
Abstract
The metabolism of yellow cassava (variety TMS 01/1368) was investigated in male albino rats fed a diet containing yellow cassava for 7 to 28 days. There were significant increases (P < 0.05) in total and free cyanide and thiocyanate in the sera and urine samples of the experimental rats compared with the control, significant increases (P < 0.05) in serum glucose, alanine aminotransaminase, aspartate aminotransaminase, and alkaline phosphatase levels of the experimental rats compared with the control, significant decreases (P < 0.05) in serum albumin of the experimental rats compared with the control, but no significant differences (P > 0.05) in the serum total proteins of the experimental rats compared with the control. The experimental rats treated for 7, 14, 21, or 28 days exhibited body weight decreases of 5.11%, 11.10%, 19.16%, and 24.18%, respectively, whereas the control group showed 9.17% gain in body weight. Total and free cyanide concentrations were detected in the liver, kidney, and heart of most of the rats in both the experimental and control groups, except for free cyanide in the control group that was not detected. Metabolism of the yellow cassava variety in experimental rats was capable of exposing the animals to cyanide, underscoring the need for its proper processing before consumption by humans.
Introduction
Cassava (Manihot esculenta Crantz) is one of the most important food crops in the tropical countries and is probably the most widely distributed human crop with high content of cyanogenic glycosides. 1 Cassava roots form important staple foods for more than 500 million people, particularly in tropical countries in Africa, Asia, and Latin America. 2 It is estimated that the crop provides about 40% of all calories consumed in Africa. 3
The use of cassava and its byproducts for food consumption or as feed is strongly influenced by the presence of toxic cyanogenic glycosides, which can lead to cyanide poisoning with symptoms of vomiting, nausea, dizziness, stomach pains, weakness, headaches, diarrhea, and in extreme cases, death. 4,5 Consumption of a monotonous diet of high-cyanide cassava by poor rural people in Africa, many of whom have only 1 meal per day and have malnutrition, was associated with the outbreak of konzo, an upper motor neuron disease that causes irreversible paralysis of the legs and occurs mainly in children and young women of childbearing age. 6 Cyanide intake from cassava was also reported to cause goiter and cretinism in iodine-deficient areas. 7,8
These metabolic disorders arising from consumption of cassava occur due to the release of hydrogen cyanide from cassava cyanogenic glucosides. 9,10 The major site of action of cyanide in biological systems is at the cytochrome oxidase in the electron transport chain where it binds to this enzyme, thereby arresting tissue respiration. The major pathway for detoxification of cyanide involves rhodanese (thiosulfate–cyanide sulfurtransferase), which catalyzes the transfer of sulfur from thiosulfate to cyanide forming thiocyanate, 11 as shown in the equation below:
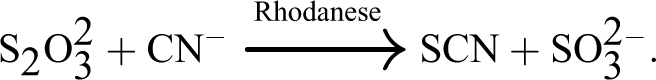
The goitrogenic effect of cassava consumption that causes enlargement of the thyroid gland results from the formation of thiocyanates. 12 Alternative pathway for cyanide detoxification provided by cyanocobalamin from hydroxyl cobalamin has also been reported. 13,14
Recently, National Root Crops Research Institute, Umudike, Abia State, Nigeria, released some yellow cassava varieties fortified with β-carotene as a way of combating vitamin A deficiency in Nigeria. Although the carotenoid contents of these cassava varieties have been reported, 5 there is no information available in the literature on the metabolic effects of consumption of yellow cassava varieties (fortified with β-carotene) in humans or experimental animals.
This study therefore sought to investigate the metabolic effects of consumption of the yellow cassava variety TMS 01/1368, also known as UMUCASS 36, on some biochemical parameters in experimental rats.
Materials and Methods
Plant Materials
Fresh roots of the yellow cassava variety (TMS 01/1368) were obtained at harvest from the experimental farm of National Root Crops Research Institute, Umudike, Abia State, in 2013. It was peeled, washed, sliced into small pieces, sun dried, processed to flour of particle size about 2 mm, using an electrical blender, and fed to experimental rats in a dietary formulation consisting of 77% cassava flour, 9% soybean, 4% vitamin mixture, 4% banana flavor, 4% groundnut oil, and 2% salt. 15 Proximate analysis of the flour of this cassava variety on a dry weight basis (9.85% moisture) indicated that it contained on the average 2.02% crude fiber, 1.38% fat, 2.35% ash, 5.67 µg/g carotene (fresh weight), and 2.57 g/100 g reducing sugar. 5
Animal Experiments
Selection of animals
Twenty-five male albino rats of the Wistar strain (weighing 150-165 g), bred in the animal house of University of Nigeria, Nsukka, were used. All animals were kept at room temperature (25°C) and had free access to drinking water and their diets. The animals were acclimatized to their environment, experimental control diets, and water for 7 days before experiments were commenced. All animal protocols were approved by the Ethical Committee of Michael Okpara University of Agriculture, Umudike, Nigeria, which was consistent with the guide for the care of use of laboratory animals as reported by Kilkenny et al. 16
Animal feeding
The 25 rats were divided into 5 groups of 5 rats each. Groups 1, 2, 3, and 4 (experimental groups) were fed daily with yellow cassava diet, whereas group 5 rats, which served as the control, were fed standard rat pellets (cassava-free diet) comprised 19% protein, 10% fat, 31% carbohydrate, 10% crude fiber, and 1% calcium with a food energy value of 290 kcal/100 g.
Experimental procedure
The body weights of the rats were measured on a daily basis, using an electronic weighing balance (Model-Scout Pro; Ohaus Corporation, USA). Animals in groups 1, 2, and 3 were euthanized on the 7th, 14th, and 21st days, respectively, whereas those in groups 4 and 5 were euthanized on the 28th day. Each animal was euthanized after overnight fast, and blood was collected directly from the hearts using 10-mL syringes and poured into nonanticoagulant tubes for serum assay of glucose, total and free cyanide, thiocyanate, alanine aminotransaminase (ALT), aspartate aminotransaminase (AST), alkaline phosphatase (ALP), and total proteins. The livers, kidneys, and hearts of the rats were harvested immediately, washed with normal saline, and stored at −20°C until use. Urine samples from the rats from each group were collected before the rats were euthanized and analyzed for total cyanide, free cyanide, and thiocyanate.
Preparation of tissues for cyanide analysis
The livers, kidneys, and hearts of the animals in each group were homogenized in 0.1 mol/L orthophosphoric acid buffer. The homogenates were centrifuged at 3000g for 10 minutes, and the supernatants were analyzed for total and free cyanide.
Plant Assay
Determination of total and free cyanide content of the cassava-based diet
The yellow cassava diet (50 g) was dissolved in 200 mL of 0.1 mol/L orthophosphoric acid buffer medium. The mixture was filtered, and the filtrate was kept in the refrigerator and analyzed within 24 hours. The total cyanide content of the diet was determined by phosphoric acid extraction, hydrolysis of cyanogenic glucosides with linamarase from cassava, followed by colorimetric determination of cyanide. Free cyanide was determined in a similar way but without the use of linamarase. 17
Animal Assays
Biochemical analysis in sera/urine
The thiocyanate concentrations in the sera and urine samples of the rats were determined using the ferric nitrate reagent method as described by Sorbo. 18 The AST and ALT concentrations in the sera of the rats were determined using the methods of Reitman and Frankel. 19 Serum concentrations of ALP were determined using the methods of Kochmar and Moss. 20 The glucose concentrations in the sera of the rats were determined using the glucose oxidase method as described by Barham and Trinder. 21 Serum total protein was determined using the Biuret method as described by Tietz. 22 Serum albumin was determined using the dye-binding (bromocresol green) method as described by Doumas et al. 23
Total cyanide assay in tissues
Aliquots of sample (1 mL) were pipetted into different test tubes. A measured quantity (0.1 mL) of linamarase enzyme was added, followed by incubation in a water bath at 40°C for 30 minutes. They were allowed to cool for 15 minutes. Thereafter, 0.1 mL of 0.2 mol/L NaOH was added, followed by the addition of 2.8 mL of phosphate buffer (0.1 mol/L, pH 6.0), 0.2 mL of chloramine T, and 0.8 mL of color developer (which consisted of 3.7 g NaOH, 7 g of 1,3-dimethylbarbituric acid, and 5.7 g of isonicotinic acid dissolved in 200 mL of distilled water). They were allowed to stand for 20 minutes and read in spectrophotometer at 605 nm. 17
Free cyanide assay in tissues
Aliquots of sample (1 mL) were pipetted into different test tubes, followed by addition of 2.8 mL of phosphate buffer, 0.2 mL of chloramine T, and 0.8 mL of color developer. They were allowed to stand for 20 minutes and read in spectrophotometer at 605 nm. 17
The total and free cyanide concentrations in the tissues were quantified from a potassium cyanide standard curve.
Data Analysis
The statistical package for social sciences (SPSS) (version 17.0; SPSS Inc, Chicago, Illinois) was used to analyze all data. Results are presented as means ± standard deviation. One-way analysis of variance was used for comparison of means. Differences between means were considered to be significant when P < 0.05.
Results
The total and free cyanide concentrations of the cassava-based diet used in this study were 51.13 ± 0.08 mg/kg and 8.06 ± 0.05 mg/kg, respectively. There was a significant elevation (P < 0.05) in the total cyanide and thiocyanate concentrations in the sera of the rats in the experimental groups compared with the control (Table 1). There were detectable levels of free cyanide in the sera of the experimental rats in the respective groups fed the test diet but nondetectable levels of free cyanide in the sera of the rats in the control group (Table 1).
Total Cyanide, Free Cyanide, and Thiocyanate (SCN) Levels in the Serum of Rats.a
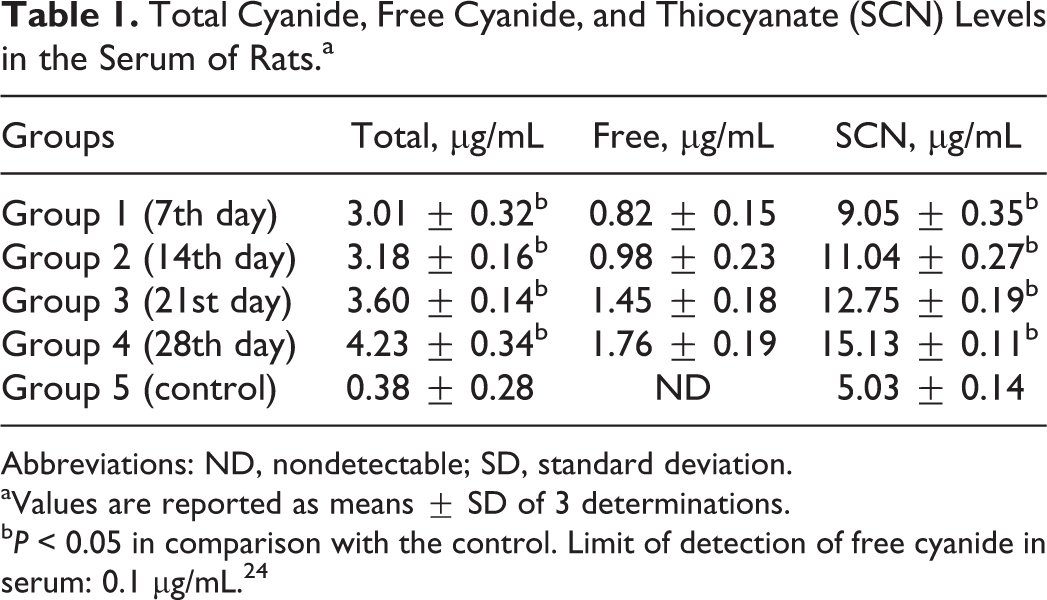
Abbreviations: ND, nondetectable; SD, standard deviation.
aValues are reported as means ± SD of 3 determinations.
b P < 0.05 in comparison with the control. Limit of detection of free cyanide in serum: 0.1 µg/mL. 24
There was a significant elevation (P < 0.05) in the total cyanide, free cyanide, and thiocyanate concentrations in the urine samples of the rats in the experimental group compared with the control (Table 2).
Total Cyanide, Free Cyanide, and Thiocyanate Levels in the Urine Samples of Rats.a
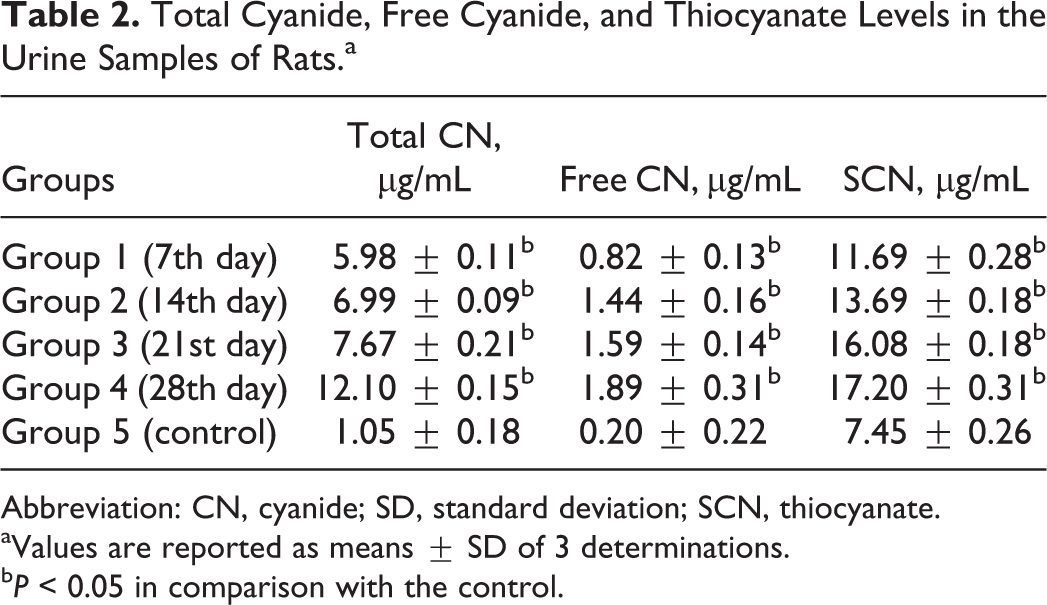
Abbreviation: CN, cyanide; SD, standard deviation; SCN, thiocyanate.
aValues are reported as means ± SD of 3 determinations.
b P < 0.05 in comparison with the control.
Significant elevation (P < 0.05) in the serum glucose concentrations of the rats was observed in the experimental group compared with the control (Table 3).
Glucose, Total Protein, and Albumin in the Sera of Rats.a
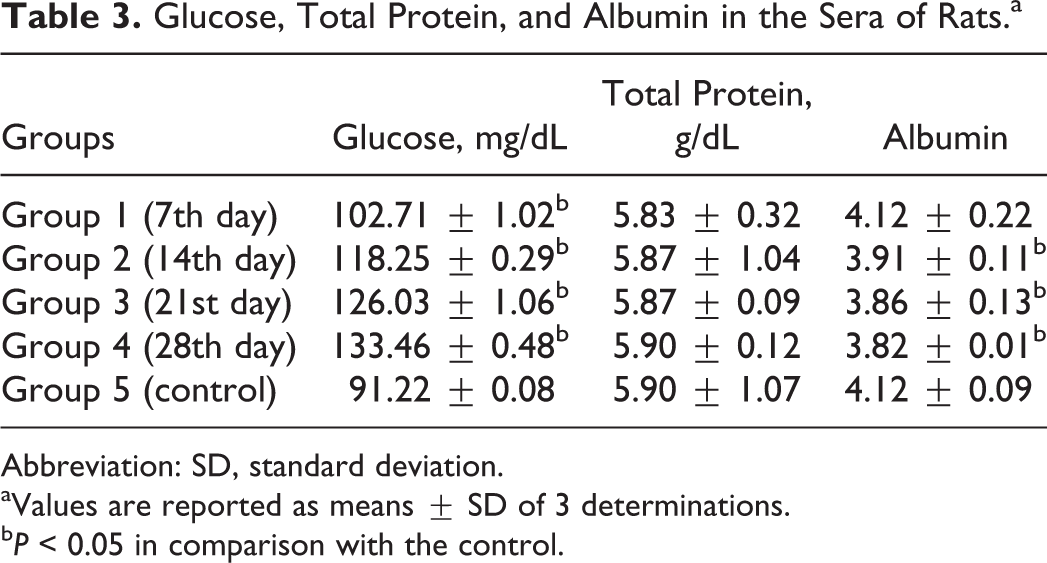
Abbreviation: SD, standard deviation.
aValues are reported as means ± SD of 3 determinations.
b P < 0.05 in comparison with the control.
There were no significant differences (P > 0.05) in the concentrations of total proteins in the sera of the rats in the experimental group compared with the control (Table 3).
There was no significant difference (P > 0.05) in the serum albumin concentration of the rats fed the test diet for 7 days compared with the control, whereas there were significant reductions (P < 0.05) in the serum albumin concentrations of the rats fed the test diet for 14, 21, or 28 days compared with the control (Table 3).
The rats fed the test diets for 7, 14, 21, and 28 days had 5.11%, 11.10%, 19.16%, and 24.18% decreases in body weights, respectively, compared with the control that recorded 9.17% gain in body weight (Table 4).
Body Weights and Percentage Changes in Rats.a
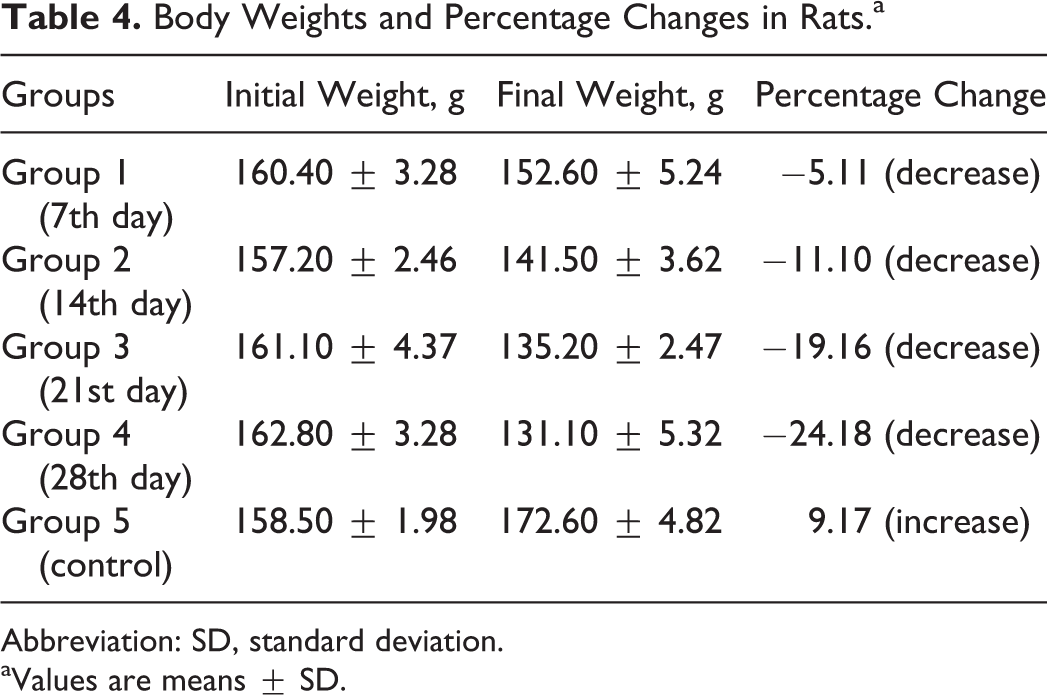
Abbreviation: SD, standard deviation.
aValues are means ± SD.
There were significant increases (P < 0.05) in the total and free cyanide concentrations in the liver and kidneys of the rats in the experimental group compared with the control (Tables 5 and 6).
Total and Free Cyanide Concentrations in the Livers of Rats.a
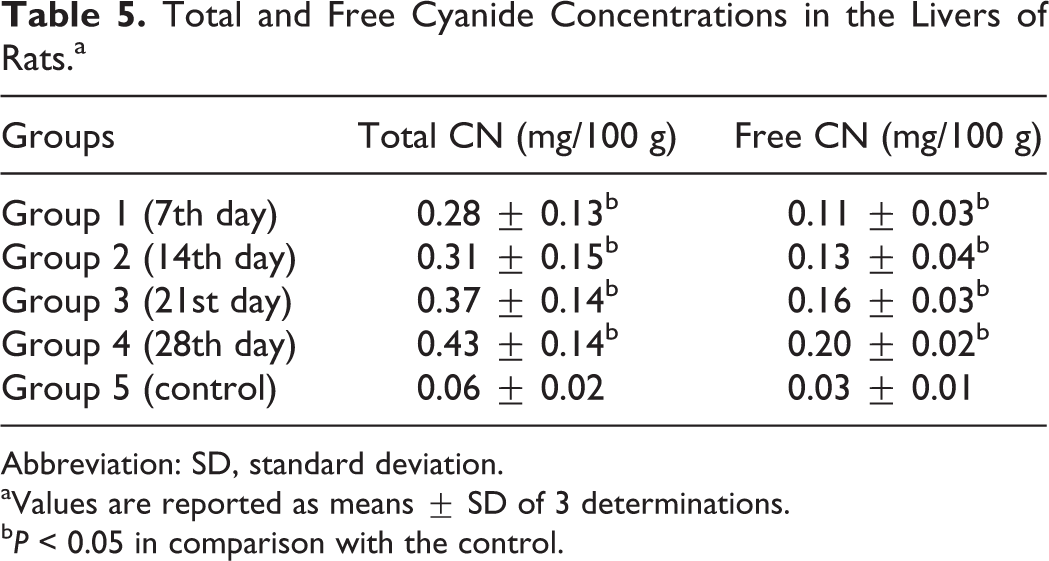
Abbreviation: SD, standard deviation.
aValues are reported as means ± SD of 3 determinations.
b P < 0.05 in comparison with the control.
Total and Free Cyanide Concentrations in the Kidneys of Rats.a
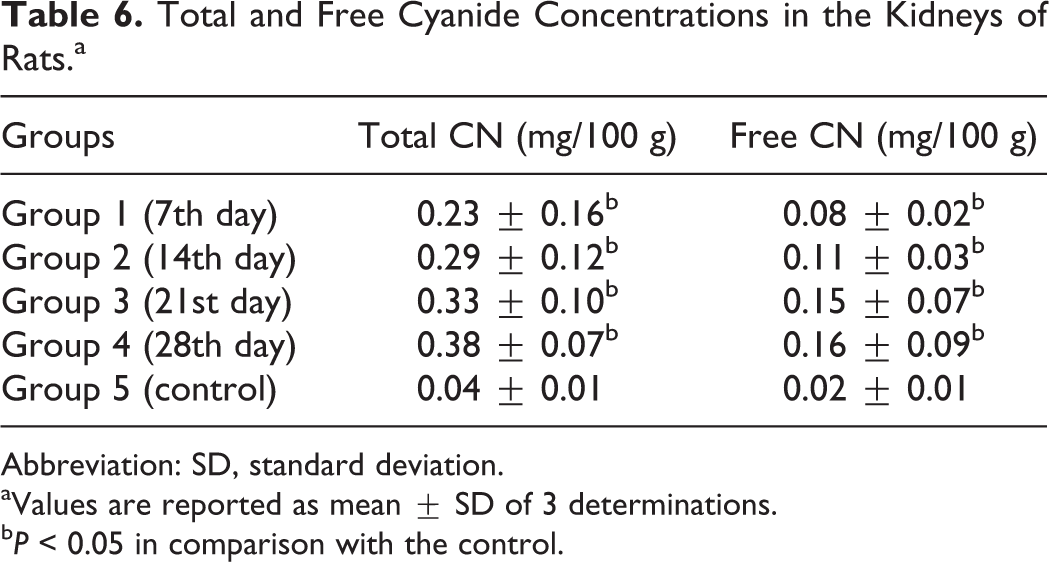
Abbreviation: SD, standard deviation.
aValues are reported as mean ± SD of 3 determinations.
b P < 0.05 in comparison with the control.
There were significant increases in the total cyanide concentrations in the hearts of the rats fed the test diet for 7, 14, and 28 days, respectively, compared with the control, no significant differences (P > 0.05) in the total cyanide concentration in the hearts of the rats fed the test diet for 21 days compared with the control, and detectable levels of free cyanide in the heart of the rats in the experimental group but nondetectable levels of free cyanide in the heart of the rats in the control group (Table 7).
Total and Free Cyanide Concentrations in the Hearts of Rats.a
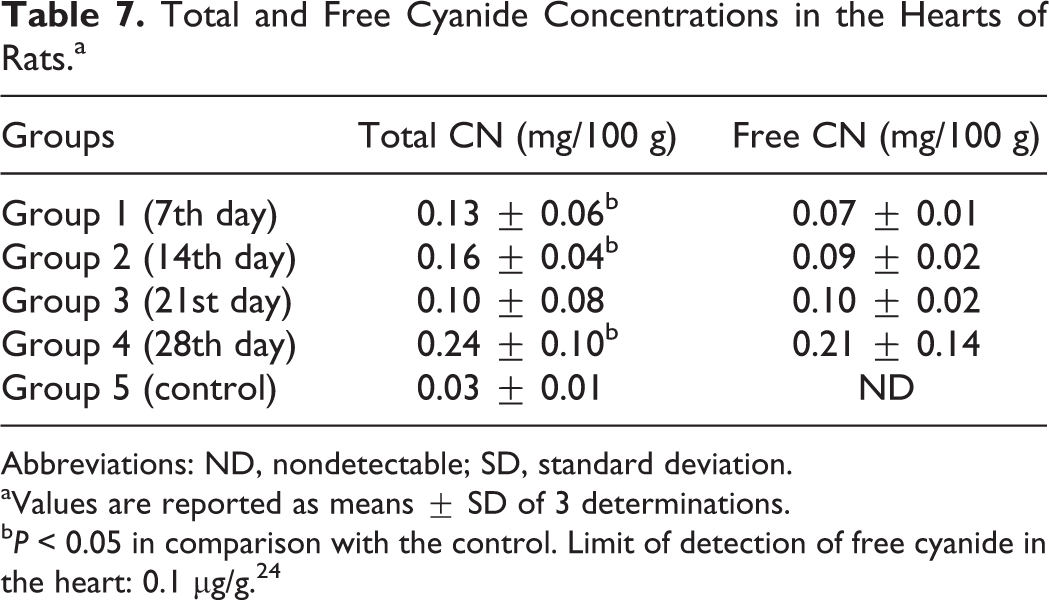
Abbreviations: ND, nondetectable; SD, standard deviation.
aValues are reported as means ± SD of 3 determinations.
b P < 0.05 in comparison with the control. Limit of detection of free cyanide in the heart: 0.1 µg/g. 24
As shown in Table 8, there were significant increases in the serum ALT, AST, and ALP of the rats in the experimental group compared with the control.
Liver Function Parameters in the Sera of Rats.a
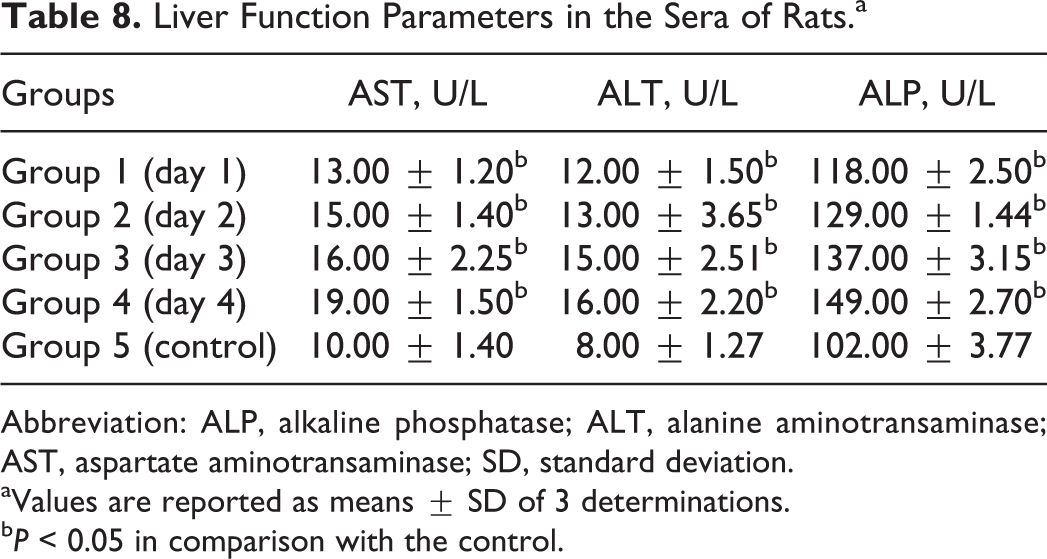
Abbreviation: ALP, alkaline phosphatase; ALT, alanine aminotransaminase; AST, aspartate aminotransaminase; SD, standard deviation.
aValues are reported as means ± SD of 3 determinations.
b P < 0.05 in comparison with the control.
Discussion
During processing of cassava, the cyanogenic glucosides (linamarin and lotaustralin) are degraded by linamarase, an intrinsic enzyme present in cassava. 1 Further degradation of the glucosides could occur as a result of microbial action. However, some of the glucosides still escape breakdown and are therefore ingested by consumers. Although mammalian intestines contain β-glucosidases and microorganisms that are capable of hydrolyzing these glucosides, it has been established that these intestinal microorganisms play no significant role in the breakdown of linamarin, the main cyanogenic glucoside of cassava. 25 Therefore, the β-glucosidases present in the brush boarders of the intestinal mucosa do not constitute a great barrier to the passage of intact cyanogenic glucosides. This is evidenced by the fact that bound cyanide is often detected in the blood and urine following consumption of cassava and other cyanogenic foods. 15 Thus, following consumption of cassava and its products, linamarin, lotaustralin, acetone cyanohydrin (partially breakdown product of linamarin), and hydrogen cyanide are present in the serum and urine of the consumer.
Therefore, the serum levels of cyanide in the experimental rats, which were higher than the reported toxic levels in rats (2.60-2.92 μg/mL), 26 suggest exposure of the rats to cassava cyanide intoxication.
Elevation of total and free cyanide in the urine samples of the rats in the experimental group compared with the control group further affirms the exposure of these animals to cyanide toxicity.
Thiocyanate remains the most reliable biomarker for cyanide exposure, being a stable metabolite of its detoxification. 27,28 Elevation of thiocyanate in the sera and urine samples of the rats in the experimental group compared with the control group could be an indication of an attempt by the experimental rats to detoxify the ingested cyanide.
Cyanide is known to alter glucose metabolism resulting in increased glucose and lactic acid levels and a decrease in the adenosine triphosphate–adenosine diphosphate ratio, indicating a shift from aerobic to anaerobic metabolism. Cyanide apparently activates glycogenolysis and shunts glucose to the pentose phosphate pathway, decreasing the rate of glycolysis and inhibiting tricarboxylic acid cycle. 29
Decreased levels of serum total proteins and albumin in the experimental rats suggest use of the sulfur-containing amino acids (cysteine and methionine) of their body to detoxify the ingested cyanide leading to decreased liver synthesis. 30 This is especially important as serum proteins, especially serum albumins are involved in cyanide intoxication. 24
The decreases in the body weights of the rats in the experimental group could arise from the mobilization of endogenous sulfur from the breakdown of body proteins of sulfur amino acids utilized in cyanide detoxification, thus reducing the quality of the proteins that are used in the growth-promoting process of the body.
The presence of cyanide in the tissues may be due to the fact that following absorption, cyanide is rapidly distributed throughout the body by the blood, 31 and since it is lipid soluble, 31 it passes through the highly perfused organs such as the liver, kidney, and heart, respectively, resulting in its detection in these organs. This may also explain the detection of cyanide in the highly perfused organs such as the brain, kidney, heart, and liver following oral exposure to cyanide as reported previously. 32
Measurement of enzymic activities of aminotransaminases (AST and ALT) and phosphatases (ALP) is of clinical and toxicological importance, as increases in the levels of these diagnostic enzymes could indicate hepatic cytoplasmic and/or mitochondrial membrane damage even if there is no detectable impairment of function. 33 Okafor et al 24 in their studies reported increased AST and ALT concentrations in the sera of women frying gari (a doe from cassava processing). The observed increase in the serum levels of these diagnostic enzymes in the rats in the experimental group compared with the control group suggests damage to the tissues as a result of cyanide ingestion.
A widely adopted traditional method that is used by different countries in Africa and other countries of the world to properly process cassava before ingestion involves peeling, washing, cutting into chips, grating to mash, dewatering in clean bags, disintegration of the lumps to particle sizes of about 3 mm, sun drying, and milling to flour. Cyanogens could further be removed from the flour using the wetting method, 33 whereby the dry flour is added to a bowl and the level marked on the inside of the bowl. Water is then added with mixing, and no more water is added when the level of the wet flour comes up to the mark. The wet flour is then placed in a thin layer not greater than 1 cm thick on a mat and allowed to stand for 2 hours in the sun or 5 hours in the shade for the hydrogen cyanide gas to escape.
In conclusion, the study indicated that metabolism of the cassava diet (formulated from the yellow cassava variety, TMS 01/1368) in experimental rats was capable of exposing the animals to cyanide intoxication. The study therefore underscores the need to properly process this cassava variety before consumption by humans.
Footnotes
Author Contributions
All authors contributed equally to this manuscript. Nelson Udeme contributed to conception and design, contributed to acquisition, analysis, or interpretation of data, drafted the manuscript, critically revised the manuscript, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. Polycarp Okafor contributed to conception and design, contributed to acquisition, analysis, or interpretation of data, drafted the manuscript, critically revised the manuscript, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. Chinedum Eleazu contributed to conception and design, contributed to acquisition, analysis, or interpretation of data, drafted the manuscript, critically revised the manuscript, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy.
Acknowledgments
The authors wish to thank the management of NRCRI, Umudike, for supplying the cassava variety used in this study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.