
Abstract

Dead cells do not mutate. This is not a new concept. A corollary to this is that cell toxicity is antimutagenic; any substance or procedure that causes cells to die or prevents them from replicating, regardless of its mechanism of action, will reduce the number of mutant cells and may wrongly lead to the conclusion that the substance is an antimutagen. However, although death is antimutagenic, it is not therapeutic.
Antimutagens are sought for practical reasons. We are exposed to mutagenic substances through the air, water, foods, industrial chemicals, and drugs. These mutagens are of concern not only because they can cause mutations in our germ cells that may be passed to our offspring but also because somatic cell mutagens have a high association with cancer induction. Most of these mutagens cannot be removed without destroying/ruining the products they are in or their removal will lead to greater immediate health risks than mutagenicity or carcinogenicity, for example, mutagenic antitumor agents, or water disinfection by-products. Therefore, the best protection against them would seem to be ingestion of substances that could prevent or counteract the effects of the mutagens—antimutagens.
The past 20 years have seen a surge in publications aimed at identifying antimutagens 1 (and potential anticarcinogens) using in vitro and in vivo tests. The primary impetus for these studies is based on the prospect of cancer from somatic cell mutation and the reduction of cancer risk from environmental chemical exposures. Although many discrete substances have been tested, a large proportion of the published studies deal with uncharacterized extracts of plants. The underlying premise is that natural products, because they are natural rather than produced by chemists, are healthful and protective, ignoring the fact that strychnine, ricin, and, coniine (found in the classical poison, hemlock) are also natural products.
In addition to plant extracts, substances reported as antimutagenic have included inorganic salts, vitamins, pharmaceuticals, industrial chemicals, microbes and microbial extracts, animal extracts and products, and even human tissue extracts and products. In the case of plant extracts, every possible part of the plant has been homogenized, extracted, and studied: entire plant, roots, stems, bark, leaves, flowers, seeds (nuts; fruit), and thorns.
Substances may act as antimutagens via a number of nonexclusive mechanisms, including:
interception and inactivation or sequestration of the mutagen, interference with the metabolic activation of the mutagen, or enhancement of its deactivation, modulation of DNA damage repair so that it is less error prone or leads to a lethal, rather than a mutagenic event, depression of the cell cycle, interference with cell reproduction, or death.
In their zeal to identify antimutagens, many authors often appear to lose their sense of perspective for hypothesis testing and often identify substances as antimutagenic and recommend animal anticancer studies, regardless of the data.
This commentary was inspired by a recently published manuscript that purports to demonstrate the antimutagenicity, in the presence of cell toxicity, of a number of uncharacterized plant extracts using the Ames Salmonella mutagenicity test. The purpose of the commentary, however, is not to minimize the study of antimutagenesis and identification of antimutagens but to address a number of study design deficiencies and inconsistencies in data interpretation that have been identified in published antimutagenesis manuscripts, and manuscripts submitted for publication and at meeting presentations. Three areas, specifically, are addressed: (a) the need to identify and characterize the toxicity of the test substance and the combination of test substance and reference mutagen; (b) the situation where the test substance can be both mutagenic and antimutagenic depending on the test concentrations and test protocol; and (c) the need to chemically characterize the substance being studied. Examples are presented to illustrate the various situations.
The goal of this commentary is not to single out individual authors or publications, so citations are not presented. The examples presented here were taken directly, or adapted, from published articles and presentations, or from manuscripts reviewed by the author. The relevant citations were provided to the Editor of International Journal of Toxicology to allow the data and statements to be verified.
Cell Toxicity
The majority of antimutagenesis studies fail because either the toxicity of the test substance is not determined or the toxic effect of the combination or co-treatment of test substance and reference mutagen is not measured. This has been seen in studies with plant and other extracts and also in studies using pure chemicals. In the more extreme cases, toxicity is seen but is ignored when interpreting the data. Toxicity is the most important consideration because it goes to the basis of the claim that a specific substance or treatment will lower the mutation frequency or prevent the induction of mutations.
Chemicals do not cause mutations. The cell causes the mutations when it attempts to replicate its DNA in the presence of the chemical damage, or an error is introduced by the cell when it attempts to repair the damage. In order for a mutation to be fixed and expressed, the cell must divide or, at the least, DNA replication must take place. Similarly, cell stasis, that is the cells are alive but reproduction does not occur or the reproductive rate is dramatically slowed, will not allow mutations to be fixed and expressed in the daughter cells. One recently published example of the relationship of toxicity to mutation reduction is seen in Table 1.
Comparison of Plant Extract Toxicity to Salmonella and Inhibition of Mutation a

aAdapted from a published data table. Percentage inhibition values are from the publication and represent the decreases in mutant counts in the presence of extract + reference mutagen. The percentage toxicity values were calculated for this table and are based on the mutant counts at different extract concentrations in the absence of reference mutagen.
In this example, the authors show that addition of the test extract in the presence of potent mutagens decreased the number of mutant colonies of Salmonella strains TA98 and TA100. The decreases in both strains were taken as evidence for “potent antigenotoxic factors” in the extract. However, although the presented data clearly show decreases in mutant colony numbers with increasing concentrations of extract, the same data tables also show increases in cell toxicity with increasing extract concentrations. Despite the decrease in mutant numbers in the presence of increasing concentrations of extract, the manuscript claimed that the extract was not toxic by itself, and there was no indication that the toxicity of the extract-plus-mutagen combination was measured. This toxic response, although included in the data presentation, was not calculated or addressed in the publication. Although the percentage toxicity does not track exactly with the percentage inhibition, that is, in TA98 the extract is more inhibitory than toxic, whereas the opposite is true in TA100; the toxicity of the extract, alone, in the absence of a measure of extract + mutagen toxicity, brings into question the conclusion that the extract is an antimutagen.
The examples in Tables 2 and 3 are adapted from unpublished in vitro and in vivo cytogenetic studies using a different plant extract. In the in vitro experiment using mammalian cells (Table 2), the plant extract, alone, and the mutagen alone, are toxic as measured by the decreased mitotic index (MI), and the combination of mutagen and extract is even more toxic. If the more chemically damaged cells cannot enter into mitosis, the chromosome aberrations will not be formed. The same plant extract produced similar effects in mice in vivo (Table 3). The proportion of bone marrow reticulocytes (RET) is not affected by the extract or mutagen alone but is significantly reduced in the presence of the combination. In both these examples, the toxicities of the individual substances or the combination call into question whether the chromosome damage responses seen are a function of an antimutagenic effect or simply a toxic response.
Decrease in Chromosome Aberration (ABS) Frequency in Mammalian Cells In Vitro With Increasing Cell Toxicity a

Abbreviations: MI, mitotic index; Freq, frequency.
a Adapted from data tables from an unpublished manuscript. Only 1 concentration of each mutagen and the plant extract was tested.
b Significantly decreased clastogenic response to untreated control, or
c relative to the mutagen, alone.
Decrease in Micronucleus (MN) Frequency in Mouse Bone Marrow Reticulocytes (RET) In Vivo With Increasing Bone Marrow Cell Toxicity a
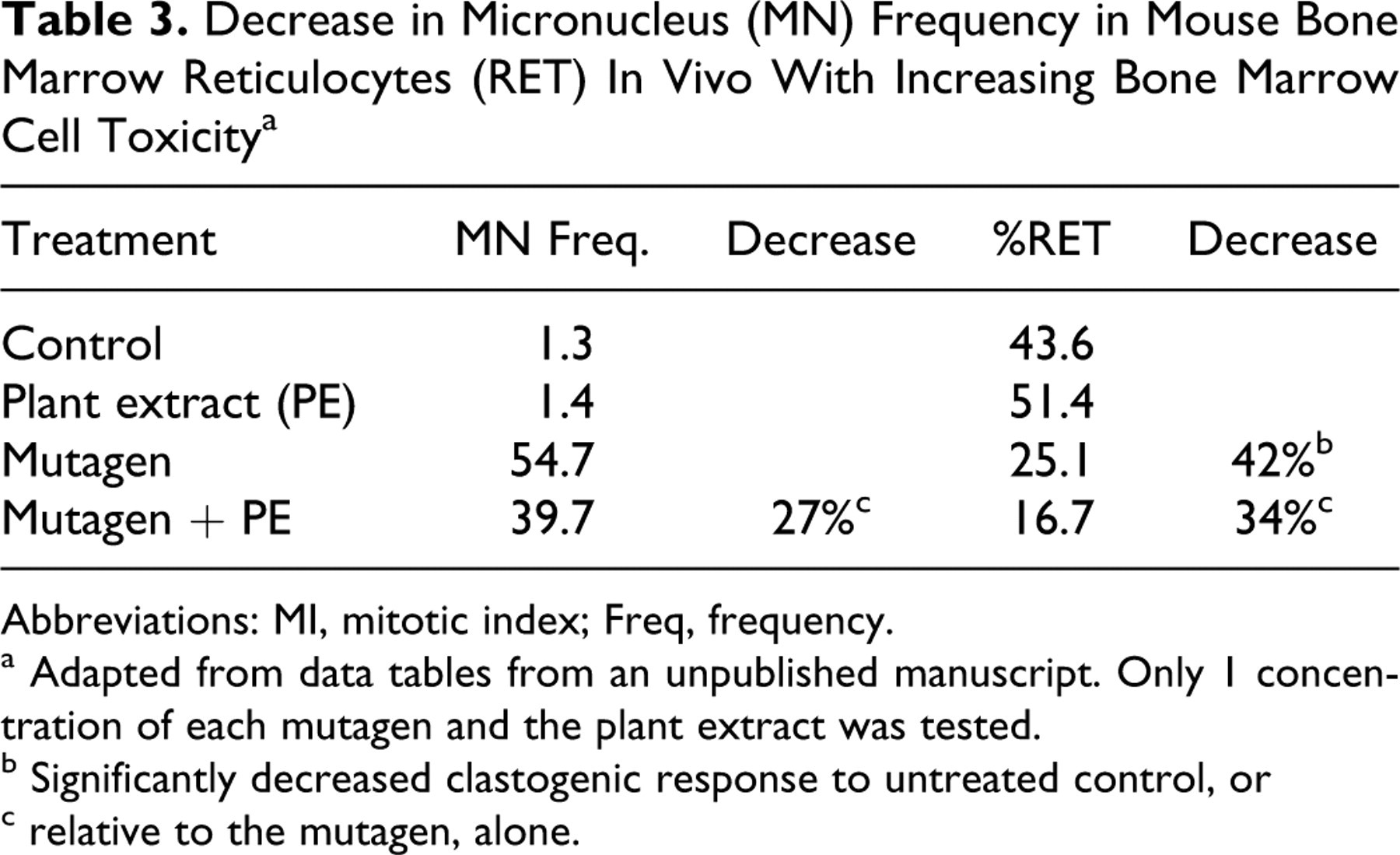
Abbreviations: MI, mitotic index; Freq, frequency.
a Adapted from data tables from an unpublished manuscript. Only 1 concentration of each mutagen and the plant extract was tested.
b Significantly decreased clastogenic response to untreated control, or
c relative to the mutagen, alone.
Another important consideration is where on the mutagenicity concentration-response curve the reference mutagen’s response falls—is it on the logarithmically increasing section or on the section where the response levels off or just short of toxicity. If the concentration of the mutagen is at or near the stationary or toxic level, a slight increase in toxicity from the addition of the test substance could lead to a decreased survival, with a parallel decrease in mutant colonies. Unless otherwise demonstrated, the decrease in mutant cells in the presence of the test substance would be the result of toxicity or the saturation or loss of activity of the S9-activating enzymes, rather than an antimutagenic effect. One way to avoid this phenomenon is to use the reference mutagen at concentrations on the logarithmically increasing portion of the mutation response curve, where the response would be most sensitive to increases or decreases in activity, thereby lessening the possibility that small changes are the result of a toxic effect.
Antimutagens That Are Also Mutagens
A search of the literature will identify a number of chemicals that have been reported as both mutagenic and antimutagenic (Table 4). For some of these, the authors have noted the presence of a mutagenic response under certain conditions or an increase in mutant numbers in one tester strain of Salmonella and an equivalent decrease in mutant numbers in another strain but, in keeping with their hypothesis, concluded that the substance was an antimutagen and protective against mutagenicity and, possibly, carcinogenicity.
Some Chemicals Reported to be Both Mutagens and Antimutagens

As was noted by one author who examined both the mutagenicity and the antimutagenicity of a substance concluded that it was an antimutagen: In the presence of [the mutagen] … [the test substance] was found to be a very powerful antimutagen. At a concentration of only [], the number of [mutagen-induced] revertants was reduced considerably. … Interestingly, when [the test substance] was added without any [mutagen] being added, it caused an increase in mutation frequency.
And, to paraphrase another group of authors, Substance X enhanced the mutagenicity of the mutagen in TA98 indicating a co-mutagenic effect, and Substance Y showed a predominant antimutagenic activity in TA98. Previous studies showed that both Substance X and Substance Y are mutagenic in TA98 with S9. Experiments are being carried out to investigate the possible antigenotoxic effects of X and Y in vivo.
What may be the ultimate denial of evidence contrary to the author’s hypothesis is found in the following published conclusion. The author was justifying the recommendation for clinical, prophylactic use of a natural chemical, which was carcinogenic in rodents and reported to be both mutagenic and antimutagenic. Most chemicals which show carcinogenic activity only after long-term administration at a high dose [in the NTP studies] are non-mutagenic carcinogens. However, [this natural chemical that was carcinogenic at high doses in an NTP study] is positive in the Ames test. It seems unlikely that a mutagenic carcinogen would induce tumors only after long-term, high-dose administration. … Thus, the results of the NTP [carcinogenicity] study should not be generalized.
Ignoring, for the moment, the effect of toxicity on mutagenic and antimutagenic effects, the finding that some classes of substances can be both mutagenic and antimutagenic is not unexpected. Substances that participate in oxidizing-reducing reactions can scavenge DNA-reactive free radicals or produce such free radicals depending on the specific test protocol variations and the redox state of the reaction mixture.
It is also necessary to review the literature to determine whether the claimed antimutagen has been previously published to be a mutagen. An example of this from a Salmonella test can be seen in Table 5. In this study, the authors proclaimed the antimutagenicity of their test chemical in strain TA102 at low doses but were not aware of or did not cite a study published 11 years earlier or other published studies, showing the mutagenicity of the same substance in Salmonella strains TA98 and TA100 at similar or lower concentrations, or positive published studies of their chemical in other test systems.
Comparison of Mutagenic and Antimutagenic Concentrations

a The fold changes were calculated from the data taken from 2 published studies using the same test chemical. Only the responses judged mutagenic (strains TA98 and TA100), or antimutagenic (strain TA102) by their respective study authors are included in this table.
A final example from a published article illustrates some of the difficulties encountered when trying to identify a substance as an antimutagen. In this study (Table 6), a pure, nonmutagenic test chemical was tested for its potential antimutagenic effects on 3 known mutagens. As was noted by the authors, at equimolar concentrations the mutagenicity of 1 mutagen was completely inhibited (antimutagenic effect), the mutagenicity of another was significantly enhanced (synergistic mutagenesis), and there was no effect on the mutagenicity of the third. This study was presented by the authors as an example of the complexity involved when attempting to identify and classify chemicals as antimutagens.
Some of the above examples and conclusions appear to be naive when presented out of the context of their publications, but they represent the zeal that these authors have for identifying antimutagens, regardless of whether they are also mutagens (and, in some cases, carcinogens).
Effect of Adding a Test Antimutagen at Equimolar Concentrations to 3 Known Mutagens a

a Adapted from a published data table. The conclusions are those of the study’s authors.
Characterization of the Test Substance
The third point addressed is an issue that is not specific to mutagenicity and antimutagenicity studies, the chemical characterization of the plant extracts being tested. It is well known that although the chemical constituents of different samples of the same genus, species, and variety of plant may be the same, their relative proportions can differ dramatically depending on where and when in the year they were grown; farming practices, including the use of fertilizers and herbicides; the chemistry of the soil; the climate; the maturity and condition of the plant at the time it was harvested; and the harvest and storage conditions prior to its use in the laboratory. Because these various factors will affect the relative proportions of biologically active and inactive constituents, variations in their relative biological activities will also be affected. As a result, the relative proportions of the major constituents, especially those that are being implicated in the antimutagenic response, need to be characterized. In the absence of such characterization, it will be difficult for others to reproduce the reported effects or to attribute the measured effect to any specific chemical component or class of chemicals.
Conclusions
The antimutagenicity literature over the past 20 years is a mixture of objective experimentation and reporting, as well as reflections of researcher’s hopes and aspirations to find a cure for induced mutation and cancer. Mechanistic studies, rather than a simple counting of revertant colonies or chromosome aberrations, will provide the best demonstration of antimutagenicity that could be translated to potential human use. However, in the absence of such studies, the field of antimutagen identification could be greatly improved and enhanced by addressing the following issues when designing, evaluating, and reporting experiments.
What is the toxicity of the antimutagen, the reference mutagen, and the combination, and can the apparent antimutagenic response be explained or confounded by that toxicity? Meaningful antimutagenesis studies need to include measures of cytotoxicity in order to preclude toxicity as the basis for any reduction in mutant numbers. Such studies should include a series of concentrations of the test substance. Do the experiments also show mutagenic activity of the test substance, or has it been reported to be mutagenic by others in the same or other test systems? If so, the basis for ignoring or downplaying the mutagenic response when interpreting the antimutagenicity data must be specifically addressed. If plant or other extracts are being used, have they been characterized as to their principal chemical constituents? If a single chemical or chemical class is being singled out as the antimutagen, have the levels of those chemicals in the specific extracts used in the presented experiments been assessed?
The identification and/or characterization of antimutagenic (and, hopefully, anticarcinogenic) chemicals for dietary or medicinal use are worthy endeavors. Zeal is commendable, and an important component of scientific research, but experiments must be appropriately designed and conducted, and conclusions based on cautious interpretation of the data rather than on the hopes of the researcher or test sponsor.
Footnotes
Acknowledgment
I thank Drs David DeMarini (EPA) and Michael Shelby (NIEHS) for their insightful comments and suggestions.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.