
Abstract
Our objective was to study the toxicokinetics of aflatoxin (AF) in pregnant mice. Aflatoxin B1 (AFB1) was administered intraperitoneally (IP) to groups of pregnant mice in single doses of 20 mg/kg on gestation day (GD) 13 and orally at the same gestational age. Controls received (IP and oral) a proportionate volume of solvent only. Maternal blood was collected at 15, 30, 45, 60, 90, 120, and 150 minutes posttreatment. Their AFB1 contents were determined. Aflatoxin B1 concentrations following maternal exposure to AFB1 were highly correlated with time after exposure. The serum concentrations were predictable and the highest serum levels were seen immediately at 15 minutes in mice given AFs IP and at 30 minutes in those given it orally. The absorption was 5.0 μg/min and elimination was 3.0 μg/min. The toxicokinetics of AFB1 have been delineated. Aflatoxins are easily and rapidly absorbed both from the gastrointestinal tract (GI) tract and through the peritoneum.
Introduction
Aflatoxins (AF) are metabolites of fungi of genus Aspergillus flavus, Aspergillus parasiticus, and Aspergillus nomius. Aflatoxin B1 (AFB1) is the most potent hepatocarcinogen 1 known and is included in category 1A by International Agency for Research on Cancer (IARC). 2 Aflatoxin M1 (AFM1), the main hydroxylated AFB1 metabolite, is also a probable human carcinogen and is classified in the group 2B of the IARC. 3,4 These toxins are hepatotoxic, mutagenic, carcinogenic, and immunosuppressive. 5,6 Human exposure to AF occurs via contaminated food, 7 cow milk, and by air particularly in farm workers. 8,9 Aspergillus produces as many as 4 toxins abbreviated AFB1, B2, G1, and G2, respectively. Aflatoxin B1 is converted to its reactive epoxide by the action of cytochrome P450-dependent mixed function mono-oxygenase enzyme systems in the liver. 10
Hepatotoxic, mutagenic, and carcinogenic effects of AFB1 have been studied extensively. The AFB1 4-hydroxy derivative, AFM1, is secreted in milk 11 and deposited in tissues. 12 Fetuses suffer the deleterious effects of AFs by maternal consumption of contaminated food during pregnancy. 13,14 Studies on possible reproductive toxic effects appear to be limited to laboratory animals with variable results. We have established a mouse model in which AFB1 is injected on gestation day (GD) 13, crosses the placental barrier and induces intrauterine growth restriction (IUGR). The objectives of the current study were to study the toxicokinetics of AF in this model.
Materials and Methods
Aflatoxin B1 (from A flavus, molecular weight [MW] 312.27), AFM1 (from A flavus, MW 328.27) and the remaining reagents were purchased from Sigma-Aldrich Chime GmbH (Steinheim, Germany). For animal treatment, lyophilized powder of AFB1 was dissolved in carboxymethylcellulose (CMC), dimethyl sulfoxide (DMSO), or corn oil:ethanol mixture at 5.0 mg/mL and used immediately. Dimethyl sulfoxide (5 mg AFB1/1 mL DMSO) was found to dissolve the toxin best and therefore further experiments reported here used DMSO as the solvent. For high-performance liquid chromatography (HPLC) calibration, lyophilized powders of AFB1 (10 mg) and AFM1 (0.2 mg) were dissolved in 4.0 mL benzene:acetonitrile (9:1) and stored at −20°C; subsequent dilutions were made in methanol.
A single dose (IP) of 20 mg/kg body weight of AFB1 was administered in 140 to 180 μL on GD 13 to pregnant mice. The controls received a proportionate volume of vehicle (IP). Animals were sacrificed by cervical dislocation at 0, 15, 30, 45, 60, 90, 120, and 150 minutes, and blood samples were collected in coated vials quickly from the inferior vena cava. At least 4 control and experimental animals were used for each time point. The same dose of AFB1 was administered orally via an orogastric tube at GD13 to pregnant mice that were sacrificed and their blood collected at the same time points. Controls received the vehicle through the same route. Their AFB1 contents were determined according to the methods of Nelson et al 15 and Blaney et al. 16 Serum was extracted using hexane and chloroform as described in the following sentences: 1.0 mL hexane was added to 0.5 mL serum and the sample was vigorously mixed. The bottom layer was collected by centrifugation at 2000 g at 25°C, for 5 minutes. One milliliter of chloroform was then added, and the sample was mixed and centrifuged as above. The bottom layer was carefully collected and the procedure was repeated twice. The final chloroform layer was dried under nitrogen and stored at −20°C for less than 3 days. The dried pellets were reconstituted in methanol:water (50:50) immediately prior to HPLC analysis.
High-performance liquid chromatography analysis was performed on a Waters (Milford, Massachusetts) system gradient liquid chromatograph (model 515) with autoinjector (model 717). Aflatoxin B1 and its primary metabolite, AFM1, were detected using a Waters multi-fluorescence detector (model 2475) equipped with a standard flow cell and operated with 365 nm excitation and 480 nm emission filters. The 4.6 mm × 25 cm Beckman Octadecylsilane (5 μm) column was operated at 25°C at 2.0 mL/min. The solvent was acetonitrile:water (25:75). The chromatograph procedure used isocratic gradient linear calibration curves, from 4 to 40 ng (13-128 pmol) for AFB1 (r 2 > .998) and 20 to 48 ng (61-146 pmol) for AFM1 (r 2 > .996), with each set of analytical runs. Final toxin concentrations were calculated using the measured volume of serum.
Limit of detection and sensitivity of the method of extraction of AFB1 were calculated using the method described by Lamplugh 17 and the results were previously published. 10 Pooled sera from the control animals were first tested for the presence of AF and then spiked by transferring known amounts of AF standards (8, 5, 2.5 ng/mL) to different 10 mL tubes and evaporated to dryness under a stream of nitrogen. Four milliliters of serum were added to each tube and incubated at 30°C for 1 hour with gentle swirling. The tubes were then stored at 4°C until analysis.
Simple means (mean) and standard deviations (SDs) were calculated for the toxin at each time period. Significant difference of means was calculated using Mann-Whitney U test. Nonparametric correlation coefficients were calculated using both Kendall and Spearman test. Time evolution was modeled by regression of AFB1 and AFM1 over time. We used a fourth-order polynomial without intercept, integrated from 0 to 150 minutes for the oral route (where serum levels do not start at a maximum), and a box-cox regression (that is-use a power transform of the concentration) for the IP route (where maximum dose is reached immediately), also integrated from 0 to 150. For AFB1, the rate of absorption and elimination was calculated by fitting a double exponential to the data point. The coefficients of the 2 were the 2 parameters.
The Research Ethics Committee of the Faculty of Medicine and Health Sciences, UAE University, approved the study.
Result
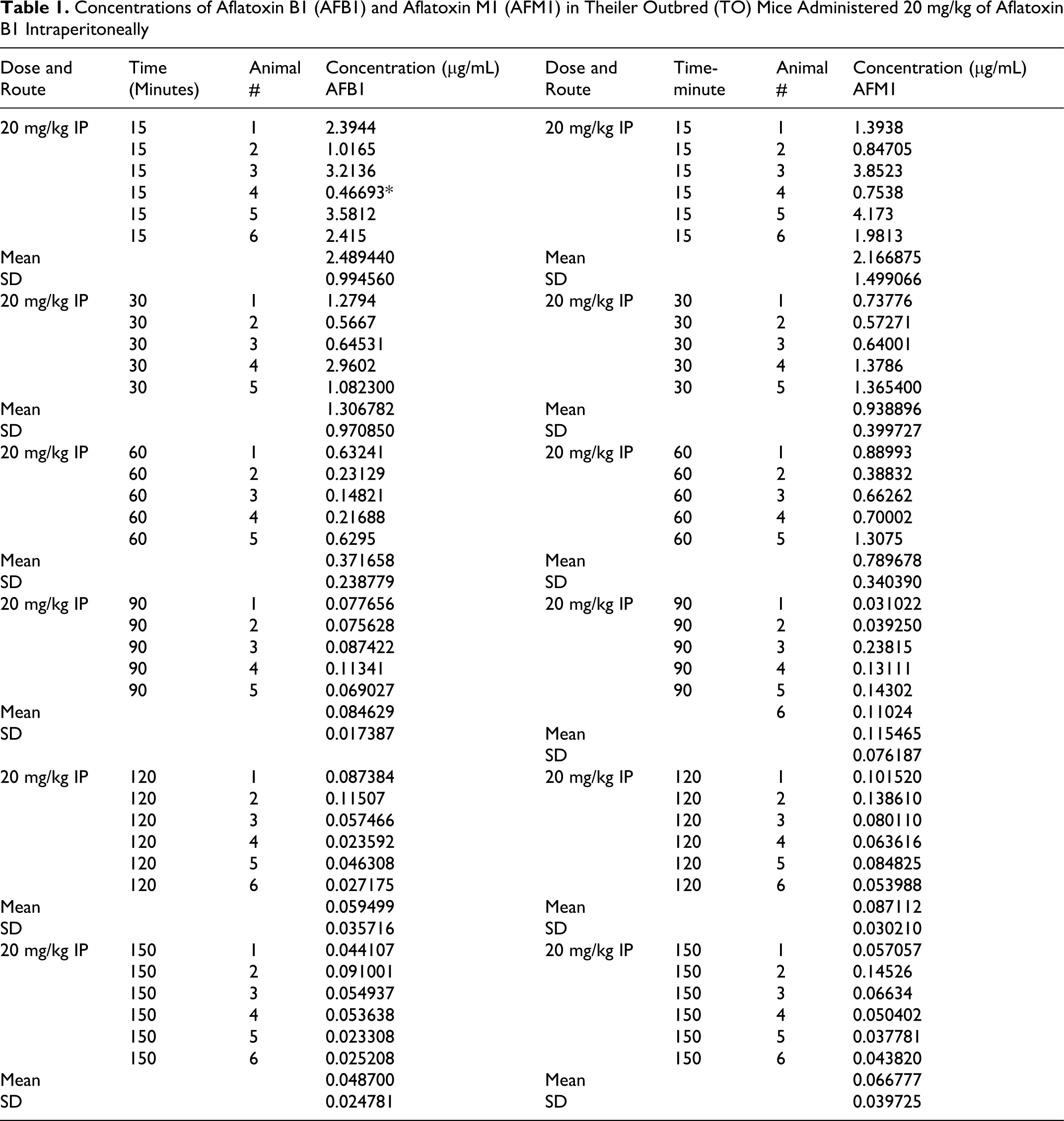
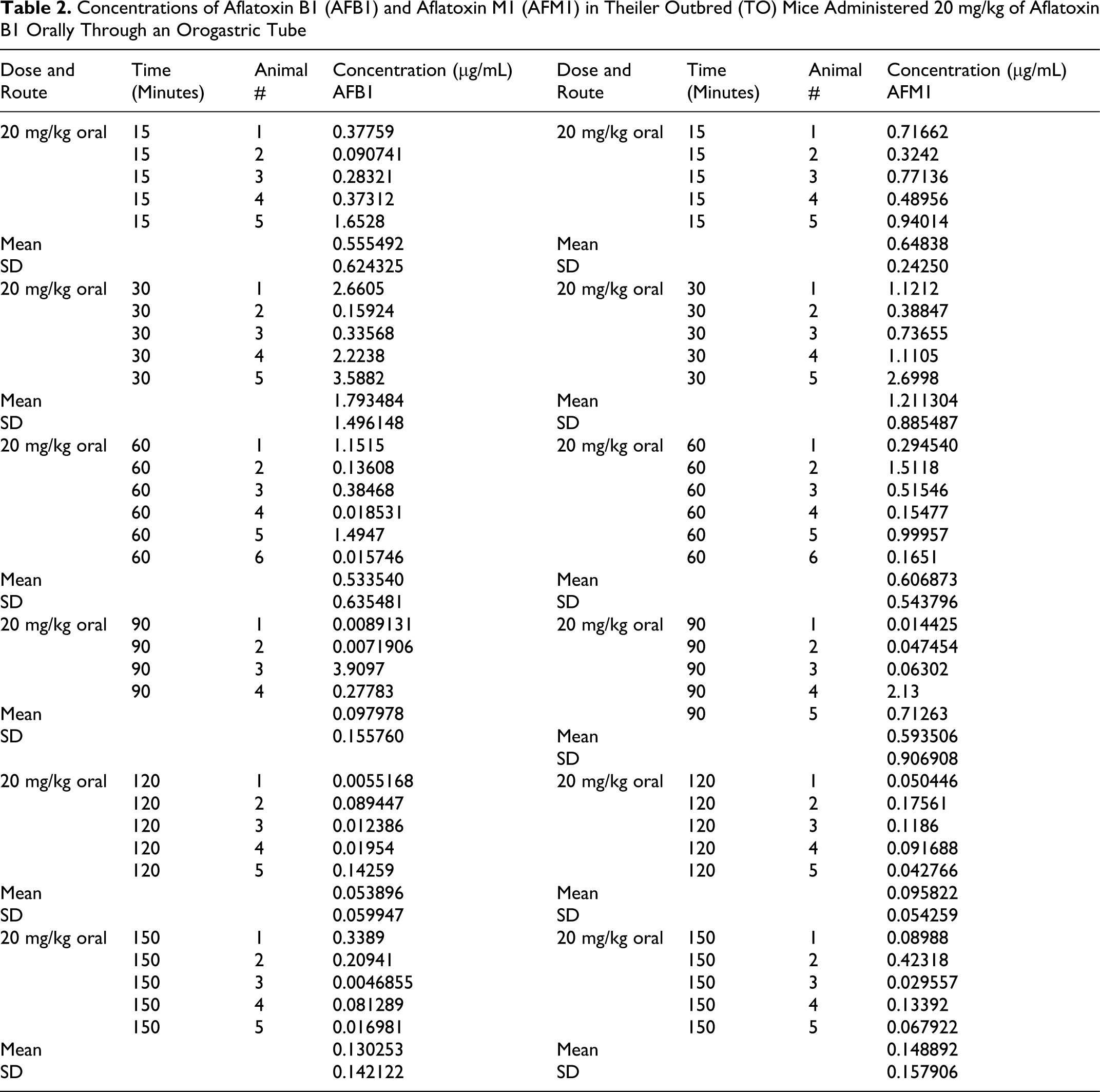
The AF levels in mice, treated with AF either suspended in corn oil or CMC, were highly unpredictable as the AFs did not form a homogeneous solution and therefore the absorption was erratic. We then used the minimal amount of DMSO (5 mg AFB1/mL DMSO) that would result in a homogenous solution with AFs. The serum concentrations were predictable and the highest serum levels were seen at 15 minutes in mice given AFs IP and slightly later at 30 minutes in those treated with AFB1 orally (Tables 1 and 2). The attached graphs illustrate the changes in concentrations over time. Both AFB1 and AFM1 followed a similar course with a similar fall in concentrations with time except at 60 and 90 minutes when the decrease in AFM1 concentration was slower (Figure 1). The correlation between AFB1 and AFM1 on one hand and time on the other was very significant (r = .99 and r = .96, respectively). The serum half-life of AFB1 was 16 minutes in the first 90 minutes and 77 minutes from 90 to 150 minutes. The serum half-life of AFM1 was 27 minutes over the entire period of observation (Figures 2 to 5). The area under the curve (AUC) for AFB1 administered IP was 62.55 μg·h/mL and that administered orally was 68.29 μg·h/mL. The AUC for AFM1 was 87.34 and 87.56 μg·h/mL in animals administered AFB1 IP and orally, respectively. Using nonlinear regression of log concentration of AFB1 (IP route) on time, we obtained a coefficient of 3.0 μg/min + 0.4 (SE), which is the elimination constant of AFB1 given through the IP route. Using parameter estimates to fit a double exponential to the oral administration and by taking logarithms of both sides, we reached the same parameter estimate of 5.0 μg/min + 2.7 (SE), which is the estimated absorption rate of AFB1 from the gut. The elimination constant for AFM1 was best estimated to be about 3.0 μg/min + 0.4 (SE).
Concentrations of Aflatoxin B1 (AFB1) and Aflatoxin M1 (AFM1) in Theiler Outbred (TO) Mice Administered 20 mg/kg of Aflatoxin B1 Intraperitoneally
Concentrations of Aflatoxin B1 (AFB1) and Aflatoxin M1 (AFM1) in Theiler Outbred (TO) Mice Administered 20 mg/kg of Aflatoxin B1 Orally Through an Orogastric Tube
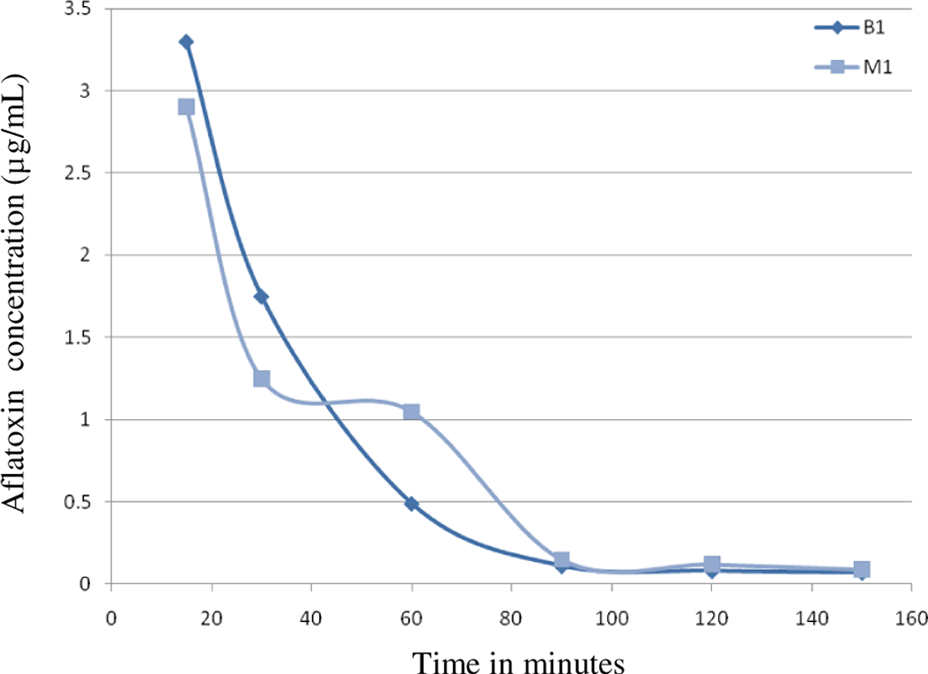
Aflatoxin (AF) B1 and M1 levels in the serum of pregnant mice injected intraperitoneally with 20 mg/kg body weight of aflatoxin B1 on day 13 of gestation. Serum levels show a rapid decline in AFB1 levels up to 90 minutes and a steep decline in AFM1 levels up to 60 minutes followed by a slower decline up to 90 minutes.
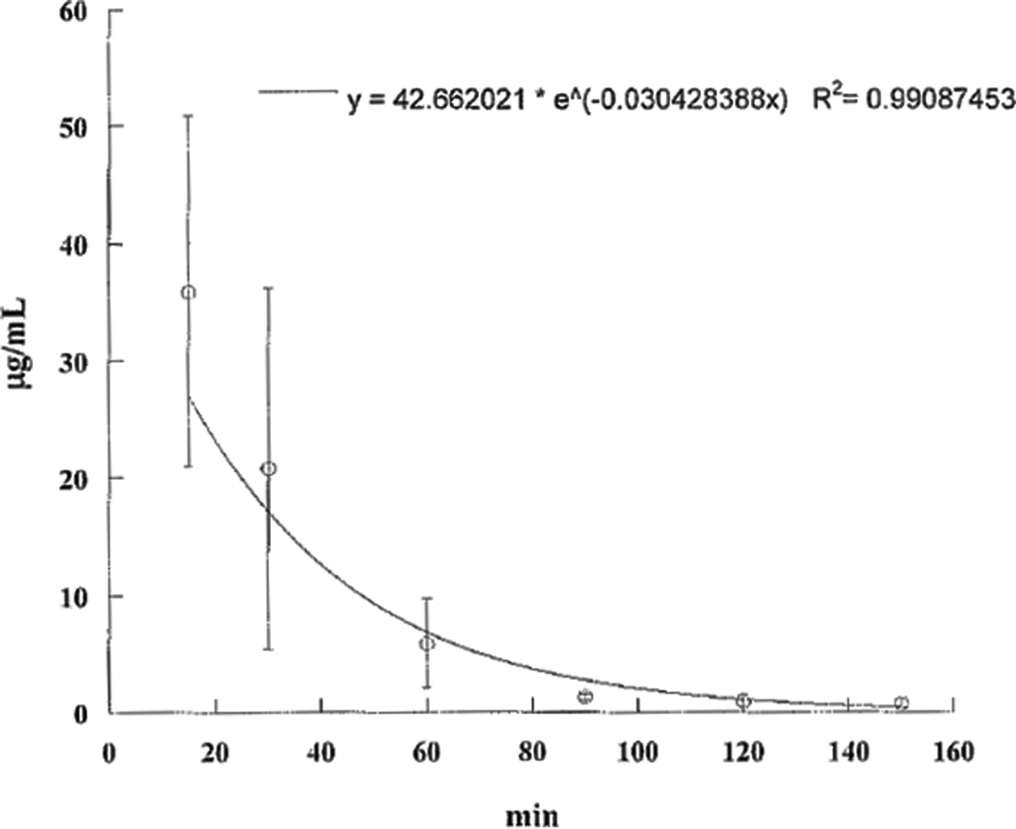
Aflatoxin (AF) B1 levels in the serum of pregnant mice injected intraperitoneally with 20 mg/kg body weight of aflatoxin B1 on day 13 of gestation. Aflatoxin B1 levels decline rapidly over the next 90 minutes.
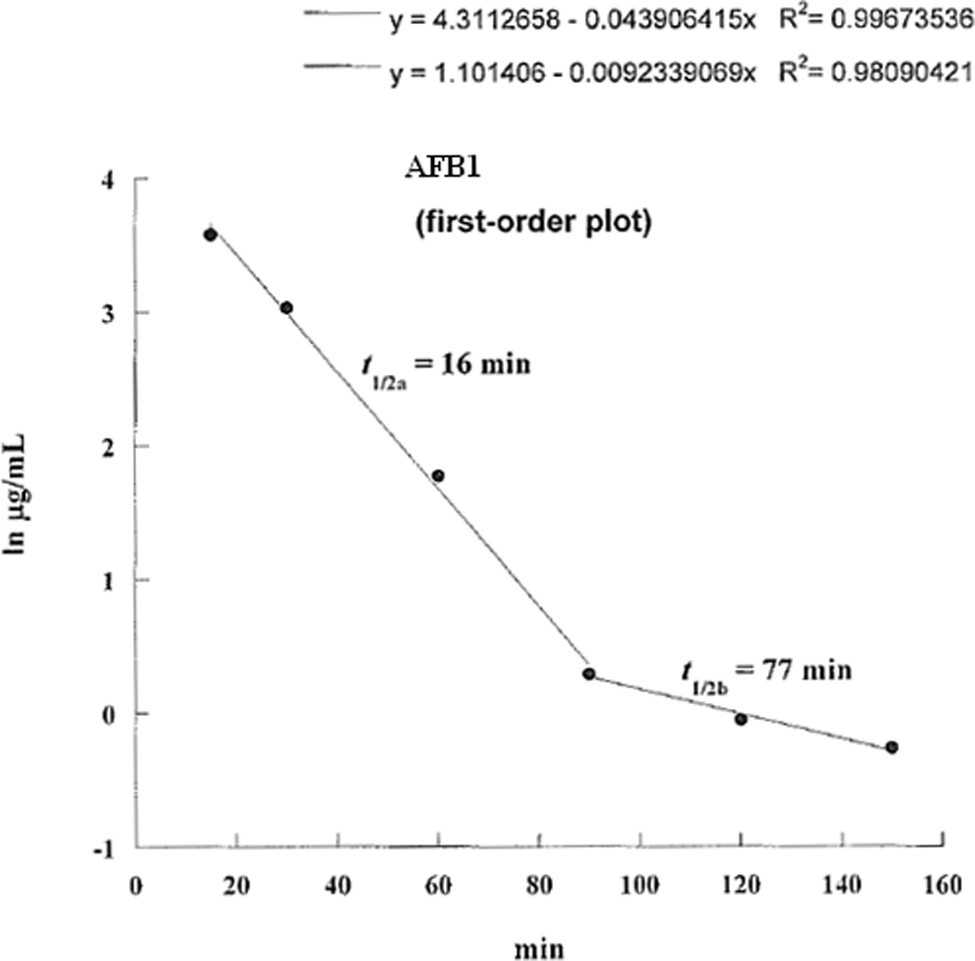
Log aflatoxin B1 levels in the serum of pregnant mice injected intraperitoneally with 20 mg/kg body weight of aflatoxin B1 on day 13 of gestation plotted against time. The half-life in the first 90 minutes is 16 minutes and from 90 to 150 minutes is 77 minutes.
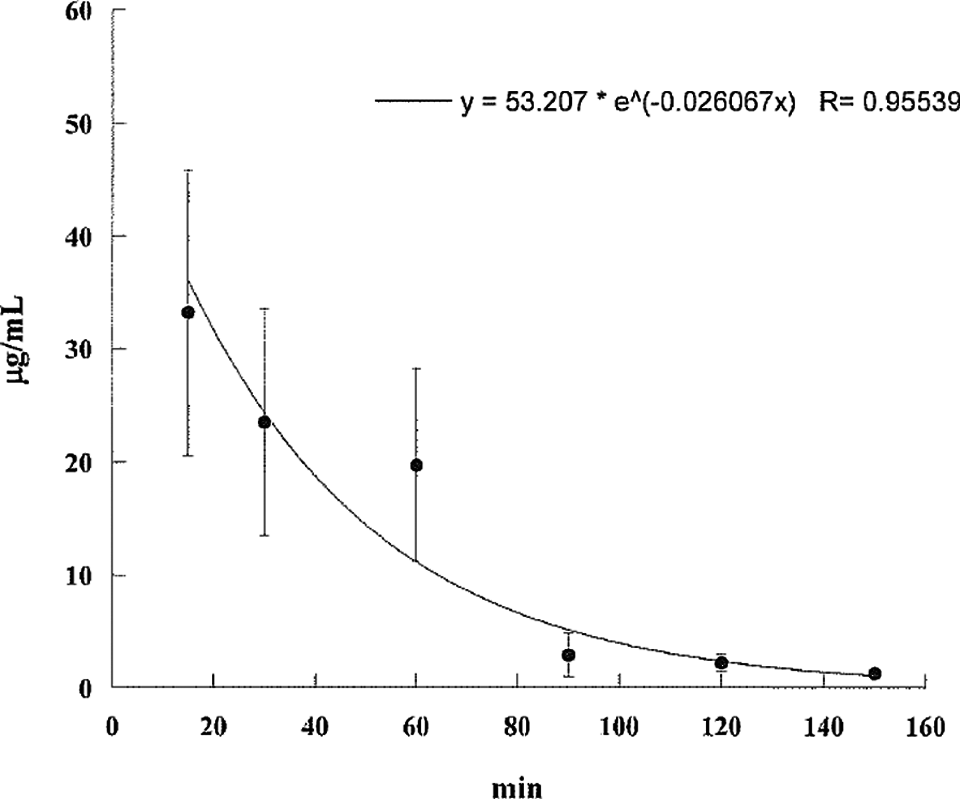
Aflatoxin M1 levels in the serum of pregnant mice injected intraperitoneally with 20 mg/kg body weight of aflatoxin B1 on day 13 of gestation. Aflatoxin M1 levels decline rapidly over the next 150 minutes.
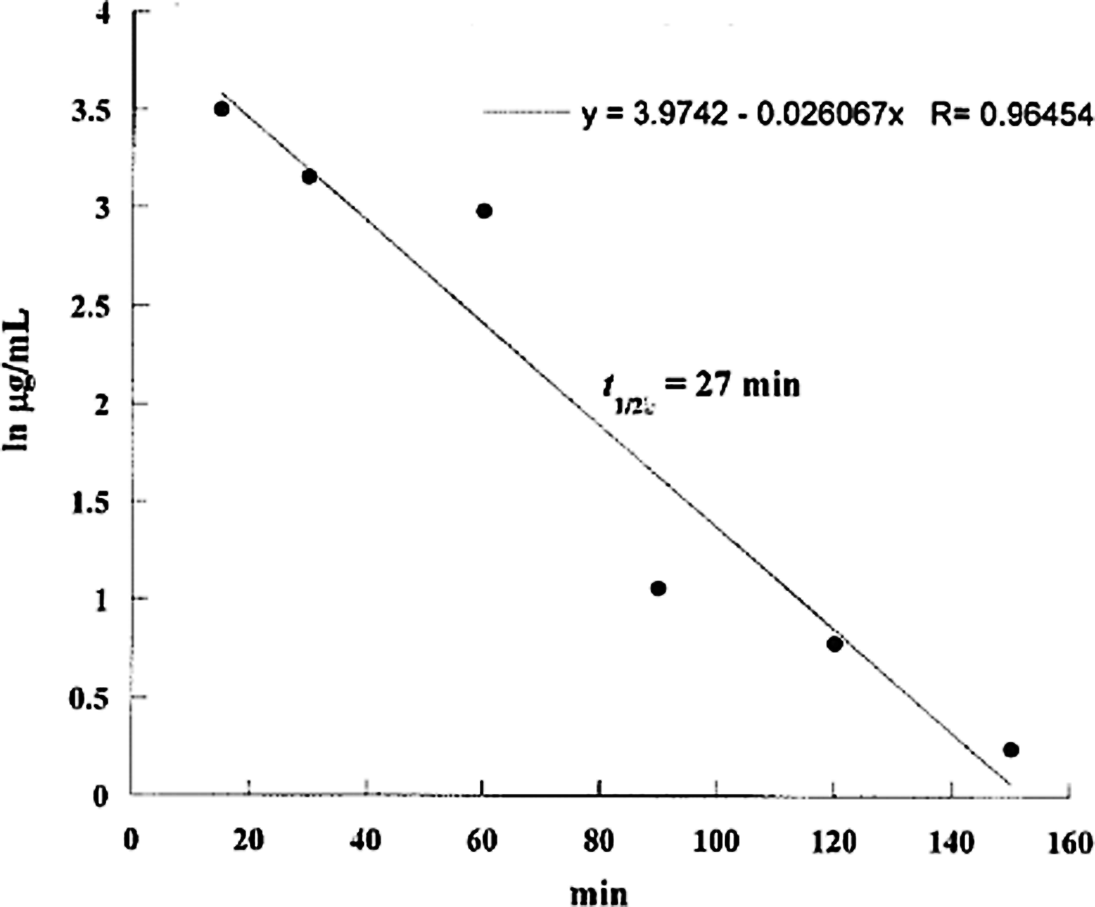
Log aflatoxin M1 levels in the serum of pregnant mice injected intraperitoneally with 20 mg/kg body weight of aflatoxin B1 on day 13 of gestation plotted against time. The half-life is calculated to be 27 minutes.
Discussion
Aflatoxin absorption from the gastrointestinal tract (GI) is rapid and excretion is via bile into the gut and via the urinary tract. There are differences in susceptibility across species 9,18 and between persons depending largely on how the toxin is metabolized into its epoxide and the reaction of the metabolites with protein and DNA. The toxin appears in the milk 8 and therefore it is important to detect as it could adversely affect the health of the infants. Most of these toxins are chemically stable so they tend to survive storage and processing and even cooking at high temperatures. The route of administration affects the result in that a smaller percentage of the dose is absorbed when given orally as compared with the toxin administered intraperitoneally (IP) or through the respiratory route, although in a study in goats and rats, 97% of AFB1 ingested was absorbed. 19 Nevertheless, the absorption is rapid and a significant level (70% that of IP route) is achieved 30 minutes after the dose is administered. In experiments using [14C]—AFB1 orally in lactating rats, about 65% was excreted through the feces, 8.8% in the urine, and 2.2% in milk. It was similar in goats except for a larger percentage of AF in urine. Liver contained 7.3% after intravenous (IV) dose and 4.9% 120 hours after oral dose in goats. 20 Aflatoxin B1, after absorption, associates with protein and goes into the liver where through the action of cytochrome P450 enzymes system, in particular CYP1A2, and CYP3A4, is oxidized to its epoxides. CYP1A2 readily oxidizes AFB1 to AFM1, AFD, and endo-8-9-epoxides. CYP3A4 oxidizes AFB1 to AFM1 and AFQ. In the current study, because AFM1 appears rapidly in the blood, it shows that the metabolism of AFB1 occurs immediately in the liver. Both AFB1 and AFM1 cross the placenta in large quantities. 21
The absorption rate of AFB1 from the gut was calculated to be 5.0 μg/min. We then fitted a double exponential to AFM1 (IP) values, and the second exponential should have been the elimination. This did not yield reliable estimators of the coefficients. Levels of AFM1 should have started at 0 (because it is formed in the liver from AFB1) and then have peaked and then be eliminated. The first observation however is too late to observe the peak. Apparently, formation of AFM1 from AFB1 is very rapid. It seems that the ratio AFB1/AFM1 is about constant, which is shown when we plot the AFB1 concentration on the X axis and the AFM1 concentration on the Y axis. Aflatoxin M1 is eliminated very rapidly, so that levels of AFM1 reflect recently metabolized AFB1, proportional to serum AFB1 levels. There are few data reported in the literature regarding metabolism and toxicity of AFM1 in humans as it is considered a detoxification product of AFB1. However, there is 1 study that reveals a higher toxic activity of AFM1 than AFB1 reported in 2 human cell lines, also in the absence of metabolic activation. 19 In the current work, we have evaluated the absorption of AFB1 and its metabolism into AFM1. Our study has also shown that AFM1 forms immediately after the absorption of AFB1 both from the IP and oral routes, and follows AFB1 closely in its elimination from the body in a short time with an elimination rate of 3.0 μg/min.
The current study is the first study to present absorption and elimination data using pregnant animals, all previous studies were on either nonpregnant or lactating animals. In humans, 1 study revealed rapid human AFB1 uptake (plasma k(a), 5.05 + 1.10 hours (−1); T(max), 1.0 hour) and urinary elimination (95% complete by 24 hours) kinetics. 22 In rats, a study about intestinal absorption kinetics, to evaluate the rate at which the 4 main AFs (AFs B1, B2, G1, and G2) are able to cross the luminal membrane of the rat small intestine, showed that the absorption of AFs in rat small intestine is a very rapid process that follows first-order kinetics, with absorption rate constants (ka) of 5.84 + 0.05 (AFB1), 4.06 + 0.09 (AFB2), 2.09 + 0.03 (AFG1), and 1.58 + 0.04 (AFG2) hours (−1), respectively. 23 The AFB1 absorption rate found in this study in rats is very similar to the one in the current study in mice. When a comparison is made between the amount of AFBI in rat and mouse stomachs after a single dose, it is clear that the mouse absorbs the toxin more quickly than the rat. The mouse absorbs 75% of ingested AFB1 after an hour while the rat absorbs 60% of the ingested dose in the same period. 24 Absorption is also rapid and follows first-order kinetics when AFB1 is administered intratracheally (IT) in rats. 25 A study of toxicokinetics of IT and orally administered [3H]AFB1 in male Sprague-Dawley rats has found the time-to-peak for the intratracheally treated group was less than that for the oral group, but the disappearance of label from the blood followed a nearly identical course in both groups. The plasma half-lives (t1/2) were 87.7 and 91.8 hours for the IT and oral groups, respectively, 26 and were many times higher than the half-life of the AFB1 in mice from our study (16 minutes in the first 90 minutes and 77 minutes in the period between 90 and 150 minutes). In lactating Holstein dairy cows given a single oral bolus of a naturally contaminated corn meal, AFB1 in plasma peaked (33.6 ng/L) as soon as 20 minutes after treatment. The plasma AFM1 was already detectable at 5 minutes (10.4 ng/L) and peaked at 25 minutes (136.3 ng/L). 27 This is very similar to the findings in the current study.
Excretion of AFB1 and its metabolites occurs primarily with bile and to a lesser extent with urine. Elimination of AFB1 appears slow in all species and strains studied. Hiesh and Wong 28 reported that the total excretion of 14C-labelled AFB1 was 80%, 72%, and 73% of an IV administered dose in male mice, rats, and monkeys, respectively, within 100 hours after IV dosing. The excretion was most intensive during the first 24 hours after dosing. However, it was reported that approximately 80% of a single IP dose of 14C-labelled AFB1 in rats was excreted within 24 hours postinjection. In the current study, the elimination constant was found to be 3.0 μg/min for both AFB1 and AFM1.
The data reported here are of particular significance in view of the potential adverse impact on growth and development of the embryo.
Conclusions
The toxicokinetics of AFB1 in pregnant mice have been delineated and show large amounts being easily and rapidly absorbed both from the GI tract and through the peritoneum.
Footnotes
The author(s) declared no conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: Research Affairs at the UAE University under a contract no. 17-7-11/02.