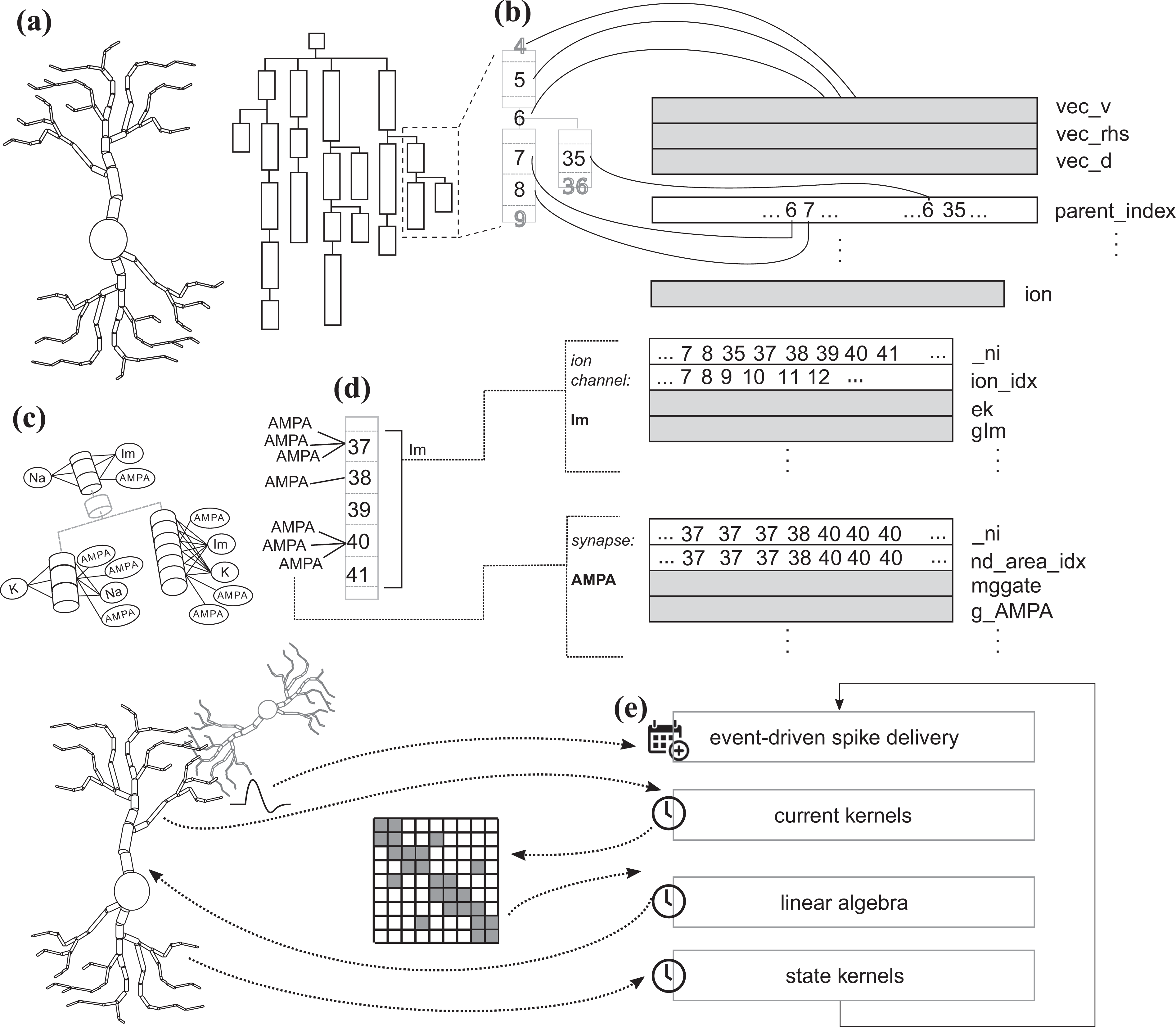
Neuron representation and data layout. (a) Neurons are represented as a tree of unbranched sections, where each section can be further split into compartments for numerical discretization. (b) Each compartment is numbered according to the schema in Hines (1984), and the tree structure is represented in memory by an array of
Neurons also have the ability to communicate with other neurons using synapses: points of contact between different neurons that are triggered when an action potential is elicited in the presynaptic cell and, at the onset of this event, determine a change in the membrane potential of the postsynaptic cell. Therefore, the simulation algorithm is composed of two parts: a clock-driven portion that advances the state of a neuron from a timestep to the next; and an event-driven part that is only executed when a synaptic event is received. Figure 1 presents a summary of the main algorithm phases and data layout.
The compartmental modeling of neurons using COBA formalism is implemented in the widely adopted software for neuroscientific simulations NEURON. The NEURON software is a long-lasting project that includes an interpreter for a custom scripting language (HOC), a domain specific language tool to expand the models of ion channels and synapses, a GUI and a domain specific language (NMODL) to expand the repertoire of available models. To reduce the complexity and concentrate on the fundamental computational properties of the simulation kernels, in this work we utilize instead CoreNEURON, a lean version of NEURON’s simulation engine based on the work by Kumbhar et al. (2016). CoreNEURON implements several optimizations over NEURON, including improved memory requirements and vectorization, at the cost of functionality. In particular, NEURON is usually still required to define a model and a simulation setup before CoreNEURON executes the simulation. The NEURON/CoreNEURON software allows neuroscientists to specify custom ion channel and synapse models using the domain specific language NMODL introduced in Hines and Carnevale (2000), which is then automatically translated into C code and compiled in a dynamic library.
The CoreNEURON data layout is shown in Figure 1. First, the neuron is modeled logically as a tree of unbranched sections, whose topology is represented by a vector of parent indices. Other relevant quantities such as the membrane potential and the tridiagonal sparse matrix are represented by double precision arrays with length equal to the number of compartments. More details about the matrix representation are given in Section 3.6. Additionally, ion channel-specific and synapse-specific quantities are held in separate data structures consisting of arrays of double precision values in Structure-of-Arrays layout (SoA), indices to the corresponding compartments and, if needed, pointers to other internal data structures.
2.3. Preliminary performance observations and motivation
Given that the simulation algorithm is composed of many phases with different characteristics, the first step in performance analysis is a search for the time-consuming hot spots. We created a data set consisting of 500 replicas of a Layer 5 pyramidal neuron from the rat’s somatosensory cortex (see Ramaswamy et al., 2015). This type of neuron was chosen because it contains all of the most common ion channel and synapse types in a reconstruction of the cortical microcircuit (see Markram et al., 2015), while the number of replicas was chosen to ensure with absolute certainty that the corresponding data set would not fit in the L3 cache. Although the number of neurons in a data set primarily affects its size, and thus the level of memory where the working set resides, network size effects can come into play, changing the frequency at which neurons exchange synaptic events, thus changing the relative weight of the spike delivery function. However, such effects are difficult to predict and depend on many external factors such as random input noise and connectivity among neurons, therefore we chose to neglect this direction of analysis and kept the number of neurons fixed. To account for different data set sizes, in the detailed analysis of individual ion channels and synapses, we also consider smaller data sets that fit in the L3 and L2 cache. In the shared-memory parallel execution, current and state kernels are usually dominant, representing roughly 80% of the total execution time, while the event-driven spike delivery and linear algebra kernels account for less than 10% each (see Figure 2(a)). In the serial execution, we observe that the relative importance of spike delivery increases slightly, however, the state and current kernels still dominate. This serial performance profile was also observed in Kumbhar et al. (2016) and is a distinctive feature of compartmental COBA models, whereas current-based point neuron models are typically dominated in serial execution by event-driven spike delivery and event bookkeeping, as shown in the work by Peyser and Schenck (2015). Unfortunately, these results are tightly linked to the benchmark setup, and it is unknown whether this is a general property of COBA models or not.
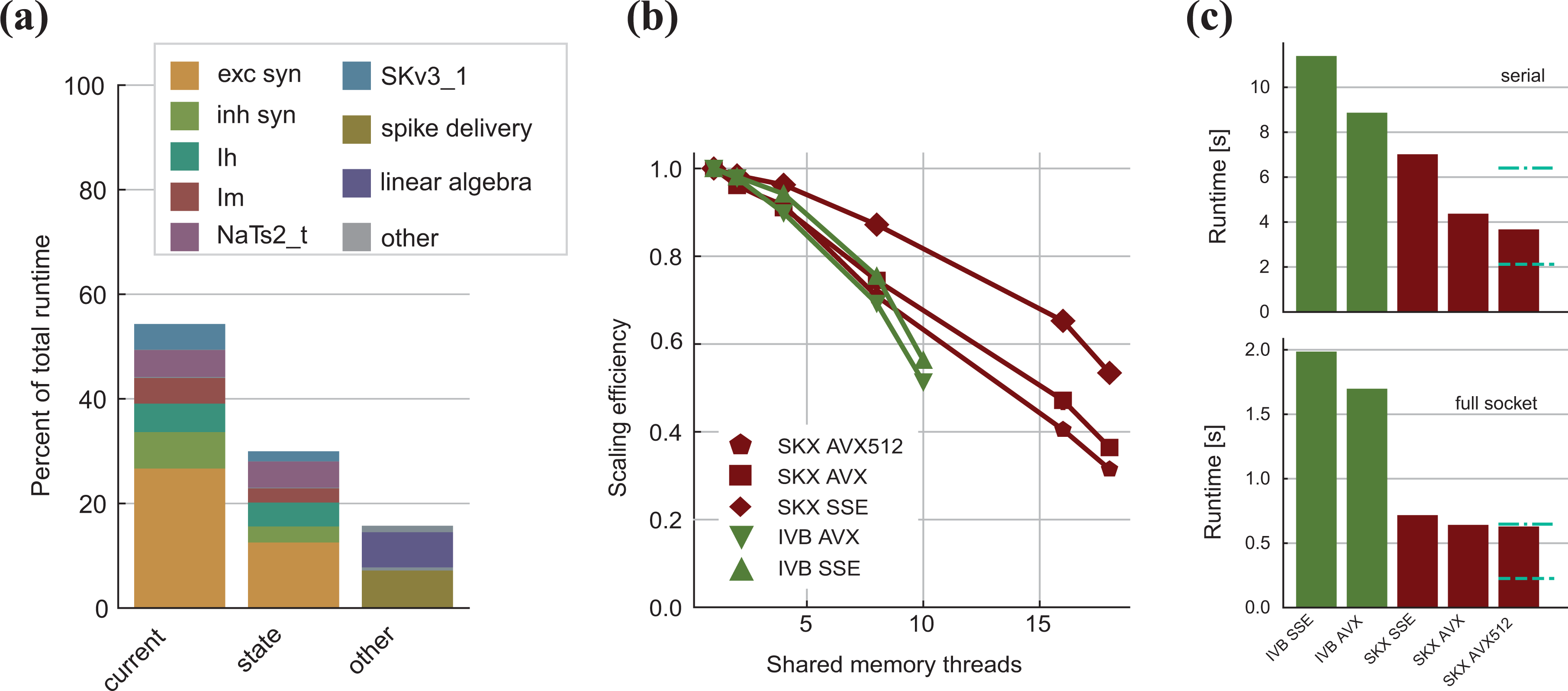
Measured performance and observations from benchmark. (a) Breakdown of the distribution of relative importance of the different kernels in the simulation of a full neuron for the SKX-AVX512 architecture using a full socket. Overhead from the rest of the execution is not shown. Linear algebra and spike delivery combined hardly exceed 10%. (b) Median strong scaling efficiency over 10 runs. Measurements exhibited little variability across different runs, so quantile error bars are not visible. Parallel efficiency degrades quickly, especially on SKX, and vectorization strengthens this negative effect. (c) Total runtime to simulate one neuron for one second in the serial case (top) and using the full socket (bottom). Overhead from the non-computational kernels is not considered. The dashed blue line represents the expected runtime if scaled perfectly with the architecture’s theoretical peak performance from IVB-AVX to SKX-AVX512, while the dash-dotted blue line marks the expected runtime if scaled perfectly with the ratio of measured memory bandwidths. We do not observe the ideal speedup, and in this article we employ performance modeling to explain the underlying reasons. IVB: Ivy Bridge; SKX: Skylake.
We have chosen two Intel architectures that were introduced about 5 years apart in order to be able to identify the speedup from architectural improvements. Judging by the peak performance numbers in Table 3, one would expect a per-socket speedup of about 7.5
One of the main in-core features of modern architectures is the possibility to expose data parallelism using vectorized (SIMD) instructions on wide registers. We investigated the benefits of vectorization at different levels of thread-parallelism. In the serial execution (see Figure 2(c)), we found that the Skylake architecture had in general better performance than Ivy Bridge, and that using wider registers improved the performance, even though the acceleration was not ideal (i.e. not in line with the larger register width). At full socket, we found that the difference between architectures was exacerbated, while we saw only minor improvements from vectorization (see Figure 2(c)). We also investigated the strong-scaling efficiency of the simulation code on different architectures (Figure 2(b)) and found that, as expected, the efficiency decreases as the level of parallelism grows. This indicates a tradeoff in terms of chip and software design: further analysis is required to understand whether it is worth investing in SIMD or shared-memory parallelism, optimize for instruction level parallelism, out of order execution or a combination of all of these.
We exploit performance modeling techniques in order to gain insight into the interaction between the CoreNEURON simulation code and modern hardware architectures. This will allow us to answer the open performance questions above as well as to generalize to different architectures for future codesign efforts.
3. Performance modeling of detailed simulations of neurons
3.1. Ion channel current kernels
Ion channel current kernels are used in the simulation algorithm to update the matrix representing the voltage equation by computing contributions from the ionic current of different chemical species. We consider in this work four ion channel types that are among the most representative:


Current kernels are typically characterized by two main features: low arithmetic intensity and scattered loads/stores. The latter can present a modeling problem, but in practice we can obtain good accuracy using a few heuristics based on domain-specific knowledge. In particular, as a first approximation, one can treat the indices in
Combining the data volume estimates with in-core predictions from IACA (using the full throughput assumption), we can generate the ECM model predictions in cycles per scalar iteration as shown in Table 4. The compiler is able to employ scatter/gather instructions for this kernel on SKX (these are not supported on IVB). As expected, the model predicts that the performance of this strongly data-bound kernel will degrade as the data resides farther from the core. Vectorization is not beneficial at all except for AVX512 with data in L1, which can be attributed to the required scalar load instructions when gather/scatter instructions are missing. To validate the predictions, we designed a serial benchmark that allowed fine-grained control over the data set size by removing all ion channels and synapses except
ECM model and serial measurements per scalar iteration (cy/it) for the
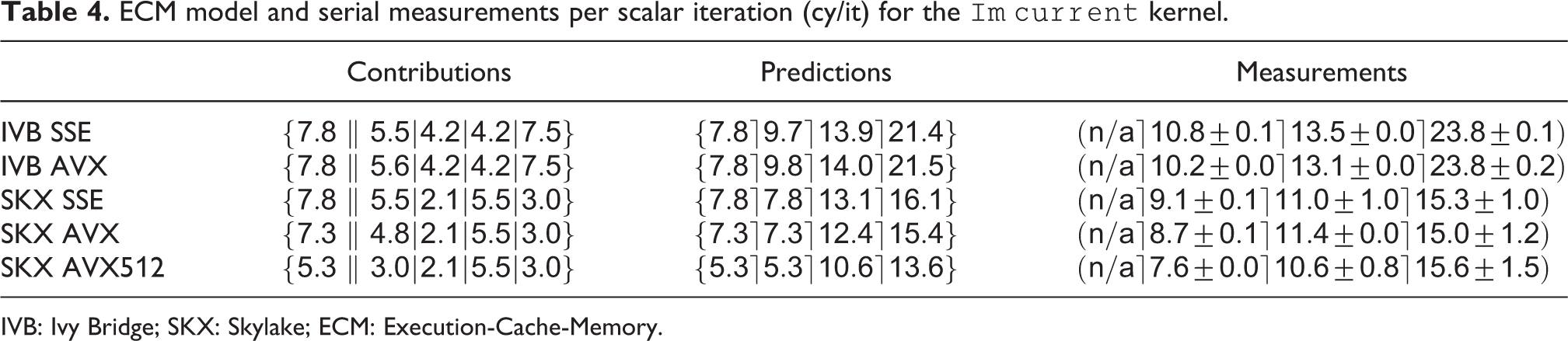
IVB: Ivy Bridge; SKX: Skylake; ECM: Execution-Cache-Memory.
On IVB, the measurements remained within 10% of the predictions for all levels of the memory hierarchy, while on SKX, the ECM predictions were a little more off, especially for data in the cache. This might be caused by our simplifying model assumptions about the data transfers between L2 and L3, for which no official documentation exists. Still the ECM model gave accurate predictions in almost all of our benchmarks and provided insight into the computational properties of this kernel.
We conclude that the
3.2. Synaptic current kernels
Synapses are arguably the pivotal component of neuron simulations. Synaptic current kernels are particularly important for performance as shown in Figure 2, and pose a modeling challenge because of their complex chain of intra-loop dependencies, memory accesses and presence of transcendental functions. There are two types of synapses in this data set: excitatory AMPA/NMDA synapses and inhibitory GABA-A/B synapses. As an example, the source code for the excitatory AMPA/NMDA synapse current is shown in Listing 2. The expensive exponentials and divides in this code are balanced by large data requirements. The kernel reads one element each from eight double and two integer arrays, and writes one element each to nine double arrays, which would amount to a traffic of 216 B per iteration. However, as shown in Figure 1, the typical structure of the
Excitatory synapse current kernel.


ECM model and serial measurements per scalar iteration (cy) for the excitatory synapse current kernel.
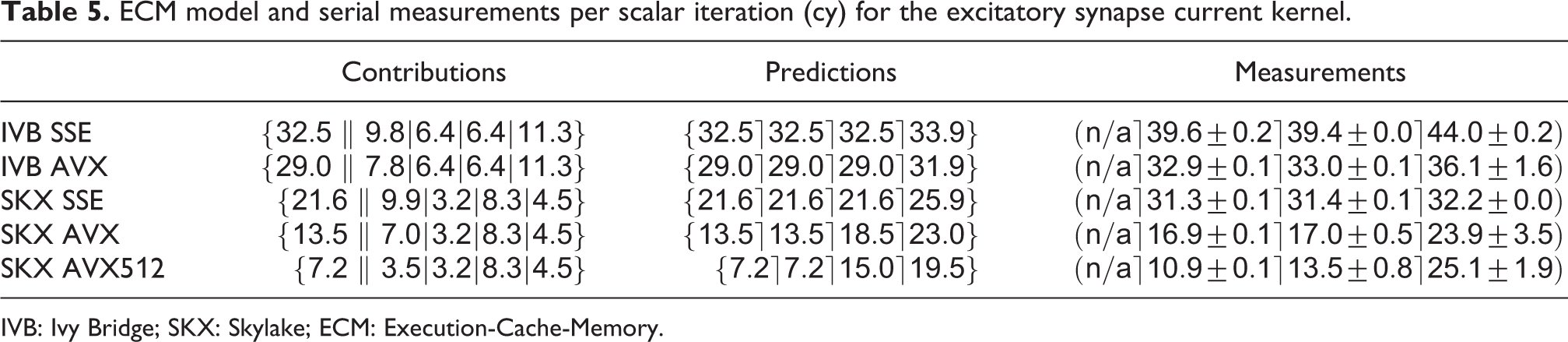
IVB: Ivy Bridge; SKX: Skylake; ECM: Execution-Cache-Memory.
The analysis reveals a complex situation. Both code versions on IVB and the SSE code on SKX are predicted to be core bound as long as the data fits into any cache. The AVX and AVX512 code on SKX, however, become data bound already in the L3 cache.
Again we used a benchmark data set containing only synapses to validate the model, with a size of roughly 80 kB and 500 kB for the L2 benchmarks and 1.5 MB and 11 MB for the L3 benchmark on the IVB and SKX architectures, respectively. On both CPUs, the model predictions are optimistic compared to measurements by a 10–50% margin. Interestingly, within each architecture, the model becomes more accurate as the SIMD width increases. Even though the predictions are not all within a small accuracy window, the model still allows us to correctly categorize the relevant bottlenecks and is especially effective in capturing the fact that on SKX with AVX512 the kernel is rather strongly data bound. Given the complex inter-dependencies between operations in the kernel, we speculate that a CP might be invalidating the full-throughput assumption of the ECM model, although this would not be sufficient to explain why the DRAM measurements are larger than the L2 and L3 measurements.
As a result from the analysis we conclude that, for an in-memory data set, the performance of the serial excitatory synapse current kernel would improve significantly only if in-core execution and data transfers were enhanced at the same time. Still the model predicts bandwidth saturation for all code variants, once run in parallel, at 4–6 cores.
3.3. Ion channels state kernels
During the execution of a state kernel, the internal state variables of an instance of an ion channel or a synapse are integrated in time and advanced to the next timestep. Figure 2(a) shows that state kernels represent a significant portion of the overall runtime, although their relative importance is largest in the single-thread execution and decreases with shared-memory parallelism.
State kernels are characterized by a very large overlapping contribution
Ih state kernel.


Again combining the IACA prediction with measured throughput data for
ECM model and serial measurements per scalar iteration (cy) for Ih state kernel.
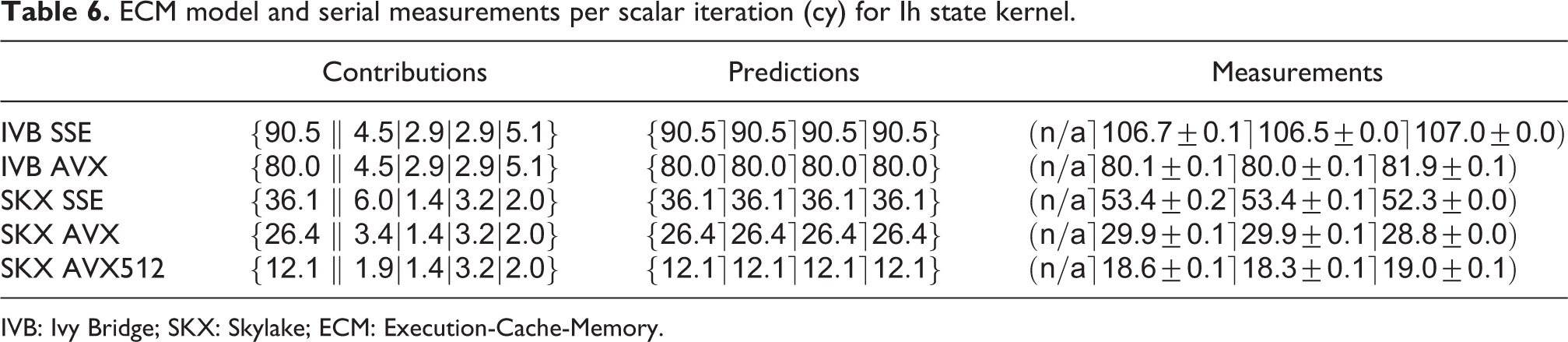
IVB: Ivy Bridge; SKX: Skylake; ECM: Execution-Cache-Memory.
Both architectures show only moderate speedup from SSE to AVX (13% on IVB and 37% on SKX, respectively). On IVB, this can be partly attributed to the mere 44% speedup for the
AVX512, on the other hand, exhibits a large speedup that cannot be explained by the above analysis. Inspection of the assembly code reveals that the compiler did not generate any divide instructions at all. Instead, it uses
We validated our predictions with data set sizes of 124 kB and 500 kB for the L2 benchmarks on IVB and SKX respectively, and a data set size of 5 MB for the L3 benchmarks on both architectures. Except for the AVX kernels, for which the accuracy is more than satisfying, the predictions are optimistic by between 15% and 35%. It must be stressed that when a loop is strongly core bound and has a long CP, the automatic out-of-order execution engine in the hardware may have a hard time overlapping successive loop iterations. Since the ECM model has no concept of this issue, predictions may be qualitative.
Despite all inaccuracies, the conclusion from the analysis is clear: Faster exponential functions, wider SIMD execution for divide instructions, and a higher clock frequency would immediately (and proportionally) boost the performance of the serial
3.4. Synaptic state kernels
Synapse state kernels have computational properties similar to ion channel state kernels, that is, a dominating in-core overlapping contribution due to exponentials and divides, coupled with low data requirements. As an example, we show the code for the excitatory AMPA/NMDA synapse in Listing 4. This kernel reads one element each from four double arrays and updates one element each from four other double arrays, thus totaling 96 B of data volume per iteration. The ECM predictions per scalar iteration are listed in Table 7. An important observation to be made here is that using the
Excitatory synapse state kernel.


ECM model and serial measurements per scalar iteration (cy) for the excitatory synapse state kernel.
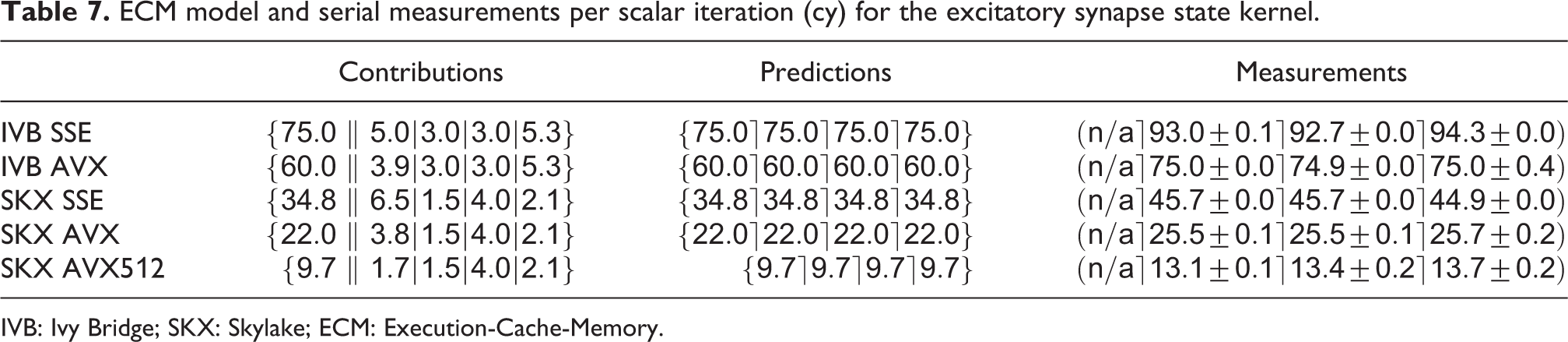
IVB: Ivy Bridge; SKX: Skylake; ECM: Execution-Cache-Memory.
3.5. Validation for all state and current kernels
To assess the validity of our performance and conclusions about bandwidth saturation on a real-world use case, we designed a representative data set based on the
Tables 8 and 9 show the predicted and measured runtimes of current and state kernels for the two architectures, all vectorization levels and serial versus full-socket parallel execution, while Figure 3 presents the performance scaling of these kernels across the cores of a chip. Interestingly, we observe a significant variability in serial runtime across kernels, with
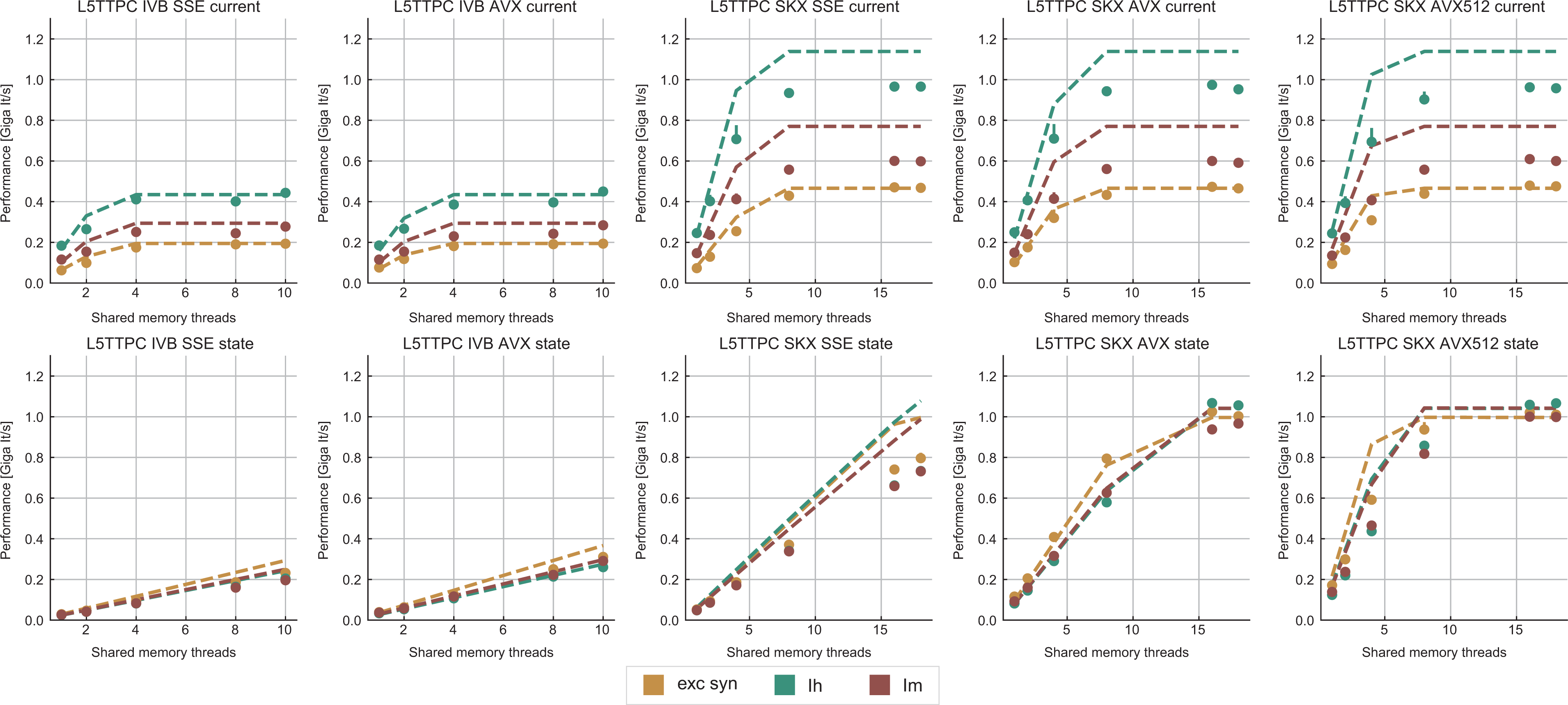
Performance predictions (dashed lines) and measurements (solid markers) for selected ion channel and state kernels, on all architectures and vectorization levels. The unit of measure for performance is Giga scalar iterations per wall clock second, denoted Giga It/s. Measurements points are computed as the median and error bars represent the 25- and 75-percentile out of 10 runs. Due to automatic clock frequency scaling, the performance predictions of each kernel were scaled by the kernel’s average clock frequency to preserve consistency with the measurements. Current kernels show a typical saturation behavior at low thread counts while state kernels either do not saturate at all (IVB), saturate at large thread counts (SKX-SSE, SKX-AVX) or saturate at moderate to low thread counts (SKX-AVX512). IVB: Ivy Bridge; SKX: Skylake.
Runtime for all current and state kernels on the IVB architecture (in-memory working set).a
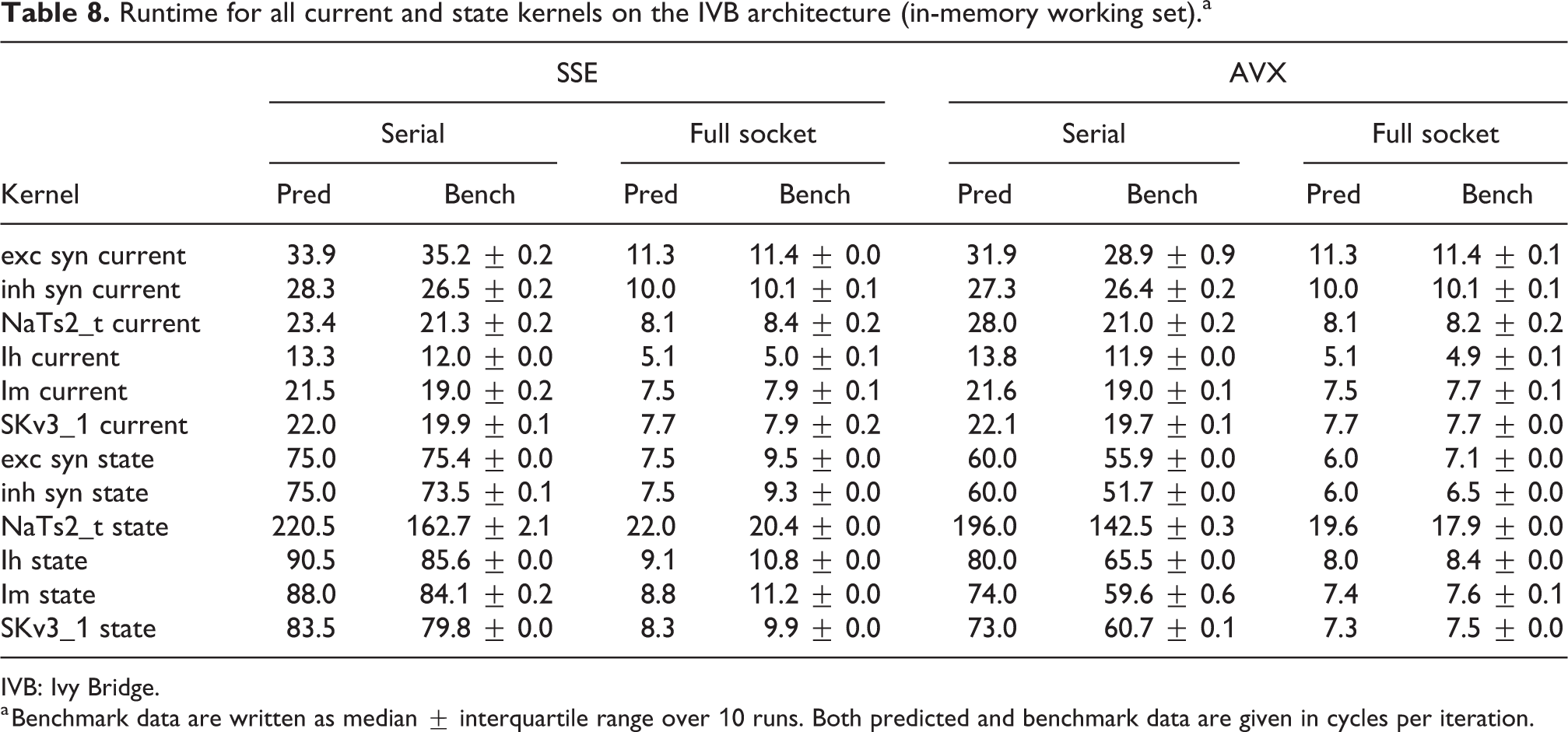
IVB: Ivy Bridge.
a Benchmark data are written as median
Runtime for all current and state kernels on the SKX architecture (in-memory working set).a
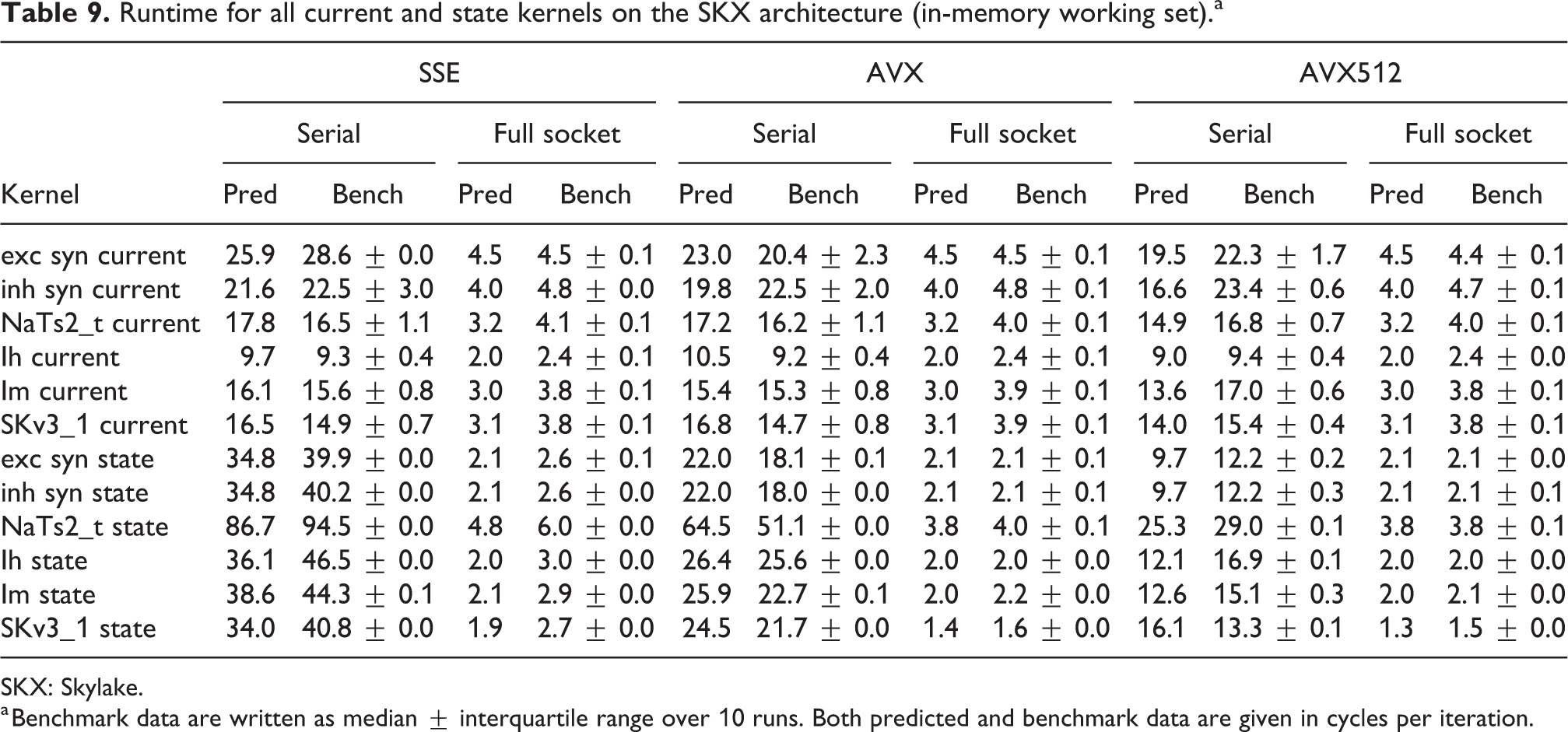
SKX: Skylake.
a Benchmark data are written as median
In the rest of this section, we address some of the largest deviations between measurements and predictions by providing a tentative explanation for the failure of our performance model. As stated in the state kernel Sections 3.3 and 3.4, a long CP in the loop kernel code could be weakening the accuracy of our predictions due to a failure of the full throughput assumption. We believe that, in order to improve our predictions, a cycle-accurate simulation of the execution and in particular of the Out-of-Order (OoO) engine would be needed, thus invalidating our requirement for a simple analytical model. At large thread counts the predictions for current kernels are always within a reasonable error bound, while those for state kernels can be off by as much as 30%. The state kernels’ performance is often in a transitional phase between saturation and core-boundedness even at large thread counts, where the ECM model in the form we use it here is known to perform poorly as shown in Stengel et al. (2015). We do not plan to employ the adaptive latency penalty method as described in Hofmann et al. (2018) to correct for this discrepancy, because it is not only a modification of the machine model but also requires a parameter fit for every individual loop kernel. We believe that this is an undesirable trait in an analytic model.
3.6. Special kernels: Linear algebra
The most common approach for time integration of morphologically detailed neurons is to use an implicit method (typically backward-Euler or Crank-Nicolson) in order to take advantage of its stability properties for stiff problems. In Hines (1984), a linear-complexity algorithm based on Thomas (1949) was introduced to solve the quasi-tridiagonal system arising from the branched morphologies of neurons. This algorithm is based on a sparse representation of the matrix using three arrays of values (
Linear algebra kernel.


To construct a performance model for this kernel, we must tackle a few challenges: Indirect accesses make it difficult to estimate the data traffic, and dependencies between loop iterations could break the full-throughput hypothesis. Moreover, a yet-unpublished optimized variant of the algorithm proposed in Hines (1984) that exploits a permutation of node indices to maximize data locality is executed by default by the simulation engine.
3
For reasons of brevity of exposition, we restrict our analysis to this optimized variant of the solver. Additionally, we will ignore the
In order to give a runtime estimate, we examine two corner-case scenarios. The first, optimistic scenario assumes that indirect accesses can exploit spatial data locality in caches and thus do not generate any additional memory traffic. The combined data traffic requirements of triangularization and back-substitution then amount to reading one element each from four double arrays and two integer arrays, and writing one element each to three double arrays, that is, 88 B per iteration. Considering the opposite extreme, it might happen that at every branching point the value of
Even though the dependencies between loop iterations could potentially break the full-throughput hypothesis, considering that compartment indices are by default internally rearranged to optimize data locality we still use the full throughput as a basis for our predictions. It should be noted that indirect addressing and potential loop dependencies hinder vectorization. IACA reports that the combined inverse throughput of triangularization and back substitution amounts to 28 cy/it for IVB and 8.12 cy/it for SKX, while
ECM model and serial measurements per scalar iteration (cy) for the linear algebra kernel.a
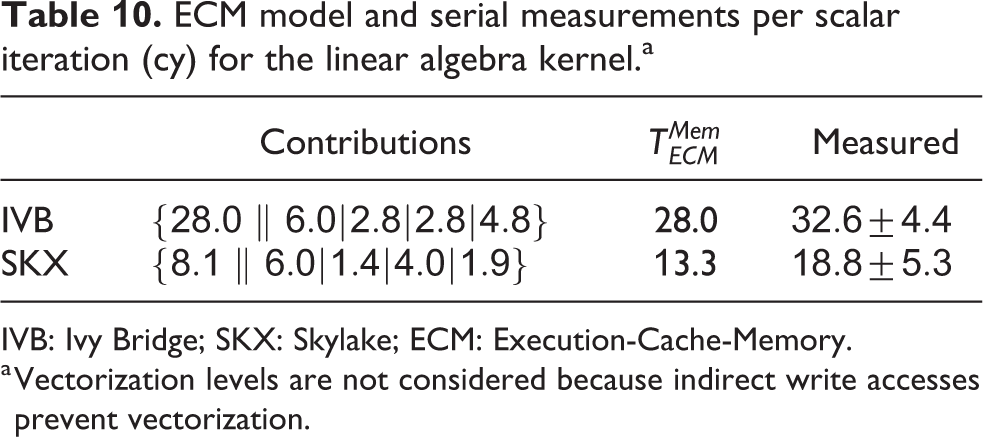
IVB: Ivy Bridge; SKX: Skylake; ECM: Execution-Cache-Memory.
a Vectorization levels are not considered because indirect write accesses prevent vectorization.
We measured the performance of the linear algebra kernel on a specially designed data set with 15 K neurons and neither ion channels nor synapses. Since the memory requirement of this kernel is only of 36 B per compartment, the number of neurons can have an important effect on performance as the data set for even a few hundred neurons could easily fit in a cache. This would lead to data reuse that, while beneficial to performance, would invalidate our model. Thus the number of neurons was chosen large enough to ensure that the only data locality effects are intrinsic to the algorithm and not a consequence of a small data set. Our predictions based on the full-throughput hypothesis are validated by measurements of both the performance (see Figure 4) and the memory traffic (last row in Table 11). This kernel highlights very strongly an important difference between the two architectures: SKX has a much better divide unit, which is able to deliver one result every four cycles, whereas IVB’s divider needs 14. This ratio is almost exactly reflected in the
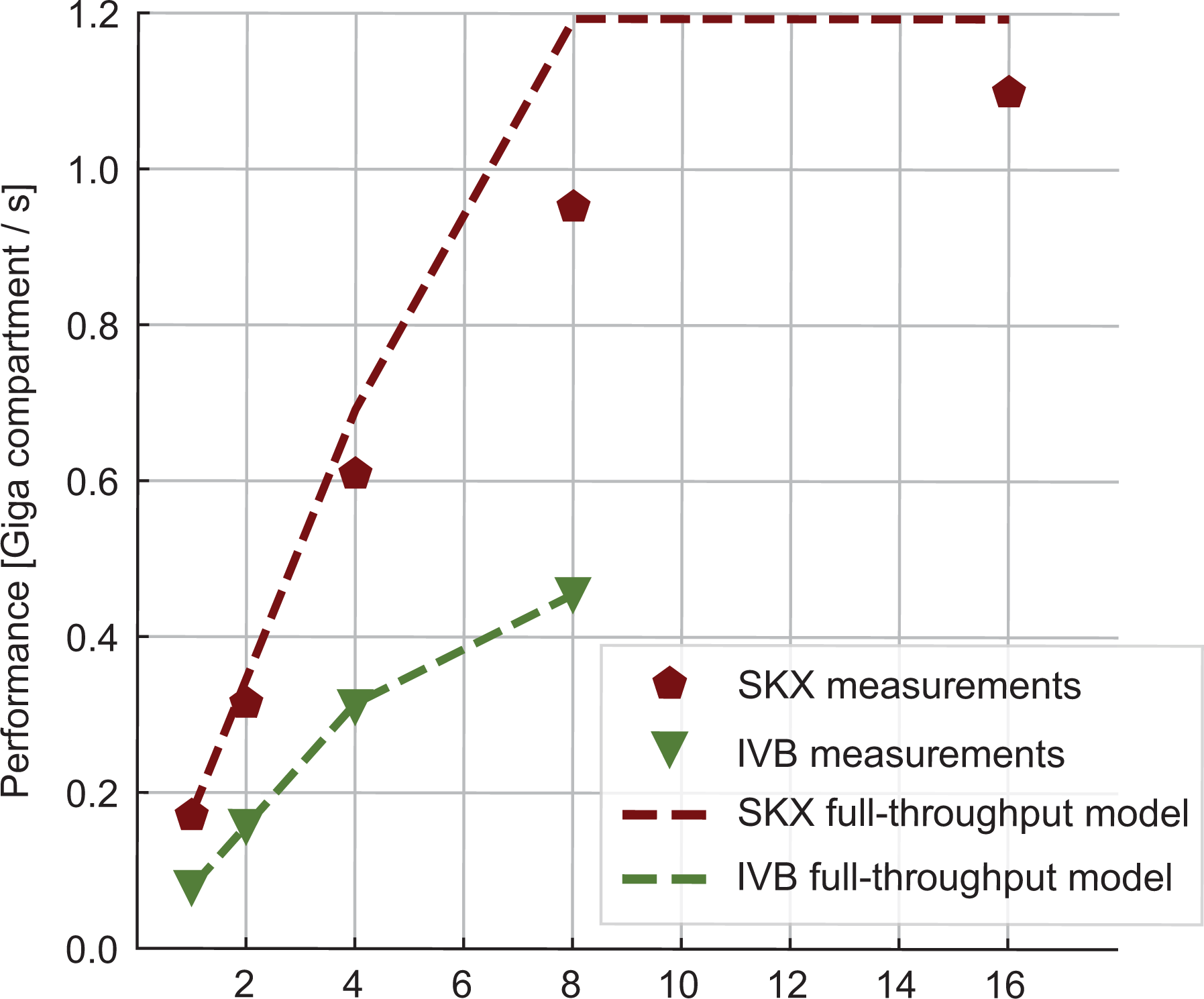
Measured performance (markers) and predictions (lines) for the linear algebra kernel in Giga-compartments per second. Dashed lines represent the model predictions in the optimistic full-throughput scenario.
Predicted (pred) and measured (meas) data volume per iteration from main memory.a
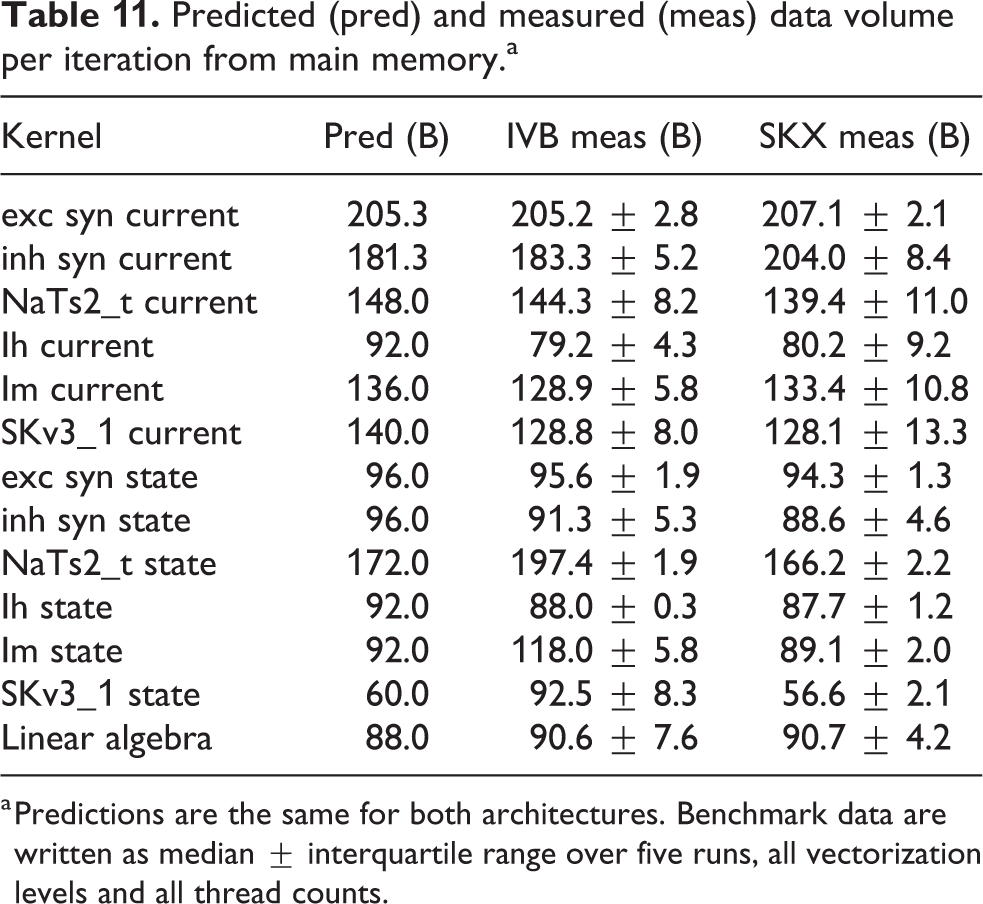
a Predictions are the same for both architectures. Benchmark data are written as median
We remark that it remains unclear whether the node permutation optimization is applicable in all cases or suffers from some constraints, and that our full-throughput predictions heavily rely on it. Therefore it may happen that, in some cases where it is impossible to reorder the nodes effectively, our predictions would only provide an optimistic upper bound on performance.
3.7. Special kernels: Spike delivery
Accounting for network connectivity and event-driven spike exchange between neurons is, in terms of algorithm design, the most distinguishing feature of neural tissue simulations. In terms of performance, however, spike delivery plays a marginal role in the simulation of morphologically detailed neurons, rarely exceeding 10% of the total runtime (see Figure 2(a)).
The source code for the spike delivery kernel of AMPA/NMDA excitatory synapses is shown in Listing 6. For benchmarking purposes, we executed this kernel as the body of a loop iterating over a vector of spike events, which was previously populated by popping a priority queue. 4 This only represents a small deviation from the original implementation in CoreNEURON, where the kernel is directly called at every pop of the priority queue. However, it was necessary to implement this in order to separate the performance of the kernels from the performance of the queue operations.
Event-driven spike delivery kernel.

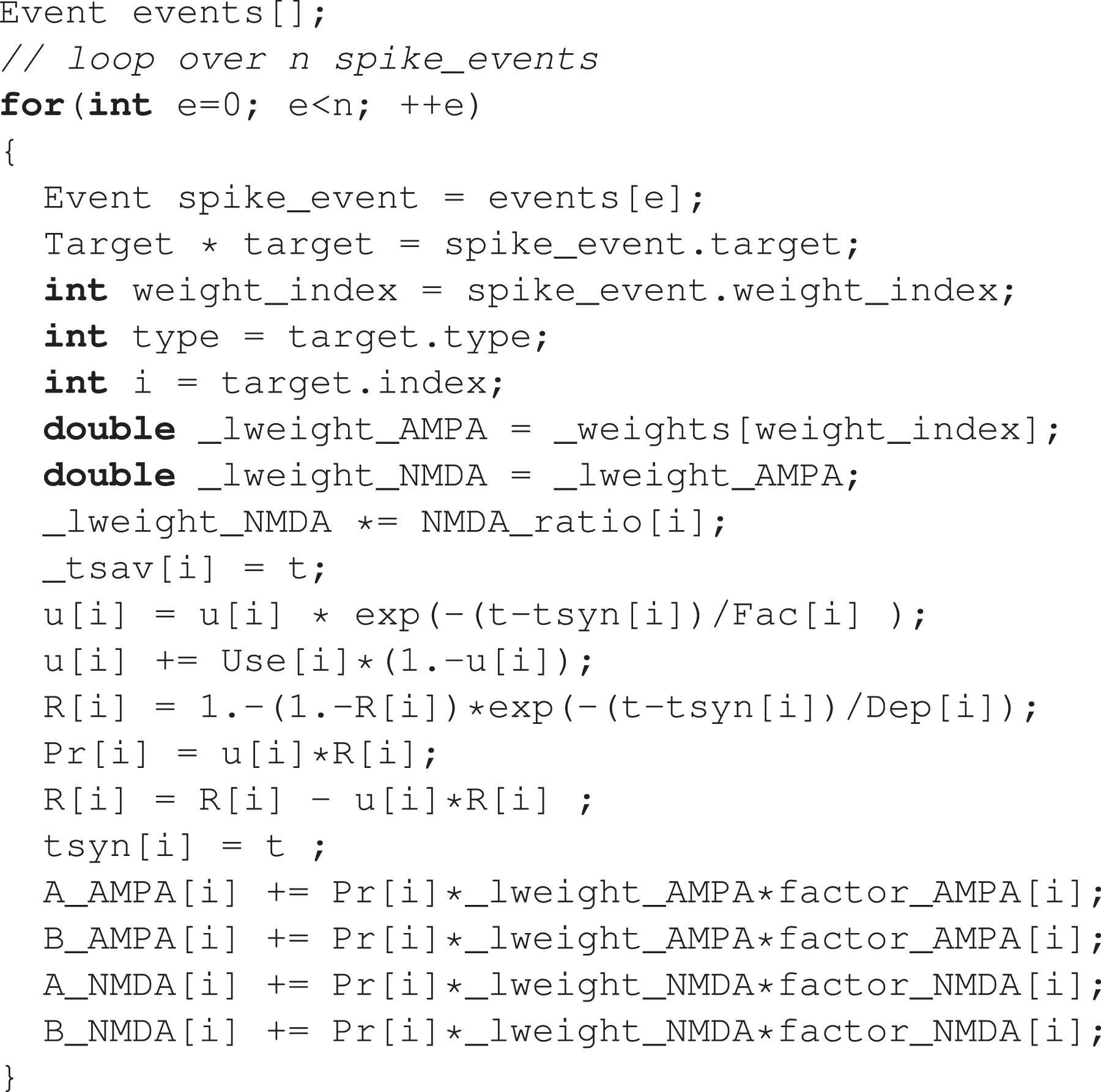
This kernel is characterized by erratic memory accesses indexed by
ECM model per scalar iteration (cy) for the spike delivery kernel.a
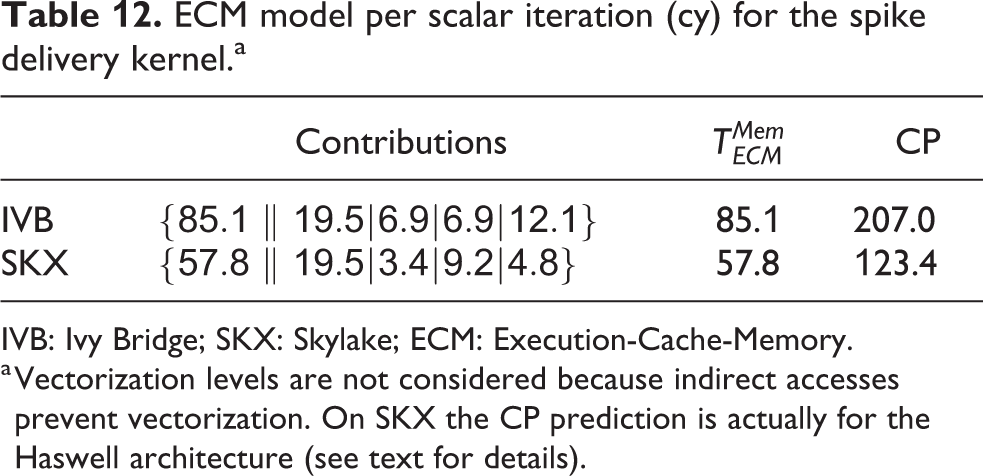
IVB: Ivy Bridge; SKX: Skylake; ECM: Execution-Cache-Memory.
a Vectorization levels are not considered because indirect accesses prevent vectorization. On SKX the CP prediction is actually for the Haswell architecture (see text for details).
Spike delivery runtime predictions and median measurements (
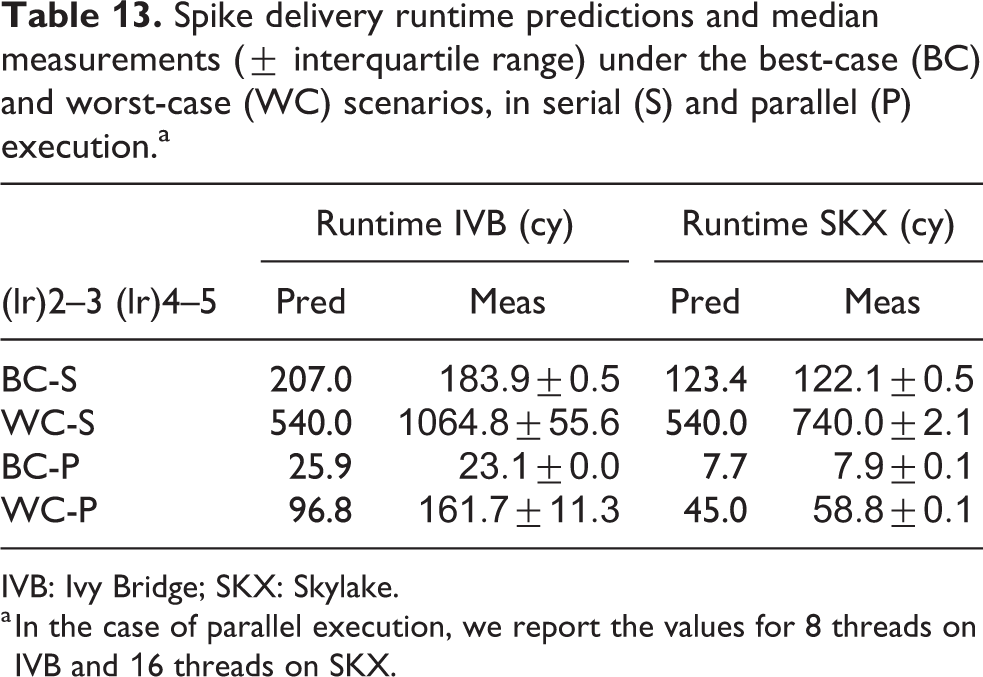
IVB: Ivy Bridge; SKX: Skylake.
a In the case of parallel execution, we report the values for 8 threads on IVB and 16 threads on SKX.
In the worst-case scenario, we assume that a full cache line of data needs to be brought in from memory for every data access. Assuming that the variables
Possible causes for degradation of accuracy in ECM model.

ECM: Execution-Cache-Memory; CP: critical path.
For validation, we designed a benchmark data set with 16 neurons and
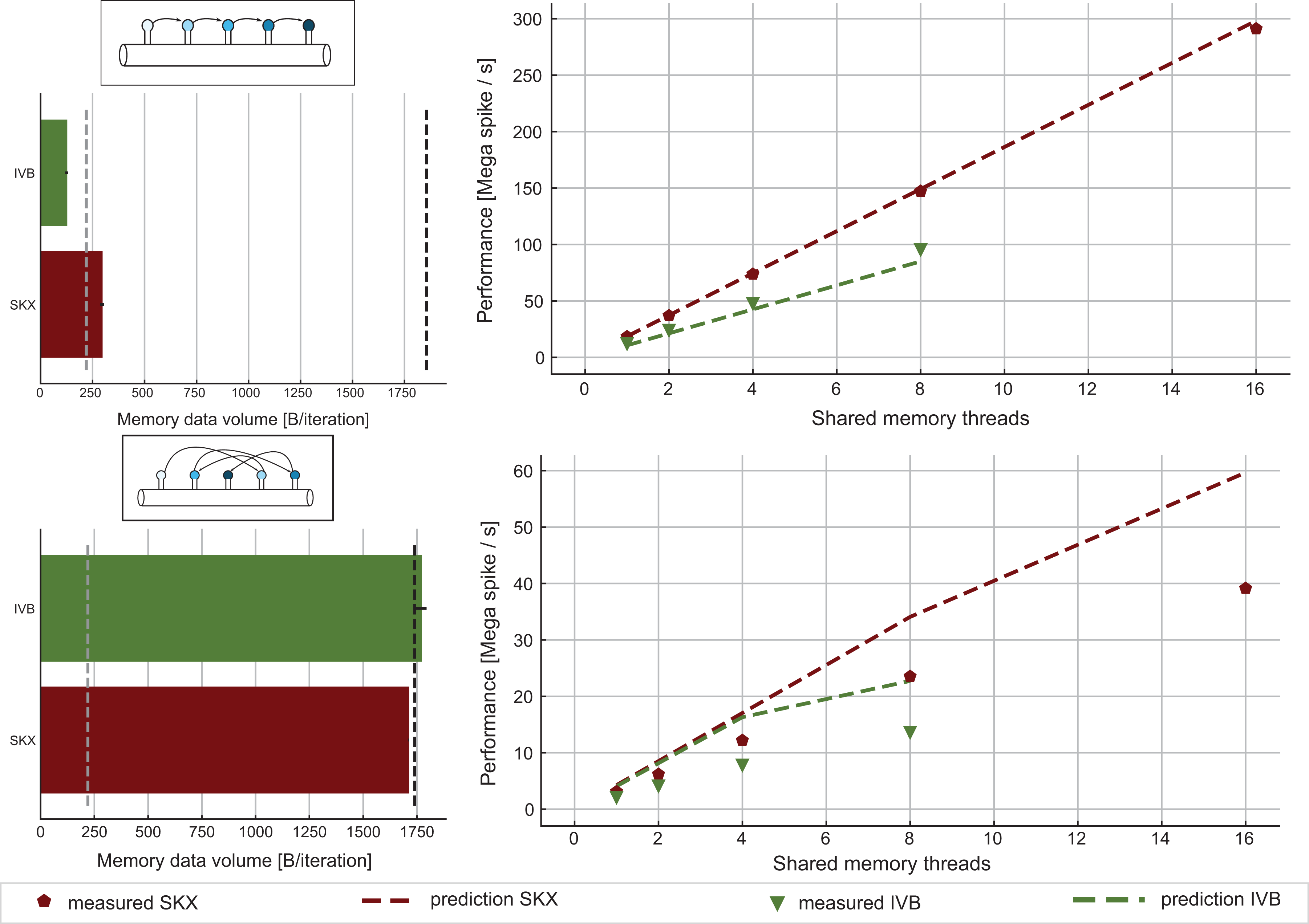
ECM model and measurements for the spike delivery kernel. Top: Best-case scenario where synapses are activated in contiguous memory order. In this case, there is no excess data traffic as shown by the bar plot on the left. The performance predictions on the right (dashed lines) are made by assuming that the kernel’s runtime is equal to its CP as predicted by IACA. Measurements (solid markers) substantiate this hypothesis. Bottom: Worst-case scenario where synapses are activated in random order. This scenario corresponds to the typical use-case. We assume that for every array access a full cache line of data traffic is generated, but only one element of the array is relevant. ECM: Execution-Cache-Memory; IACA: Intel Architecture Code Analyzer; CP: critical path.
4. Discussion
Using the ECM performance model, we have analyzed the performance profile of the simulation algorithm of morphologically detailed neurons as implemented in the CoreNEURON package. Within its design space, the ECM model yielded accurate predictions for the runtime of 13 kernels on real-world data sets. It must be stressed that some of these kernels are rather intricate, with hundreds of machine instructions and many parallel data streams. This confirms that analytic modeling is good for more than simple, educational benchmark cases. We have also, for the first time, set up the ECM model for the Intel Skylake-X architecture, whose cache hierarchy differs considerably from earlier Intel server CPUs. Our analysis shows that the non-overlapping assumption applies there as well, including all data paths between main memory, the L2 cache and the victim L3. Note that a reproducibility appendix is available at Cremonesi et al. (2019).
As expected, the modeling error was larger in situations where the bottleneck was neither streaming data access nor in-core instruction throughput. By making a few simplifying assumptions we were still able to predict with good accuracy the performance of a kernel with a complex memory access pattern and dependencies between loop iterations such as the tridiagonal Hines solver Hines (1984).
On the other hand, if the bottleneck is the memory latency, which is the case with the spike delivery kernel, the ECM model could only provide upper and lower bounds. In this case where the deviation from the measurement was especially large, we could at least pinpoint possible causes for the failure. It is left to future work on the foundations of the ECM model to extend its validity in those settings.
In conclusion, the ECM model was always able to correctly identify the computational characteristics and thus the bottlenecks of the 14 kernels under investigation, thus providing valuable insight to the performance-aware developers and modelers. In the following we use these crucial insights to give clear guidelines for both the optimization of simulation code and the codesign of an “ideal” processor architecture for neuron simulations. Given that memory locality plays a crucial role in our analysis, we structure our discussion based on the size of the biological network being simulated, that is, the total number of neurons in the data set. We mostly concentrate on the Skylake-X architecture since it is the most recent one, and no ECM model was published for it to date. Where relevant, we compare to the Ivy Bridge architecture to show the benefit of hardware improvements over three generations of Intel server processors.
4.1. Small networks (in cache)
4.1.1. Serial performance properties of small networks
One of the main insights offered by the ECM model is the possibility to identify and quantify the performance bottleneck of each kernel. In the simulation of morphologically detailed neurons, we found that ion channel current kernels are data bound while all state kernels are core bound for all cache levels, all SIMD levels and both architectures considered. The case of excitatory synapse current kernel was special in that on both SKX and IVB, the kernel was core bound as long as the data set fits in the caches, but switched to data-bound when the data comes from memory. This effect was most prominent on SKX-AVX512. In the extreme strong scaling scenario where data fits in the cache, this points to two optimizations: optimize expensive operations such as
4.1.2. SIMD parallelism and small networks
The possibility to execute high-throughput SIMD vector instructions can potentially provide great returns in terms of speedup at a low hardware and programming cost. In this analysis, we observed that wider SIMD units were indeed capable of providing benefits in terms of reduced runtime, but we also failed to observe the ideal speedup factor. Moreover, Skylake-X showed diminishing returns as the SIMD units grew wider. Applying the ECM model to the scenario where data comes from cache, we discovered that all state kernels show significant speedups from vectorization and would benefit even more from even wider SIMD units. The synapse current kernels benefit from SIMD instructions at least for data in the L1 or L2 cache. Ion channel current kernels show only small speedups from vectorization because their performance is solely determined by the speed of the data transfer, even when the working set fits into a cache.
The importance of high-throughput
4.2. Large networks (out of cache)
4.2.1. Memory bandwidth saturation of large networks
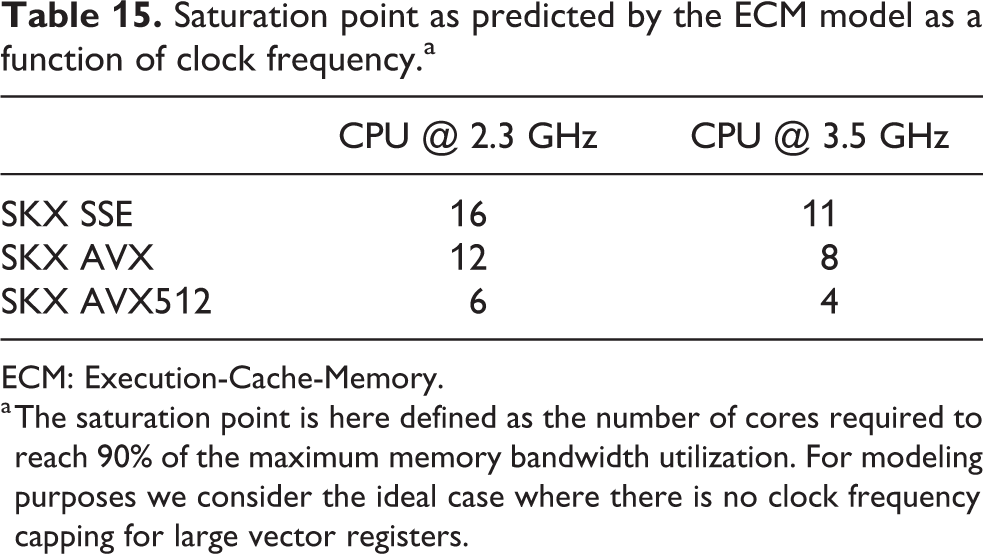
At constant memory bandwidth, a sufficient number of cores and/or high enough clock speed will render almost every code memory bound. One of the key insights delivered by the ECM model is how many cores are required to achieve saturation of the memory bandwidth during shared-memory execution, and what factors this number depends on. We applied saturation analysis to the full simulation loop by predicting the memory bandwidth of each kernel for different numbers of cores and compared it to the ratio of measured memory bandwidth to theoretical maximum bandwidth, weighted by the runtime of each kernel. Figure 6 shows the results, highlighting the remarkable power of the AVX512 technology on SKX, which is able to almost fully saturate the memory bandwidth using only seven cores. Since in-core features come essentially for free but more cores are more expensive, this means that in the max-filling scenario where the number of neurons being simulated is large and the data fits in main memory, the most cost-effective hardware platform for this code among the architectures considered is one with AVX512 support, high clock speed and a moderate core count. To further quantify the tradeoff between clock speed and saturation on SKX-AVX512, we computed the saturation point, which we define as the number of threads required to utilize at least 90% of the theoretical memory bandwidth, at different clock frequencies for the SKX architecture (assuming no clock frequency reduction). The results in Table 15 highlight once again that, as long as the working set is in main memory, vectorization pushes the bottleneck towards the memory bandwidth in the shared-memory execution. We have to allow some room for error in the measurements of the memory bandwidth and the over-optimistic ECM model near the saturation point as shown in Stengel et al. (2015), but the model indicates clearly that cores can be traded for clock speed, which provides a convenient price-performance optimization space.
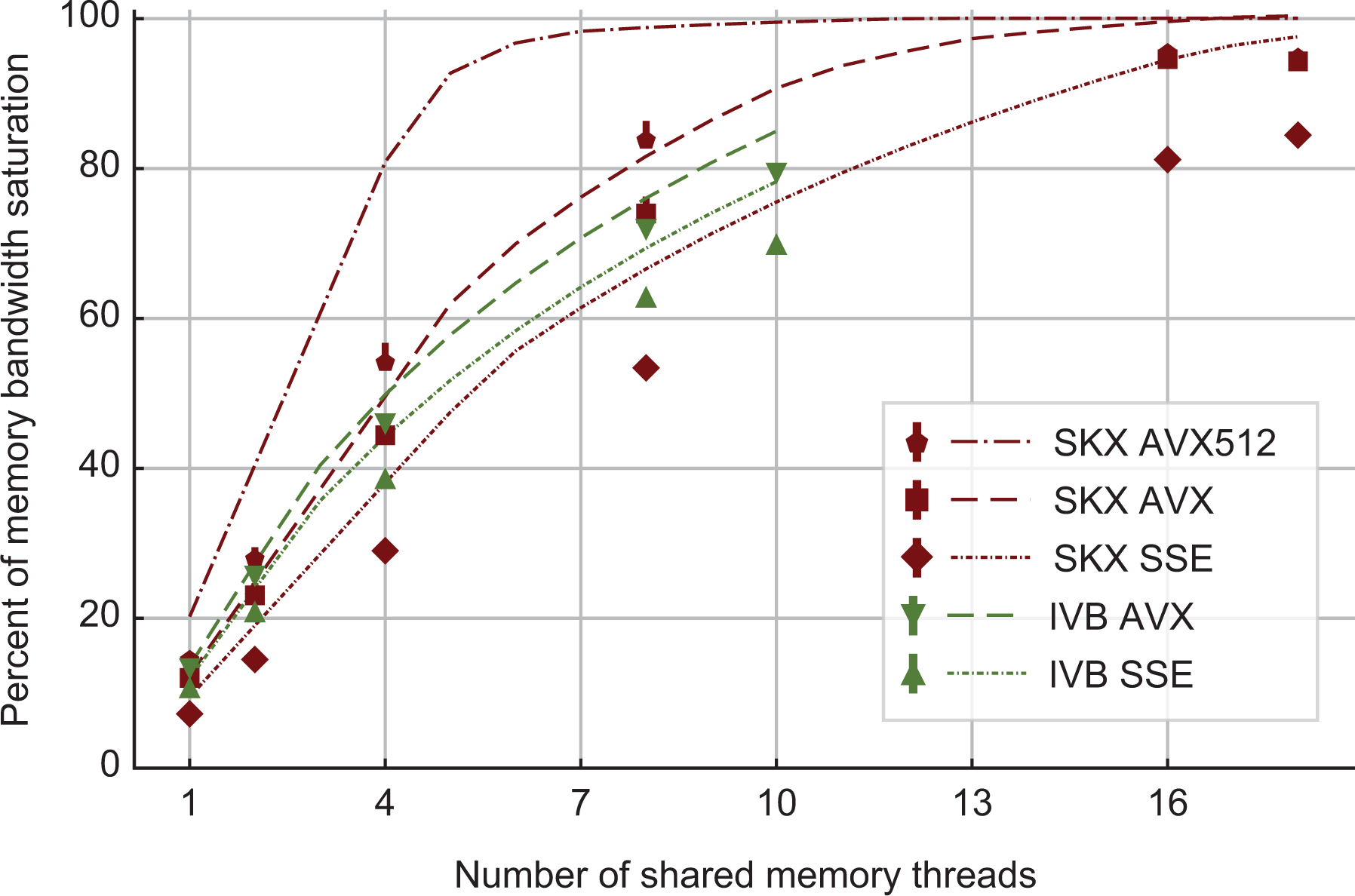
Predicted and measured memory bandwidth utilization, as a fraction of the maximum memory bandwidth. Dashed lines are obtained by predicting the average memory bandwidth of the full application while solid markers represent bandwidth measurements by LIKWID. Due to automatic clock frequency scaling, the maximum memory bandwidth was rescaled by the ratio of average clock frequency to nominal clock frequency. On SKX, AVX512 code can saturate the memory bandwidth of the socket at less than half the total number of cores even at the base clock frequency. SKX: Skylake.
Saturation point as predicted by the ECM model as a function of clock frequency.a
ECM: Execution-Cache-Memory.
a The saturation point is here defined as the number of cores required to reach 90% of the maximum memory bandwidth utilization. For modeling purposes we consider the ideal case where there is no clock frequency capping for large vector registers.
4.2.2. Wide SIMD and large networks
For in-memory data sets, wide SIMD execution helps to push the saturation point to a smaller number of cores, as shown in Table 15 and Figure 6, but it will certainly not increase the saturated performance. Hypothetical hardware with even wider SIMD units would thus have to be supported by a larger memory bandwidth to be fully effective. Moreover, as clearly shown by the ECM model analysis, wider SIMD execution would ultimately make even the state kernels data bound. In the mid-term future it would hence be advisable to put more emphasis on fast clock speeds and better memory bandwidth than on pushing towards wider SIMD units, at least for the workloads discussed in this work.
When choosing the most fitting cluster architecture one is thus left with the decision between a larger number of high-frequency chips with moderate memory bandwidth and a smaller amount of lower-frequency chips with large memory bandwidth and more cores. Roughly speaking, larger bandwidth is more expensive than faster clock speed, but the decision has to be made according to the market and pricing situation at hand, which unfortunately tends to be rather volatile.
4.2.3. Memory hierarchy for large networks
There is practically no temporal locality in the data access patterns of almost all kernels. This means that cache size is insignificant for determining the performance of large networks of detailed neuron simulations. Unfortunately, cache size is not a hardware parameter that one is free to choose when procuring clusters of off-the-shelf CPUs. Moreover, using the decomposition of the runtime by the ECM model, we observe that contributions from different levels of the memory hierarchy are rather evenly distributed. Hence, the runtime of data-bound kernels could best be improved by reducing the data volume. A common programming technique to solve this problem is loop fusion, by which two or more back-to-back kernels that read or write some common data structures are cast into a single loop in order to leverage temporal locality and thus increase the arithmetic intensity. The structure of the NEURON code does not easily allow this.
5. Conclusions
In this work, we have demonstrated the applicability of the ECM analytic performance model to analyze and predict the bottlenecks and runtime of simulations of biological neural networks. The need for such modeling is demonstrated by the ongoing development efforts to optimize simulation code for current state of the art HPC platforms, coupled with demands for simulators able to handle faster and larger data sets on present and future architectures. Using the performance model we identified high-frequency cores capable of high-throughput
No attempts have been made so far towards porting NEURON kernels to traditional vector processors (which have again become available recently), and porting to GPGPUs is still in an exploratory phase, but at least for large networks, where abundant parallelism is available, the characteristics we have identified let us expect speedups according to the memory bandwidth difference to standard multicore CPUs: a device with 1 TB/s of memory bandwidth, such as the SX-Aurora “Tsubasa” by NEC (2018), should outperform one Skylake-X socket by a factor of 9–10.
In the reconstruction and simulation of brain tissue, performance engineering and modeling is now a pressing issue limiting the scale and speed at which computational neuroscientists can run in silico experiments. We believe that our work represents an important contribution in understanding the fundamental performance properties of brain simulations and preparing the community for the next generation of hardware architectures.
6. Future work
Two shortcomings hinder the comprehensive applicability of the ECM model for all the kernels in CoreNEURON: the inability to correctly describe latency-bound data accesses, and long CPs in the loop body. Both shortcomings may be addressed by refining the model, that is, endowing it with more information about the processor architecture. This data, however, is not readily available (and it might never be). In case of CP analysis, the Open Source Architecture Code Analyzer (OSACA) by Laukemann et al. (2018) is planned to become a versatile substitute for IACA, which does not provide CP prediction for modern Intel CPUs. OSACA has recently been extended to support CP and loop-carried dependency detection (see Laukemann et al., 2019). Data latency support would require a fundamental modification of the model, and work is ongoing in this direction.
Supplemental Material
Supplemental Material, ijhpca-CHWS - Analytic performance modeling and analysis of detailed neuron simulations
Supplemental Material, ijhpca-CHWS for Analytic performance modeling and analysis of detailed neuron simulations by Francesco Cremonesi, Georg Hager, Gerhard Wellein and Felix Schürmann in The International Journal of High Performance Computing Applications
Supplemental Material
Supplemental Material, Rebuttal_BBP-IJHPCA_2019 - Analytic performance modeling and analysis of detailed neuron simulations
Supplemental Material, Rebuttal_BBP-IJHPCA_2019 for Analytic performance modeling and analysis of detailed neuron simulations by Francesco Cremonesi, Georg Hager, Gerhard Wellein and Felix Schürmann in The International Journal of High Performance Computing Applications
Footnotes
Acknowledgments
The authors would like to thank Johannes Hofmann for fruitful discussions about low-level benchmarking and Thomas Gruber for his contributions to the LIKWID framework. The authors are also indebted to the Blue Brain HPC team for helpful support and discussion regarding CoreNEURON.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by funding to the Blue Brain Project, a research center of the École polytechnique fédérale de Lausanne, from the Swiss government’s ETH Board of the Swiss Federal Institutes of Technology.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.