
Abstract
Bisphenol A (BPA) is an estrogenic chemical used in the production of polycarbonate plastics and epoxy resins. Our earlier studies have demonstrated that BPA is a potent reproductive and genotoxic agent and affects the normal physiological functions. The objective of this study was to evaluate whether exposure to BPA induces oxidative stress. The male Holtzman rats were orally gavaged with BPA (0.01 mg and 5.0 mg/kg/bw) over the period of 6 days. Animals were euthanized by cervical dislocation at the end of the treatments; bone marrow cells and blood lymphocytes were aspirated; testis and epididymis were collected, immediately frozen in liquid nitrogen, and stored at −80°C. These samples were utilized for the determination of lipid peroxidation and various antioxidant enzymes such as superoxide dismutase, catalase, and nonenzymatic reduced glutathione. The results demonstrated that BPA caused an increase in lipid peroxidation and a decrease in activity of various enzymatic and nonenzymatic antioxidants in bone marrow cells, blood lymphocytes, and testicular and epididymal tissues. The findings of the current study suggest that BPA exposure induced oxidative stress, which could be one of the possible mechanisms causing reproductive and genetic toxicity.
Introduction
There is an increasing concern about the environmental contaminants that disrupt the male reproductive function of humans and play a major role in the decline of quality and quantity of semen. 1 Bisphenol A (BPA) is one of the environmental toxicants demonstrated in both in vitro and in vivo assays to act as an endocrine-disrupting chemical. 2 It is widely used in the manufacture of polycarbonate plastic (eg, water bottles, baby bottles), epoxy resins (eg, inside coating of metallic food cans), dental sealants, and as a nonpolymer additive to other plastics. 3 Human population are widely exposed to BPA, as it leaches from the inner lining of tin cans and microwave containers during heating of food materials, 4 from dental sealant into saliva, 5 and into beverages from polycarbonate bottles due to the repeated usage or contact with any acidic/alkaline contents. 6 It has been also observed that BPA enters the environment via open disposal or recycling of products which contains BPA. 7 The BPA levels have been estimated in various environmental samples such as surface water (0.0005-0.41 mg/L), sewage effluent (0.018-0.702 mg/L), sediments (0.01-0.19 mg/kg), sewage sludge (0.004-1.363 mg/kg), and landfill leachate. 8 -10
A significant concentration of BPA has been detected in urine and serum samples of human population, 11 in maternal and fetal plasma, 12 in placental tissue, 13 and in the milk of lactating mothers. 14 The increased BPA levels in maternal serum and amniotic fluid have been associated with abnormal karyotype in the fetuses at early second trimester miscarriage. 15 An association between blood BPA levels and history of 3 or more consecutive first trimester miscarriages has been documented, which was approximately 3 times higher than controls with no history of live birth, infertility, or miscarriages. 16
Rodent studies have shown that prenatal exposure of rats to BPA is associated with an increased risk of breast cancer in adult female rats 17 and hyperplasia of prostate in male rats, resulting in greater risk of prostate cancer. 18 Bisphenol A has the potential to induce aneuploidy and DNA adduct formation in Syrian hamster embryo cells. 19 Moreover, BPA has been shown to form DNA adduct in both liver and mammary cells of female CD-1 mice. 20 Bisphenol A treatment has been shown to disrupt the cytoplasmic microtubular complex as well as mitotic and meiotic spindle formations. 21,22 It also induced aneuploidy and chromosome congression failure in oocytes of mice exposed to low concentration of BPA. 23 Bisphenol A has been shown to disrupt the expression of thyroid hormone–related genes in aquatic species. Bisphenol A downregulated the expression of T3-response genes, including stromelysin 3, leucine zipper transcription factor, metalloproteinase 2, and the tissue inhibitor of metalloproteinase 2 in the African clawed frog. 24 It has been also demonstrated that BPA retarded hatching in zebra fish 25 and slowed hatching, yolk absorption, and first feeding in juvenile rainbow trout. 26
Previous studies from our laboratories on reproductive toxicity of BPA have documented that perinatal exposure of pregnant females caused significant impairment in the fertility of not only F1 male offspring but also subsequent F2 and F3 generations. 27 It also caused an increase in postimplantation loss in all the 3 generations; however, this increase was significant in F3 generation, indicating that BPA exerts its effects via male germ line. 27 In another study, normal cycling female rats when mated with adult male rats exposed neonatally to BPA showed a significant increase in postimplantation loss. 28 These males also exhibited a significant change in sperm count and motility along with hormonal imbalance. Our group has also demonstrated the effects of neonatal exposure of male rat to BPA leading to aberrant DNA methylation in testis of adult male rats. 29 Furthermore, a significant hypomethylation observed at the H19 ICR in the spermatozoa and resorbed embryo caused perturbation in the expression of Igf2 and H19, ultimately leading to postimplantation loss. 30 These reports indicate that apart from endocrine effect, BPA acts via epigenetic mode, which could be one of the possible mechanisms of BPA-induced adverse epigenetic effects on male fertility. The BPA exposure has been also reported to alter the expression of peroxisome proliferator-activated receptors (PPARs) in mammals. 31 The PPARs is considered to be the primary mechanism of action in plasticizer-induced transcriptional changes, 32 reproductive organ toxicity, 31 and oxidative stress. 33
In a recent study, we have demonstrated that exposure to BPA for 6 consecutive days in adult male and female rats at no obseved adverse effect level (NOAEL) dose (5.0 mg/kg/bw/d) 34 and even at 10 µg/kg/bw/d led to a significant increase in clastogenic activity in bone marrow cells and DNA fragmentation in blood lymphocytes. 35 Further, our group also demonstrated that BPA exposure led to a significant increase in postimplantation loss, dominant lethal mutation, and sperm DNA damage and a decrease in testicular daily sperm production, epididymal sperm count, and motility. 36 Bisphenol A has been reported to induce reactive oxygen species (ROS), thereby altering the level antioxidants in liver 37 and epididymal sperm of rats. 38
Although the reproductive and genetic toxicity of BPA has been demonstrated in experimental animals, the mechanism of action needs to be elucidated. Several studies have reported that oxidative stress leads to DNA damage. 39 -41 The present study was undertaken to evaluate whether exposure to BPA induces oxidative stress in bone marrow cells, lymphocytes, and reproductive organs.
Materials and Methods
Chemicals
Bisphenol A (CAS No. 80-05-7; ∼ 99% purity) was purchased from Sigma Chemical Co (St Louis, Missouri). Phosphoric acid, metaphosphoric acid, butylated hydroxytoluene (BHT), n-butanol, EDTA, and malondialdehyde (MDA) were obtained from Merck Schuchardt (Germany). Thiobarbituric acid and pyrogallol were obtained from HiMedia Laboratories (Mumbai, India). 1, 1, 3, 3-Tetraethoxypropane, 5,5-dithiobis 2-nitrobenzoic acid (DTNB), and Histopaque 1077 solution were obtained from Sigma Aldrich (Germany) .
Dose Selection
The BPA exposure is reported to exert its biological effects at very low doses similar to amounts typically found in environment, which give rise to much concern on the risk of human exposure to BPA. 42 Earlier studies by our group demonstrated that 2.4 and 10 μg/kg dose of BPA were capable of inducing reproductive toxic effects in rats. 27 Recently, we have showed that a dose of 5.0 mg/kg/bw and even at 10 µg/kg/bw led to a significant increase in clastogenic activity in bone marrow cells and DNA fragmentation in blood lymphocyte of treated Holtzman rats. 35 Based on the above information, following doses, namely, 0.01 mg and 5.0 mg/kg/bw, were selected in the present study.
Dose Preparation
Bisphenol A was dissolved in distilled ethyl alcohol (99.9% purity) and diluted with sesame oil to obtain a final concentration (0.01 mg and 5.0 mg/kg/bw) of BPA. The dose formulations were stored in an amber-colored bottle and kept at 37°C overnight. They were subsequently kept at room temperature throughout the study. Sesame oil was used as a vehicle control.
Animal Handling and Care
Holtzman strain male and female rats (8 weeks of age) that were randomly bred in our animal house were used in the present study. The animals were kept in cages with autoclaved paddy husk for bedding and maintained under controlled temperature (22°C ± 2°C), humidity (50%-70%), and 14-hour:10-hour dark/light cycle. The animals were fed a diet of soy-free, in-house-prepared rat pellets (consisting of crude protein, fiber, and nitrogen-free extract) and water (purified by ultraviolet and reverse osmosis) ad libitum throughout the study. The quality of food and water provided was routinely monitored by qualitative and quantitative analysis. The ethical clearance for the use of animals in the study was obtained from the institutional animal ethics committee prior to the initiation of the study. The experiments were performed in accordance with the guidelines of the Committee for the Purpose of Control and Supervision of Experimental Animals, India.
Animal Treatment
The rats were randomly divided into 3 groups, each consisting of 5 male rats. Group 1 was administered sesame oil by oral gavage (1.0 mL/kg/bw), which served as vehicle control. Groups 2 and 3 were administered with different concentrations of BPA (0.01 mg and 5.0 mg/kg/bw) by oral gavage once a day for the period of 6 days. Rats were euthanized by cervical dislocation 24 hours after the last dose administered.
Lymphocytes and Bone Marrow Cell Preparation
After the 24 hours of last dose administered, the rats were anesthetized by intraperitoneal injection of ketamine/xylazine, and blood samples were collected in heparinized tube via the retro orbital plexus. 43 Immediately after the blood collection, these rats were sacrificed by cervical dislocation. The blood samples were diluted with phosphate-buffered saline (PBS; pH 7.0) in equal ratio and then layered on the density gradient (Histopaque 1077) in 1:2 ratios. The white milky layer of mononuclear cells, that is, lymphocytes, was carefully removed after centrifugation at 500g for 20 minutes. The layer was then washed twice with the same buffer and centrifuged at 3,000g for 10 minutes to get the required pellet of lymphocytes. Simultaneously, both femur bones of same rats were taken out by dissection and cleaned to remove muscles and other tissues, their ends were cut, bone marrow cells were aspirated using syringe and needle (21 G) with 3.0 mL of fetal bovine serum (FBS), and centrifuged at 1,200g for 4 minutes. Cell pellets were washed twice with 2.0 mL of PBS to remove FBS. The lymphocytes and bone marrow pellets were weighed using automatic electronic balance and homogenized in ice-cold 0.9% NaCl to produce 1:9 homogenates. Homogenates were centrifuged at 10,000 rpm for 10 minutes at 4°C. The supernatant was collected and frozen at −20°C for the assessment of superoxide dismutase (SOD), catalase (CAT), glutathione (GSH), and MDA.
Testicular and Epididymal Preparation
From the same treated animals, testis and epididymis were rapidly excised on a petri dish which were placed on ice and were rinsed and washed in ice-cold normal saline solution. They were weighed using automatic electronic balance and homogenized using Teflon homogenizer in ice-cold 0.9% NaCl to produce 1:9 homogenates. Homogenates were centrifuged at 10,000 rpm for 30 minutes at 4°C. The supernatant was collected and frozen at −20°C for the assessment of SOD, CAT, GSH, and MDA.
Lipid Peroxidation
Lipid peroxidation (LPO) was determined in the various samples following the procedure. 44 Malondialdehyde forms as an intermediate product of the peroxidation of lipids and serves as an index of the intensity of oxidative stress. In brief, the 3.0 mL of 1% of phosphoric acid, 150 µL of BHT solution, and 500 µL samples were added in 15 mL of glass tubes. Afterward, 1.0 mL (0.6%) of TBA solution was added, and the mixture was vortexed for 30 seconds. The samples were then incubated in boiling water bath for 45 minutes. The tube was left for 10 minutes at room temperature for cooling. The 4.0 mL of n-butanol was added to the above glass tube, vortexed, centrifuged at 1,000 rpm for 15 minutes, and colored layer (orange-pink) was aspirated using the pasture pipettes. The absorbance of the colored layer was measured using spectrophotometer at 532, 530, and 520 nm. The difference was used for calculation of thiobarbituric acid–reactive substance (ie, MDA). The MDA standard was prepared from 1, 1, 3, 3-tetraethoxypropane in the range of 4.403 nmol/0.1 mL. It was proceeded in the same as the samples. The OD difference at 535 and 520 nm was obtained for MDA standards and used for calculating the tissue MDA levels. All values were expressed as nanomole MDA/g tissue.
Superoxide Dismutase
Superoxide dismutase level in the various samples of BPA-treated rats was determined by the pyrogallol autoxidation method. 45 The supernatant obtained from treated and vehicle control tissue was used for estimation of SOD (samples were kept on ice). In brief, 900 µL (50 mM Tris–HCl + 1 mM EDTA) of buffer and 50 µL of tissue supernatant were added sequentially in 15 mL of glass tube and vortexed. Further, the 50 µL (10 mM) of pyrogallol solution was added and the tubes were vortexed. Immediately, the absorbance of the solution was measured using spectrophotometer at 420 nm over the period of 3 minutes at the interval of 0, 60, 120, and 180 seconds.
Reduced GSH
Glutathione level in the various samples of BPA-treated rats was estimated by the method with a slight modification. 46 In brief, 250 µL of sample was added to the 1.76 mL of distilled water and 3.0 mL of 1.67% metaphosphoric acid to precipitate the protein, mixed, and centrifuged at 3,000 rpm for 10 minutes. The resulting supernatant obtained after centrifugation was then taken for GSH estimation. To the 2.0 mL of supernatant, 3.0 mL of 0.4 M Na2HPO4 buffer solution and 0.5 mL of DTNB were added, and then the intensity of yellow color was read at 412 nm. The GSH contents in the samples were expressed as micromole GSH/g tissue.
Catalase
Catalase activity in the various samples of BPA-treated rats was determined according to the method. 47 The supernatant obtained from treated and vehicle control was used for estimation of CAT (samples were kept on ice). In brief, 1.95 mL of 50 mM phosphate buffer (pH 7.0) and 50 µL of sample were mixed in the 15 mL of tube by vortexing, to which 1.0 mL of H2O2 (30 mM) was mixed and vortexed. Immediately, the absorbance of the colored layer was measured using spectrophotometer at 240 nm. The readings were recorded at intervals of 0, 15, and 30 seconds.
Statistical Analysis
The 1-way analysis of variance with Tukey multiple comparison tests was used to determine the level of significance between the control and treated groups for various samples. All the values were reported as mean (standard deviation). P < 0.05 was considered as statistically significant.
Results
Lipid Peroxidation
The BPA treatment caused a significant increase (P < 0.05) in the levels of LPO in different samples at doses 0.01 mg and 5.0 mg/kg/bw as compared to control. The content of MDA in a bone marrow cells and blood lymphocytes are depicted in Figure 1. The MDA levels increased by 69.0% and 119.7% in bone marrow cells and 36.4% and 90.9% in blood lymphocytes at 0.01 and 5.0 mg doses, respectively, as compared to control. Similar increases were observed in testicular and epididymal tissues, as depicted in Figure 2. The MDA levels were also increased in testicular (53.34% and 135.0%) and epididymal tissues (151.9% and 327.6%) at 0.01 and 5.0 mg doses as compared to control, respectively. Compared with the control group, a dose-dependent elevation of MDA contents was observed in each group. The increase in the extent of MDA level due to the increase in concentration of BPA suggests the excess generation of free radicals.
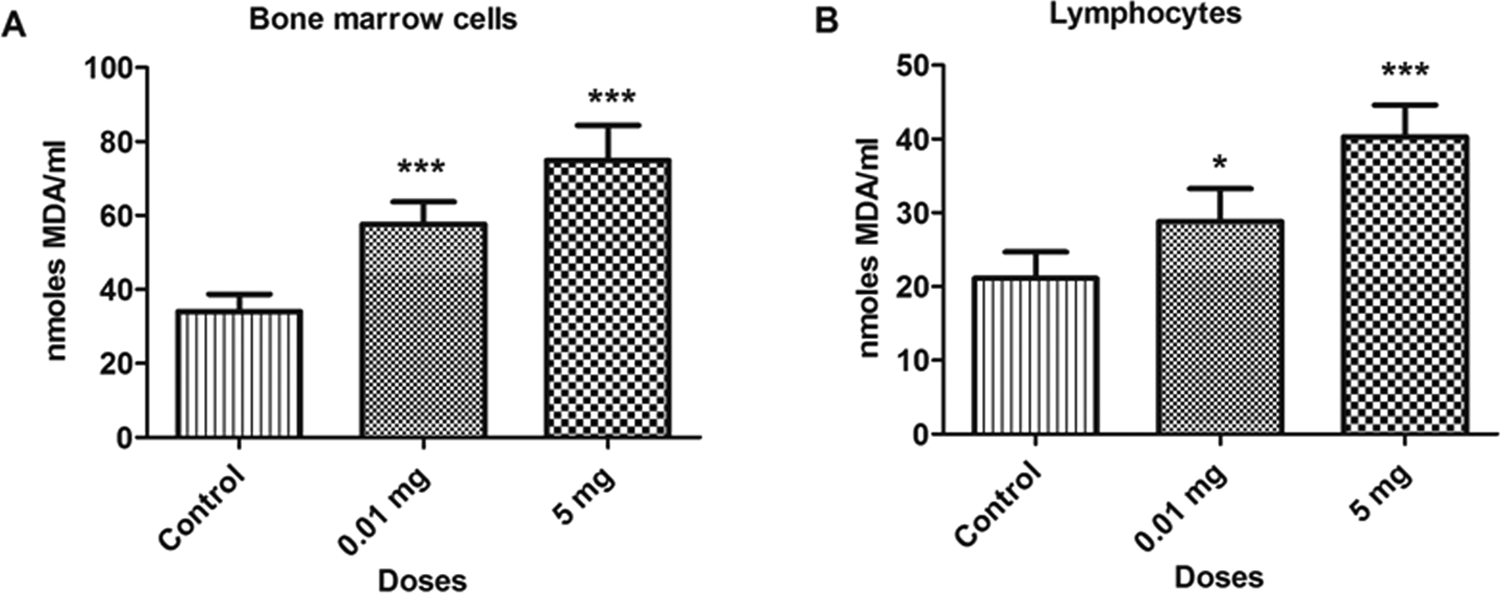
Bar diagrams showing an increase in lipid peroxidation between treated and control rats with the exposure to bisphenol A (BPA). A, Bone marrow cells. B, Lymphocytes. Data are shown as mean ± standard deviation (SD). *P < 0.05, ***P < 0.001. The significance level was set at P < 0.05.
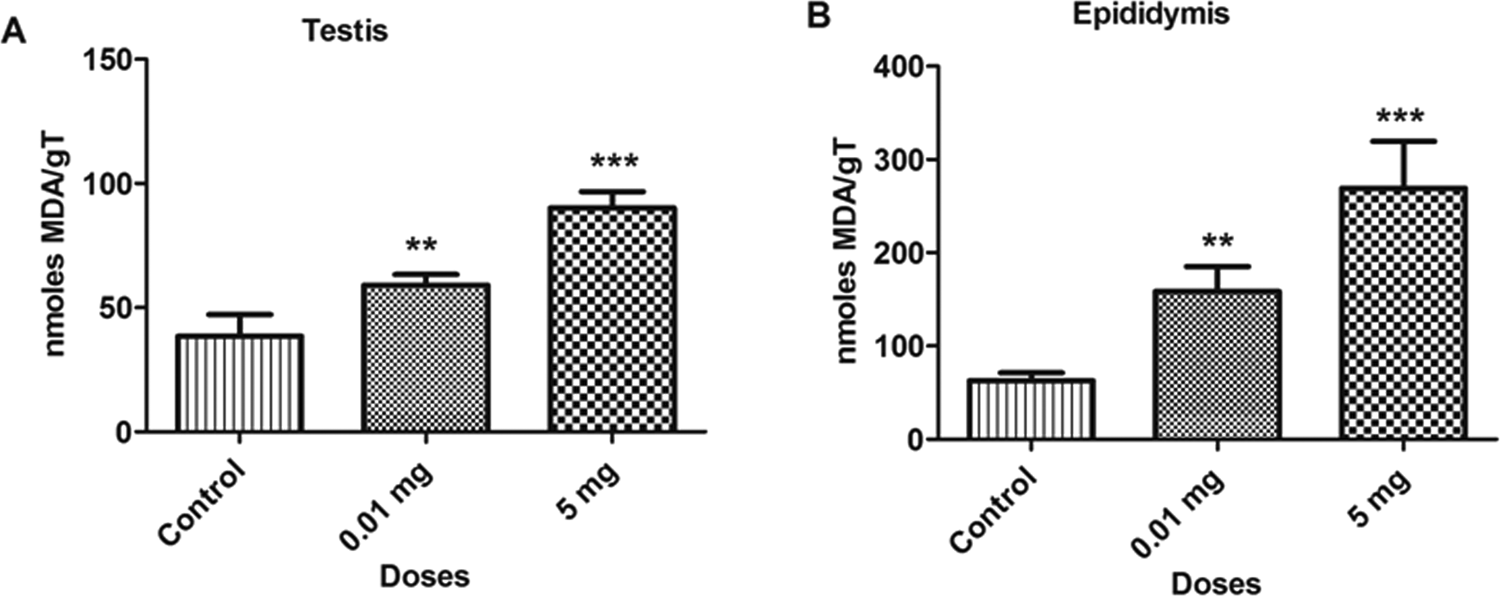
Bar diagrams showing an increase in lipid peroxidation between treated and control rats with the exposure to bisphenol A (BPA). A, testicular tissue. B, Epididymal tissue. Data are shown as mean ± standard deviation (SD). **P < 0.01, ***P < 0.001. The significance level was set at P < 0.05.
Superoxide Dismutase
A significant decrease was observed in SOD levels with BPA treatment (P < 0.05) in various samples at doses 0.01 and 5.0 mg/kg/bw as compared to control. The concentration of SOD in bone marrow cells and blood lymphocytes are depicted in Figure 3. The SOD levels decreased by 22.5% and 52.6% in bone marrow cells and 16.1% and 38.4% in blood lymphocytes at 0.01 and 5.0 mg doses, respectively, as compared to control. Similar decreases were also observed in testicular and epididymal tissues, which are depicted in Figure 4. The SOD levels was also decreased in testicular (38.1% and 76.8%) and epididymal tissues (36.2% and 55.6%) at 0.01 and 5.0 mg doses, respectively, as compared to control. The decrease in the level of SOD in various samples might be due to the inability of the cells to generate enough SOD or greater functioning in combating oxidative stress.
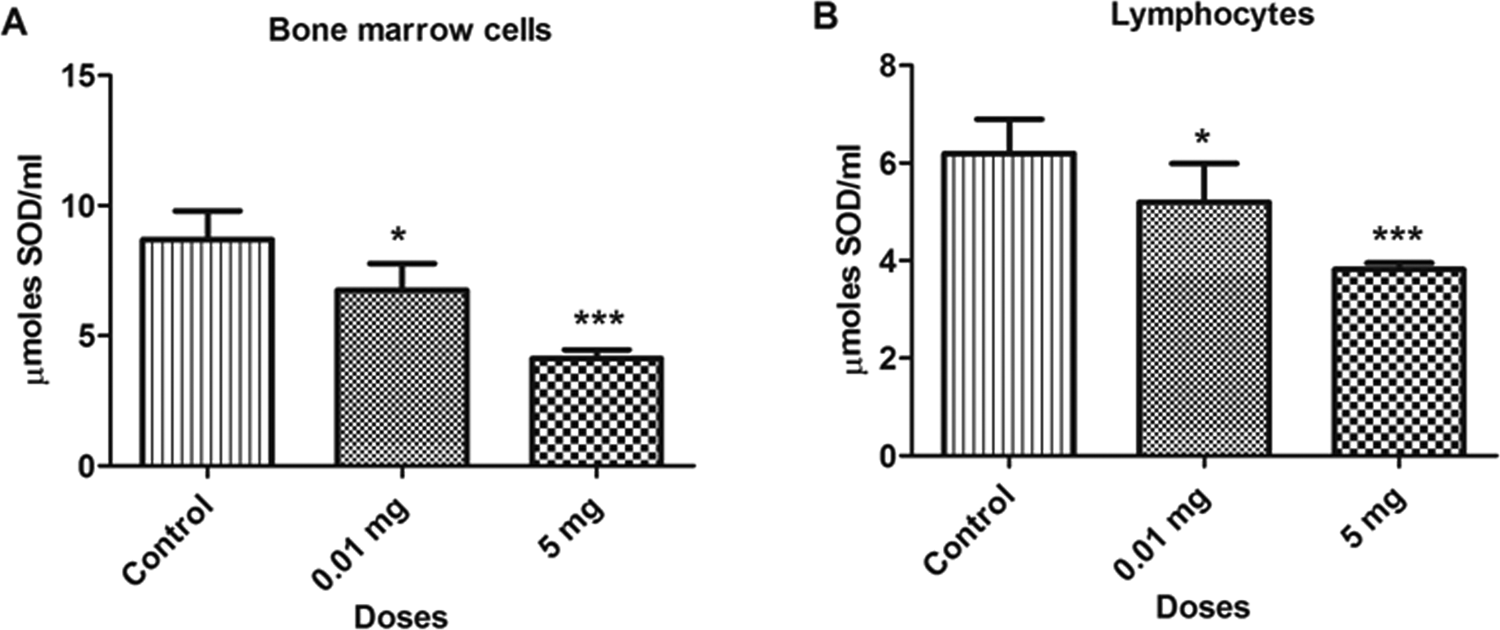
Bar diagrams showing a decrease in the superoxide dismutase (SOD) activity between treated and control rats with the exposure to bisphenol A (BPA). A, Bone marrow cells. B, Lymphocytes. Data are shown as mean ± standard deviation (SD). *P < 0.05, ***P < 0.001. The significance level was set at P < 0.05.
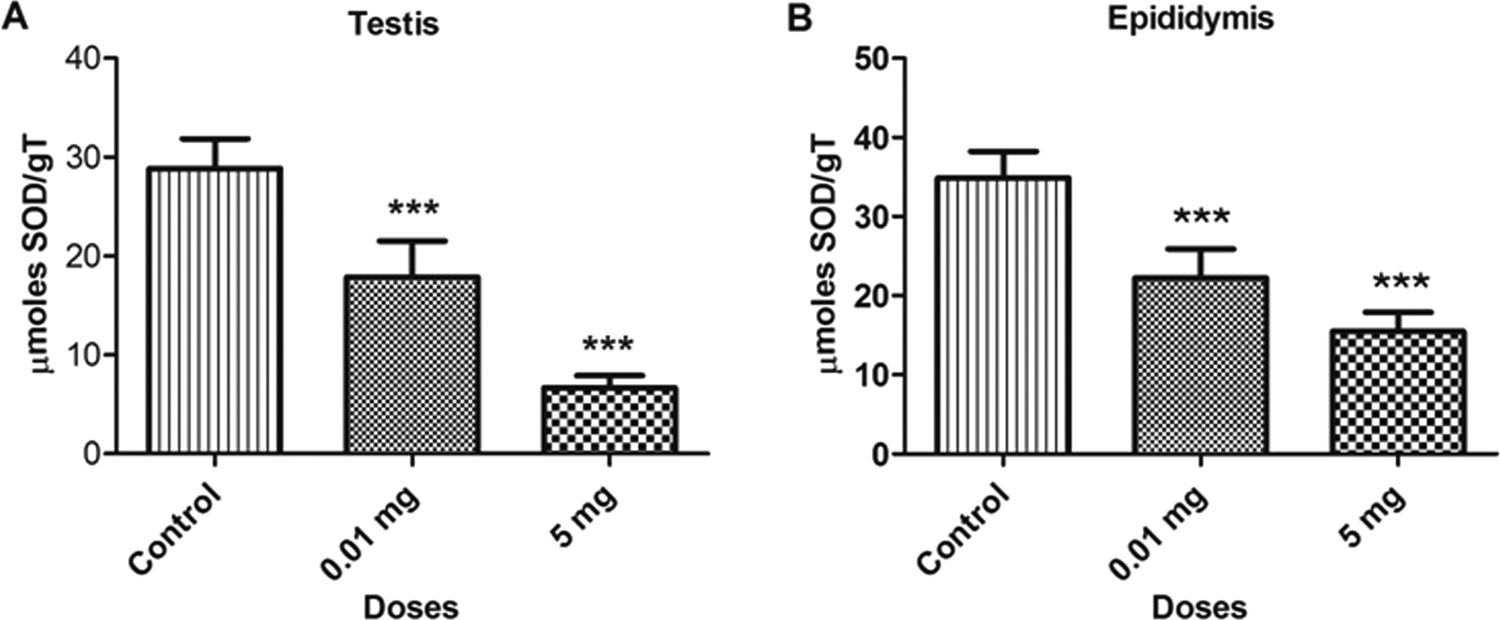
Bar diagrams showing a decrease in superoxide dismutase (SOD) activity between treated and control rats with the exposure to bisphenol A (BPA). A, Testicular tissue. B, Epididymal tissue. Data are shown as mean ± standard deviation (SD). ***P < 0.001. The significance level was set at P < 0.05.
Reduced GSH
The BPA treatment caused a significant decrease (P < 0.05) in the concentration of reduced GSH levels in different samples such as bone marrow cells, blood lymphocytes, as depicted in Figure 5. The GSH levels decreased by 22.9% and 47.5% in bone marrow cells and 50.1% and 69.6% in blood lymphocytes at 0.01 and 5.0 mg doses, respectively, as compared to control. Similar decreases were observed in testicular and epididymal tissues, which are depicted in Figure 6. The GSH levels was also decreased in testicular (31.9% and 50.6%) and epididymal tissues (43.6% and 52.7%) at 0.01 and 5.0 mg doses, respectively, as compared to control. The decrease in the GSH level with the increase in BPA concentrations shows consumption of GSH for the scavenging of ROS.
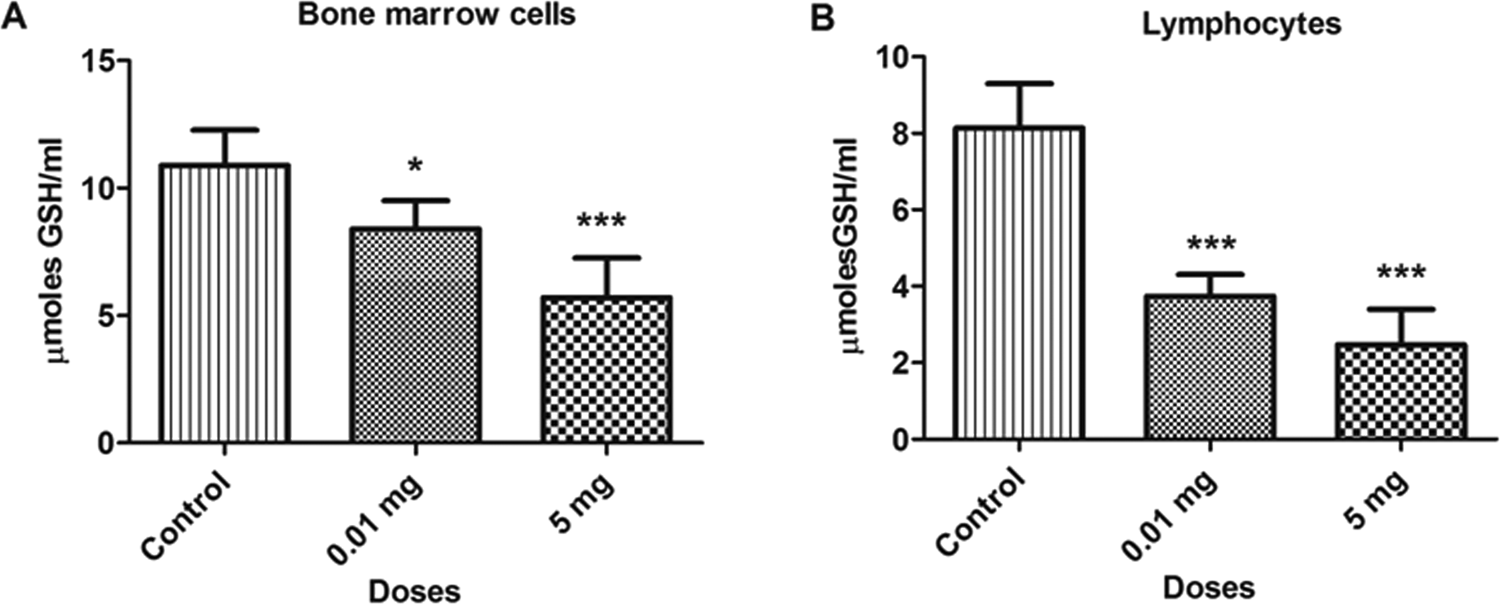
Bar diagrams showing a decrease in glutathione (GSH) activity between treated and control rats with the exposure to bisphenol A (BPA). A, Bone marrow cells. B, Lymphocytes. Data are shown as mean ± standard deviation (SD). *P < 0.05, ***P < 0.001. The significance level was set at P < 0.05.
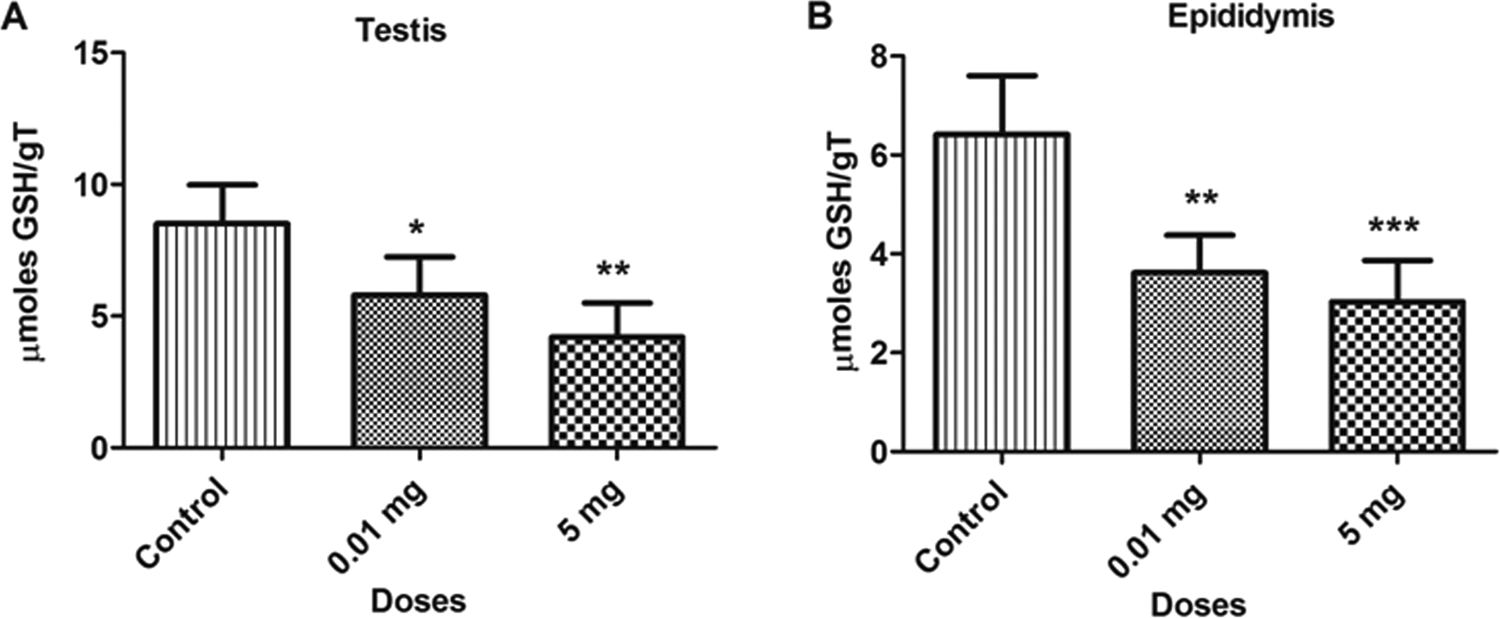
Bar diagrams showing a decrease in glutathione (GSH) activity between treated and control rats with the exposure to bisphenol A (BPA). A, Testicular tissue. B, Epididymal tissue. Data are shown as mean ± standard deviation (SD). *P < 0.05, **P < 0.01, ***P < 0.001. The- significance level was set at P < 0.05.
Catalase
The BPA treatment significantly decreased (P < 0.05) the levels of CAT activity in various samples at doses 0.01 and 5.0 mg/kg/bw as compared to the control group. The concentration of CAT in bone marrow cells and blood lymphocytes are depicted in Figure 7. The CAT levels decreased by 36.5% and 52.1% in bone marrow cells and 23.3% and 40.4% in blood lymphocytes at 0.01 and 5.0 mg doses, respectively, as compared to the control group. Similarly, the reduction in the levels of CAT was also observed in testicular and epididymal tissues, which are shown in Figure 8. The CAT levels was also decreased in testicular (43.1% and 56.7%) and epididymal tissues (44.9% and 59.7%) at 0.01 and 5.0 mg doses, respectively, as compared to control. The reduction in the CAT levels reflects the increase in oxidative stress due to which CAT was consumed to catalyze the decomposition of hydrogen peroxide to water and oxygen.
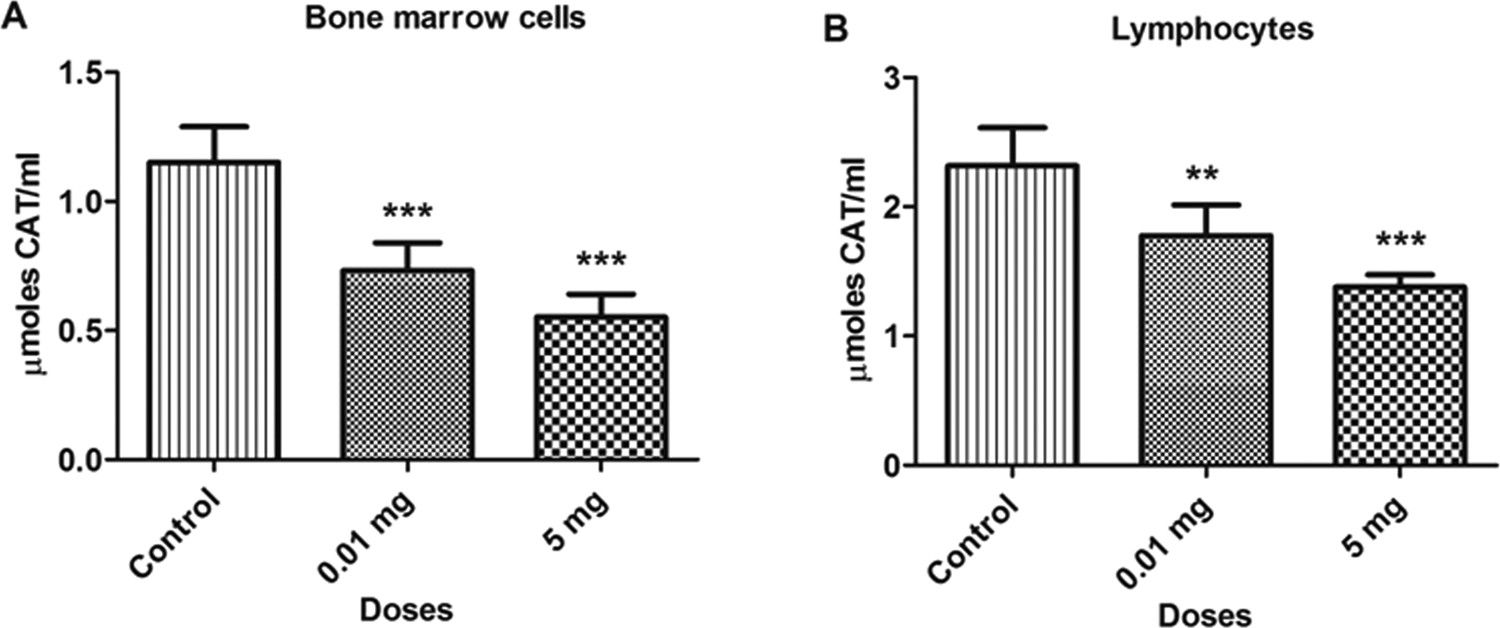
Bar diagrams showing a decrease in catalase activity between treated and control rats with the exposure to bisphenol A (BPA). A, Bone marrow cells. B, Lymphocytes. Data are shown as mean ± standard deviation (SD). **P < 0.01, ***P < 0.001. The significance was set at P < 0.05.
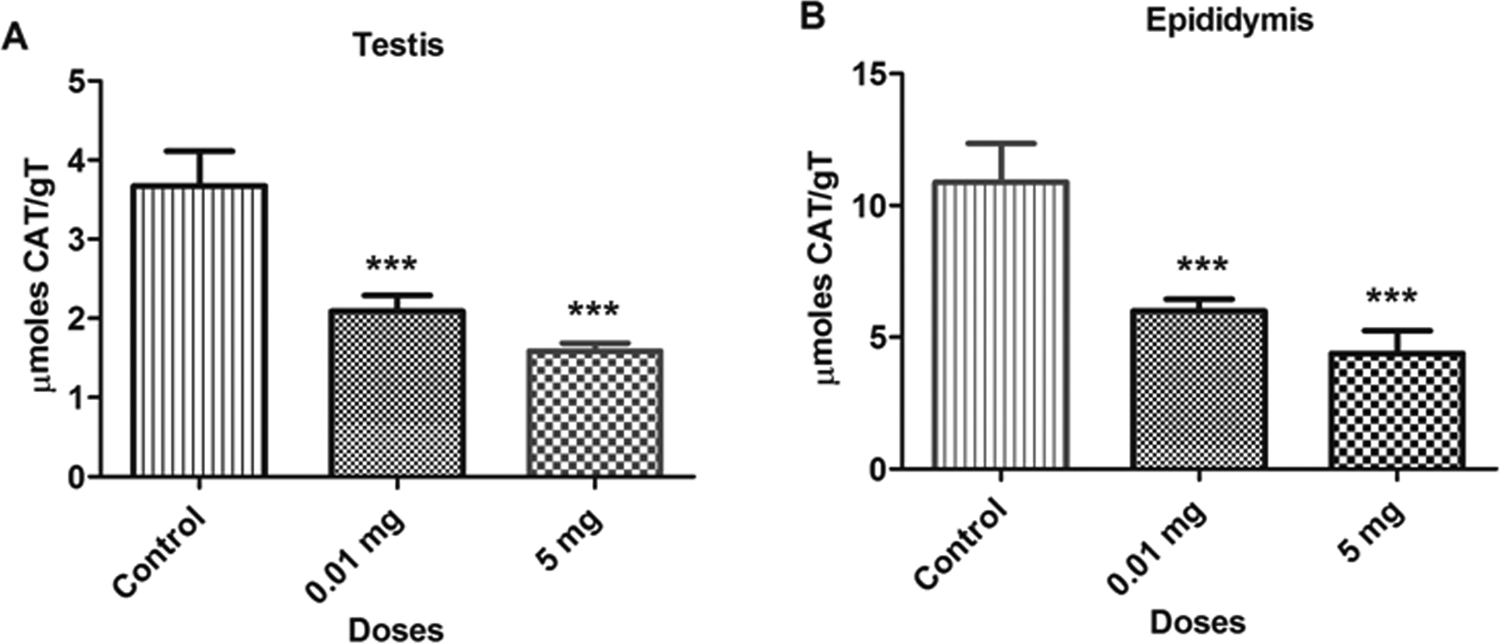
Bar diagrams showing a decrease in catalase activity between treated and control rats with the exposure to bisphenol A (BPA). A, Testicular tissue. B, Epididymal tissue. Data are shown as mean ± standard deviation (SD). ***P < 0.001. The significance level was set at P < 0.05.
Discussion
In the present study, we examined the effects of 2 different concentrations (0.01 and 5.0 mg/kg/bw) of BPA on various oxidative stress parameters in male rats. Bisphenol A was administered by oral gavage in rats over the period of 6 consecutive days. The BPA exposure induced the oxidative stress in bone marrow cells, lymphocytes, testis, and epididymis by decreasing the activity of enzymatic and nonenzymatic antioxidants, namely, SOD, CAT, and reduced GSH, whereas the levels of LPO increased significantly as compared to control animals.
Reactive oxygen species are produced as a result of normal cellular metabolism in living organisms. It has been observed that ROS perform its function in physiological cell processes at low to moderate concentrations. However, they produce adverse modifications to cell components, such as lipids, proteins, and DNA at high concentrations. 48 Aerobic organisms have a well-programmed antioxidant systems, which include enzymatic and nonenzymatic antioxidants. Antioxidants counteract the damaging effects of oxidation in body tissues by terminating oxidative chain reactions as a result of removing free radical intermediates and inhibit other oxidation reactions too. They counteract by donating electrons to the free radicals and oxidized themselves; hence, antioxidants are often reducing agents. 49 Regardless of the presence of this antioxidant system, excess production of ROS due to exposure of chemicals may result in a number of clinical disorders. Oxidative stress caused by excess ROS production is known to damage nucleic acid, lipid, and protein, resulting in the onset of various diseases, namely, cancer, infertility, and neurodegenerative diseases. 50 -53 Cell death, DNA mutation, replication errors, and genomic instability can occur if the oxidative DNA damage is not repaired prior to DNA replication. 54 -57
Oxidative stress has been proven as a basic mechanism of BPA toxicity in animal models for years. Bisphenol A have the potential to cause oxidative stress by disturbing the redox status in cells. 58 Kabuto et al revealed that injection of BPA induces overproduction of hydrogen peroxide in the mouse organs. 59 They also demonstrated a decrease in the levels of reduced GSH and an increase in the levels of oxidized GSH by hydroxyl radicals. It is of interest to note that BPA (0.05 and 1.2 mg/kg/d) exposure to mice for 5 days decreased the expression of GPX3 protein and increased the levels of MDA, a product of LPO, which leads to mitochondrial dysfunction and liver damage. 60 Mitochondrial membrane damage is an early event leading to cell death by apoptosis and may be initiated by LPO. 61 In many cases, the cellular defense network is overwhelmed by the oxidative attack leading to DNA damage.
The decreased levels of SOD in various samples caused by BPA exposure in our study may be due to the inability of the cells to generate enough SOD, which may arise from severe cellular damage or greater functioning in combating the oxidative stress. Superoxide dismutase protects tissues from oxidative stress and damage by catalyzing the conversion of superoxide anion radical into the less toxic hydrogen peroxide, a more stable ROS. 62 The decrease in SOD level can result in an increase in superoxide-free radical as well as an elevation of other ROS and intensification of LPO process. Catalase catalyzes the conversion of hydrogen peroxide, a more reactive ROS, into water, in this manner, providing protection against ROS. 55 The reduction in the activity of CAT may reflect the inability of cells to eliminate hydrogen peroxide generated in the cells. This may be due to enzyme inactivation caused by excess ROS production in cells. 63 In our study, we observed a significant decrease in CAT activity in BPA-administered rats as compared to control. Thus, the decrease in CAT activity in the cells increased the toxic effect due to excess of the free radical generation.
Reduced GSH is an antioxidant molecule and the most abundant intracellular nonprotein thiol present in cells. The GSH participates in catalytic cycles of several antioxidant enzymes such as GSH peroxidase and GSH reductase, which plays a very important role in reducing oxidative stress. 64 The reduction in GSH shows the failure of primary antioxidant system to act against oxidative stress. 65 Hence, the depletion of intracellular GSH is usually regarded as a measure of oxidative stress. In the present study, there was a decrease in the levels of GSH, which might be due to inability of cells to generate enough GSH, due to severe cellular damage or due to greater utility in combating the oxidative stress.
Lipid peroxidation is one of the main manifestations of oxidative damage initiated by ROS, and it has been linked to the altered membrane structure and enzyme inactivation as well as excessive damage of cellular macromolecules (protein, lipids, and nucleic acids), which has been shown to be a major contributor to the toxicity of contaminants. 66 It has been observed that increase in the level of LPO has been correlated with maturation arrest, decreased spermatozoa concentration, and morphology, as well as most notably motility due to alterations in the membrane potential. 67 -71 Increase in MDA level also positively correlate with DNA damage in spermatozoa. 72 -74 Further, the increased MDA level was caused by oxygen-free radical, which leads to midpiece abnormalities and decreased sperm counts. 75 Thus, the increased MDA levels were observed in our study due to the exposure of BPA in various samples which indicated an increase in generation of ROS, leading to high LPO activity and thereby enhancing the membrane disruption and DNA damage.
It is evident from the literature that BPA generates ROS by decreasing the activity of antioxidant enzymes and increasing LPO, thereby causing oxidative stress in liver and epididymal sperm of rats. 37,38 The results obtained in the present study corroborate with others who demonstrated that BPA can generate ROS that causes oxidative damage in the brain and liver of rats. 76 -78 The current study showed that BPA even at lower dose (0.01 mg) caused a significant increase in LPO, along with a decrease in the activity of SOD, CAT, and reduced GSH in bone marrow cells and lymphocytes. Earlier studies from our laboratory on genotoxicity of BPA have been documented that exposure of BPA to adult male and female rats below the NOAEL dose (5.0 mg) and even at 10 µg, which is very close to environmental exposure, led to a significant increase in the chromosome breaks and fragments in bone marrow cells as well as DNA fragmentation in blood lymphocyte. 35 The males also exhibited a significant increase in the levels of LPO and a decrease in reduced GSH in liver. In another study, we have also demonstrated that same dose of BPA caused a significant increase in the dominant lethal mutation rate and sperm DNA fragmentation in rats. 36
Oxidative stress is the major cause of DNA fragmentation in spermatozoa and is an important factor reported in the etiology of male infertility. Male germ cells are extremely vulnerable to oxidative stress as the sperm membrane is rich in unsaturated fatty acids 79 and lacks the capacity for DNA repair. 80,81 To counteract the effects of ROS, the testes are equipped with antioxidant defense systems that prevent cellular damage. 82 In the present study, we observed that BPA exposure enhanced a significant increase in LPO that was accompanied by a significant decrease in reduced GSH, SOD, and CAT levels in the testis and epididymis. At the level of the testes, oxidative stress is capable of disrupting the steroidogenic capacity of Leydig cells 83 as well as the capacity of the germinal epithelium to differentiate normal spermatozoa. 84 This effect of BPA might be the mechanism by which BPA-induced testicular toxicity in rats and mice can be explained. 85,86
The urinary BPA concentrations were positively associated with MDA, 8-OHdG, and CRP levels in the postmenopausal women. The findings of this study suggest that BPA exposure would promote oxidative stress and inflammation, in which postmenopausal women are likely to be more susceptible to BPA-induced health effects. 87 Recently, it has been reported that BPA exposure significantly induced DNA damage in zebra fish, which is associated with a significant increase in ROS production. 88 They also demonstrated that BPA induced both necrotic and apoptotic cell death in whole-mount zebra fish by a dose-dependent manner because of excess ROS-induced oxidative damage of cellular macromolecules promoting cell death. Moreover, BPA has also been reported to enhance oxidative stress and LPO by disturbing the redox status in cells promoting the cellular death in brain of exposed rodents. 89
Although the exact molecular mechanisms of action of BPA on the production of ROS is still remains unclear. However, a wide variety of scientific evidence supports the hypothesis that natural estrogens, synthetic estrogen diethylstilbestrol, and BPA generate ROS during biotransformation and that certain ROS, predominantly quinones, a reactive metabolites which can react with DNA and cause DNA damage. 90 Based on the results of the present study and the considerable body of literature on effects of BPA at similar and much higher doses, it was suggested that BPA induced oxidative stress, which could be one of the possible mechanisms for genotoxic activity of BPA.
In summary, the results of the present study revealed that both low and high doses (0.01 and 5.0 mg/kg/bw) of BPA exposure generate excess of ROS by decreasing the levels of SOD, CAT, reduced GSH and increasing LPO in bone marrow cells, blood lymphocyte, and reproductive organs, thereby causing oxidative stress. The merit of this study is that BPA led to the induction of oxidative stress in bone marrow cells, blood lymphocytes, and testicular and epididymal tissues of treated rats even at environmentally relevant dose. The present study also suggests that BPA induced oxidative stress, which could be one of the possible mechanisms of BPA-induced genetic damage in the somatic and germ cells of male rats resulting in reproductive and genetic toxicity.
Footnotes
Acknowledgments
The authors are grateful to Dr Vikas Dighe, Dr Rohit Dhumal, Dr Madhuwanti Mahajan, Dr Tanvi Doshi, Sharad Bhagat, and Mrs Annette Fonseca for helpful and valuable discussion. The authors are also grateful to Jayant Tare, Pravin Salunke, Subhash Kadam, and Maruti Mali for helping in animal experiments. The authors would also like to thank Mr Vaibhav Shinde for giving his technical guidance while preparing the artwork of manuscript.
Author Contributions
G. Vanage contributed to conception and design, acquisition and analysis, drafted the manuscript, and critically revised the manuscript. D. Tiwari contributed to conception, acquisition, drafted the manuscript, and critically revised the manuscript. Both authors gave final approval and agree to be accountable for all aspects of work ensuring integrity and accuracy.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by Department of Science and Technology and Indian Council of Medical Research (VII-PRDSF/101/05-06/TDT).