
Abstract
G protein–coupled receptors (GPCRs) exist as collections of conformations in equilibrium, and the efficacy of drugs has been proposed to be associated with their absolute and relative affinities for these different conformations. The serotonin 2A (5-HT2A) receptor regulates multiple physiological functions, is involved in the pathophysiology of schizophrenia, and serves as an important target of atypical antipsychotic drugs. This receptor was one of the first GPCRs for which the functional selectivity phenomenon was observed, with its various ligands exerting differential effects on the phospholipase A2 (PLA2) and phospholipase C (PLC) signaling pathways. We aimed to develop a multiplex functional assay in 96-well plates for the simultaneous measurement of the PLA2 and PLC pathways coupled to 5-HT2A receptors; this approach enables the detection of either functional selectivity or cooperativity phenomena in early drug screening stages. The suitability of the method for running screening campaigns was tested using the Prestwick Chemical Library, and 22 confirmed hits with activities of more than 90% were identified; 11 of these hits produced statistically significant differences between the two effector pathways. Thus, we have developed a miniaturized multiplex assay in 96-well plates to measure functional selectivity for 5-HT2A receptors in the early stages of the drug discovery process.
Introduction
5-HT2A receptors are G protein–coupled receptors (GPCRs) that regulate multiple physiological functions and are relevant targets of atypical antipsychotic drugs. 1 5-HT2A receptors were one of the first GPCRs for which the functional selectivity phenomenon was observed, with different ligands differentially activating arachidonic acid (AA) release and inositol phosphate (IP) formation for the same receptor.2,3 This receptor, upon binding to an agonist, activates phospholipase C (PLC) through Gαq, resulting in the formation of IP and diacylglycerol (DAG). 4 Furthermore, the receptors can also activate phospholipase A2 (PLA2), thus inducing AA release, although the trigger mechanism remains unclear. 5
We have previously demonstrated the existence of 5-HT2A homooligomers both in transfected cells and in the human brain. This oligomerization evidenced a negative cooperativity phenomenon that was observed for certain ligands, such as clozapine, through the PLA2 signaling pathway, while other ligands, such as haloperidol, demonstrated no cooperativity between protomers in this pathway. In contrast, no cooperativity was observed for the PLC-mediated pathway. 6
Phenomena such as cooperativity or functional selectivity result from an equilibrium between dynamic receptor conformations, and the efficacy of drugs has been proposed to be associated with their absolute and relative affinities for these different conformations. 7 The current challenge in receptor pharmacology is development and optimization of methods to detect conformational changes. 8
Our hypothesis in this work was that the development of a multiplex and miniaturized methodology that can simultaneously measure activation of the PLC and PLA2 signaling pathways by 5-HT2A receptors would allow the evaluation of functional selectivity and cooperativity phenomena in the first stages of drug discovery. Our aim in this work was to develop a miniaturized functional assay for the simultaneous measurement of the PLC and PLA2 signaling pathways coupled to the 5-HT2A receptor using the Prestwick Chemical Library for technology validation.
Materials and Methods
Drugs and Reagents
[3H]myo-Inositol (20.3 Ci/mmol), [3H]arachidonic acid (204.7 Ci/mmol), and [14C]arachidonic acid (57.1 mCi/mmol) were purchased from PerkinElmer Life Science (Waltham, MA). Clozapine and serotonin hydrochloride were purchased from Sigma-Aldrich (St. Louis, MO). AG 1-X8 resin was purchased from Bio-Rad (Hercules, CA). RNA binding YSi SPA beads and OptiPhase SuperMix scintillation cocktail were purchased from PerkinElmer. UniverSol-ES liquid scintillation cocktail was purchased from MP Biomedicals (Santa Ana, CA). Bovine serum albumin (BSA) fraction V, fatty acid-free, was purchased from Roche (Basel, Switzerland). MultiScreen filter plates were purchased from Millipore (Billerica, MA). All other reagent-grade materials were purchased from Sigma-Aldrich.
Cell Culture
Chinese hamster ovary cells stably expressing human 5-HT2A receptors at a density of ~200 fmol/mg protein (CHO-FA4 cells) were maintained in standard tissue culture plates (150 mm diameter) in Dulbecco’s Modified Eagle’s Medium-F12 (Gibco, Carlsbad, CA) supplemented with 10% (v/v) fetal bovine serum (FBS; Sigma-Aldrich), 1%
Measurement of arachidonic acid release and inositol phosphate accumulation
Measurements of AA release and IP accumulation were simultaneously performed using the same wells.2,9
Cells were seeded into 24- or 96-well tissue culture plates at a density of 5 × 104 or 2 × 104 cells/well, respectively. After 24 h, the culture medium was replaced by serum-free medium with 10 μCi/mL [3H]myo-inositol (20.3 Ci/mmol) for 24 h (if appropriate) and 1 μCi/mL [3H]arachidonic acid (202 Ci/mmol) or 0.2 μCi/mL [14C]arachidonic acid (57.1 mCi/mmol) for 4 h at 37 °C.
After the labeling period, the cells were washed at 37 °C with Hanks’ Balanced Salt Solution (HBSS; Sigma-Aldrich) supplemented with 20 mM HEPES, 20 mM LiCl, and different concentrations of fatty acid-free BSA (experimental medium), as described in the results section. After washing, the cells were incubated at 37 °C for 20 min with experimental medium containing either vehicle or the indicated concentrations of drugs. At the end of the incubation time, either 400 μL of the medium was added to counting tubes with UniverSol-ES liquid scintillation cocktail or 90 µL of the medium was added to a flexiplate with OptiPhase SuperMix cocktail (24- and 96-well plates, respectively) to detect either [3H] or [14C], respectively, which corresponds to AA release. The remaining medium was discarded, and 100 mM formic acid was added to the cells for 30 min at 4 °C. Aliquots of the medium were collected to measure IP accumulation. Two methods for this analysis were employed: AG 1-X8 resin or RNA binding YSi SPA beads. For the AG 1-X8 resin, after cell lysis, the medium was transferred to MultiScreen filter plates with AG 1-X8 resin. After filtering, the resin was washed with 100 μL of distilled water for 5 min and filtered again. Subsequently, 100 µL of elution buffer (1 M ammonium formate and 0.1 M formic acid) was added for 5 min and filtered over another plate. Fifty microliters of eluate was transferred to a flexiplate with 150 µL of OptiPhase to measure [3H]IPs. For the RNA binding YSi SPA beads, after incubation with formic acid, 20 μL of lysate was removed and transferred to a 96-well flexiplate with 80 μL of a 1:16 dilution of RNA binding YSi SPA beads to measure the accumulation of IPs.
Radioactivity was quantified with a WALLAC Microbeta TriLux 1450-023 liquid scintillation counter.
Screening for 5-HT2A Receptor Antagonists from the Prestwick Chemical Library by Measuring both the PLA2 and PLC Signaling Pathways
The Prestwick Chemical Library comprising 1120 compounds was used for screening.
Cells were seeded in 96-well culture plates at a density of 2 × 104 cells/well and were labeled with 10 μCi/mL [3H]myo-inositol in serum-free medium 24 h later. After 20 h, the medium was replaced by fresh medium containing 0.2 μCi/mL [14C]arachidonic acid. Four hours after the labeling period, cells were washed once and incubated for 10 min at 37 °C with experimental medium. The compounds were added using a JANUS automated workstation (PerkinElmer) at a final concentration of 10 µM. We used clozapine 10 μM as an inhibition control and serotonin 1 µM as a stimulus control. The compounds were incubated for 20 min, and then 90 µL of media was transferred using JANUS to a flexiplate with 150 μL of OptiPhase for the measurement of [14C], which corresponds to AA release. The remaining medium was discarded, and 200 μL of 100 mM formic acid was added to the cells for 30 min at 4 °C. Twenty microliters of the lysate was transferred with JANUS to a flexiplate and mixed with 80 μL of an RNA binding YSi SPA bead solution to measure the accumulation of [3H]IPs from the cells.
Radioactivity was quantified with a WALLAC Microbeta TriLux 1450-023 liquid scintillation counter.
Data Analysis
Nonlinear fitting of the concentration–response curves was carried out using Prism 4.0 software (GraphPad Software, La Jolla, CA) by applying both a four-parameter logistic equation and a two-site competition equation. Statistical comparisons between fits were performed using extra sum-of-squares F tests. 10 Statistical comparisons between different effector pathways were performed using Student t tests. Statistical significance was set at p < 0.05.
Results
5-HT-Induced AA Release in CHO-FA4 Cells Expressing 5-HT2A Receptors in 24-Well Plates
PLA2 activation was measured after the stimulation of 5-HT2A receptors stably expressed in CHO-FA4 cells grown in 24-well plates. The natural agonist serotonin (5-HT) promoted the release of AA in a concentration-dependent manner, exhibiting a potency (pEC50) of 7.02 ± 0.13 (
5-HT-Induced AA Release in CHO-FA4 Cells Expressing 5-HT2A Receptors in 96-Well Plates
The BSA concentration in the assay medium was first optimized. Cells were seeded at 20,000 cells/well 48 h before the assay. During the last 4 h, the cells were labeled and then washed as indicated in the protocol described above. BSA concentrations of 0.5%, 1.5%, and 2% were tested in the experimental medium. As shown in Figure 1 , the profile obtained for 5-HT was identical to that observed using 24-well plates, with 2% BSA providing the highest signal/background ratio ( Table 1 ).
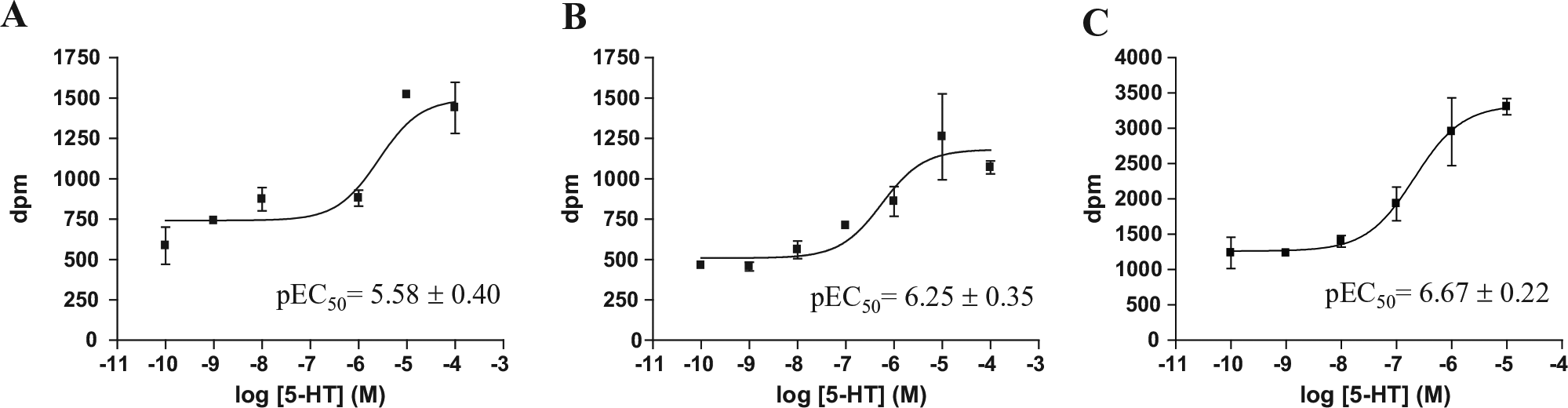
Concentration–response curves for serotonin-induced AA release in 96-well plate with different concentrations of BSA in the assay medium. (
Potency (pEC50) and Signal/Background Ratio of 5-HT-Induced AA Release in CHO-FA4 Cells Expressing Human 5-HT2A Receptors.

Values represent mean ± SEM of triplicate measurements.
Simultaneous Measurement of [14C]AA Release and [3H]IP Accumulation in CHO-FA4 Cells Expressing 5-HT2A Receptors in 96-Well Plates
Serotonin stimulated AA release and IP formation in a concentration-dependent manner, demonstrating a monophasic profile for signaling pathways, with pEC50 = 7.16 ± 0.17 (
Fig. 2A
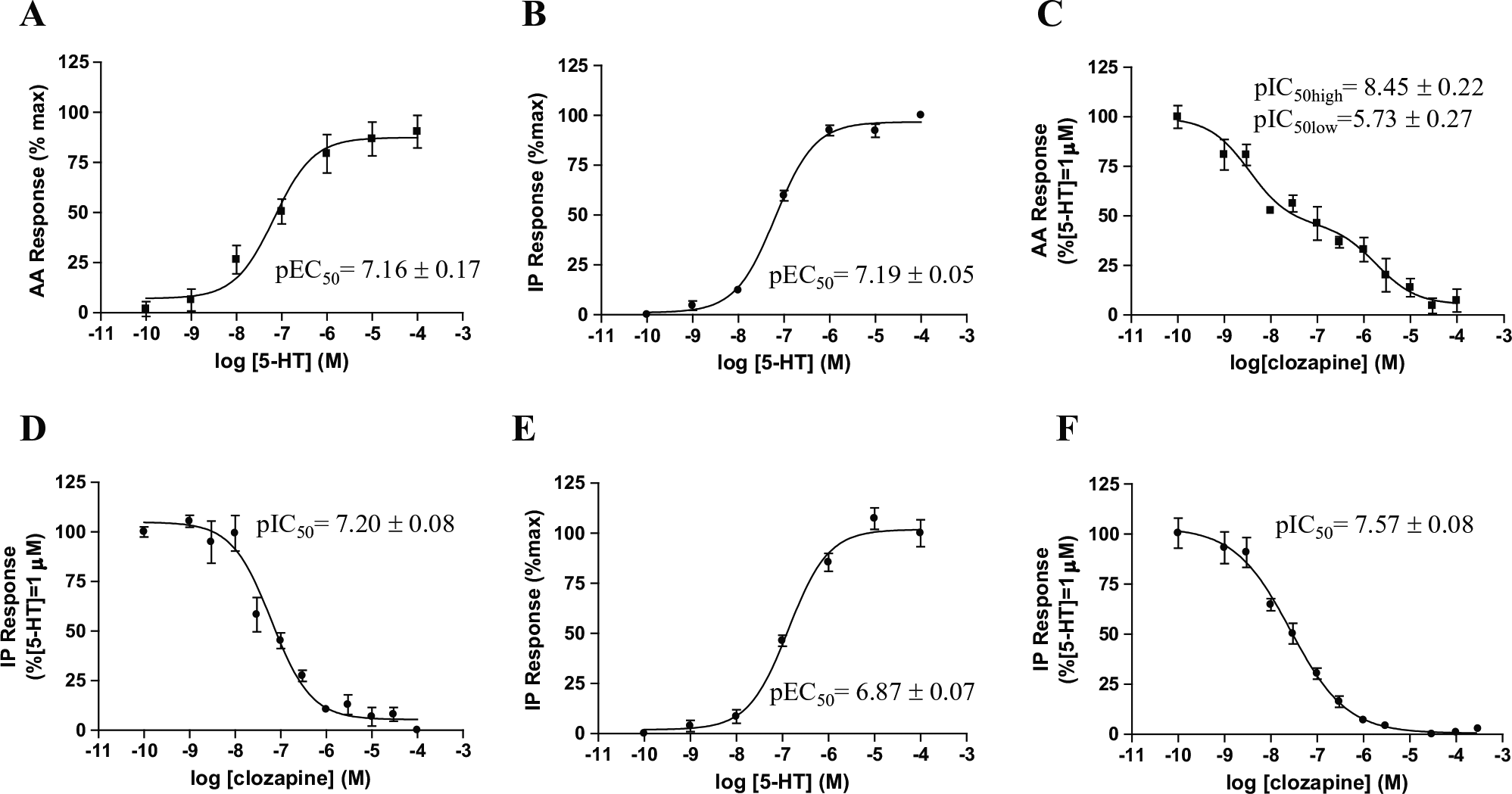
Concentration–response curves for 5-HT and clozapine. (
Potency of 5-HT (pEC50) and Clozapine (pIC50) Determined from the Experiments Illustrated in Figure 2.

[14C]AA release and [3H]IP accumulation (functional assays) in CHO-FA4 cells expressing human 5-HT2A receptors. [3H]IP accumulation was measured by using AG 1-X8 resin. Values represent the mean ± SEM of three independent experiments (n = 3) performed with triplicate measurements.
The antagonistic potency of clozapine was evaluated for the PLA2 and PLC pathways, and different inhibition profiles were observed for the two pathways. Clozapine inhibited 5-HT-induced AA release with a biphasic inhibition profile, whereas, for the PLC-mediated pathway, the compound generated a monophasic competition profile (
Fig. 2C
The use of ion exchange chromatography for measuring inositol phosphate formation is not an easily automatable process; therefore, we employed a previously reported approach to quantify the accumulation of IPs using dilutions of RNA binding YSi SPA beads.
11
The profiles obtained for both evaluated compounds were similar to those obtained by ion exchange chromatography, with pEC50 = 6.87 ± 0.07 for 5-HT and pIC50 = 7.57 ± 0.08 for clozapine (
Fig. 2E
Evaluation of Prestwick Chemical Library as Proof of Concept
Prestwick Chemical Library, which is composed of 1120 compounds, was employed to validate the method described above. The hit threshold was fixed at mean + 3 SD, and 38 hits with activities greater than 90% were identified. The Z′ average value was 0.7 ± 0.08 for IP formation and 0.5 ± 0.1 for AA release. Twenty-two hits were confirmed by concentration–response curve studies for both effector pathways. Eleven of these compounds generated significantly different effects on the two effector pathways (
Fig. 3
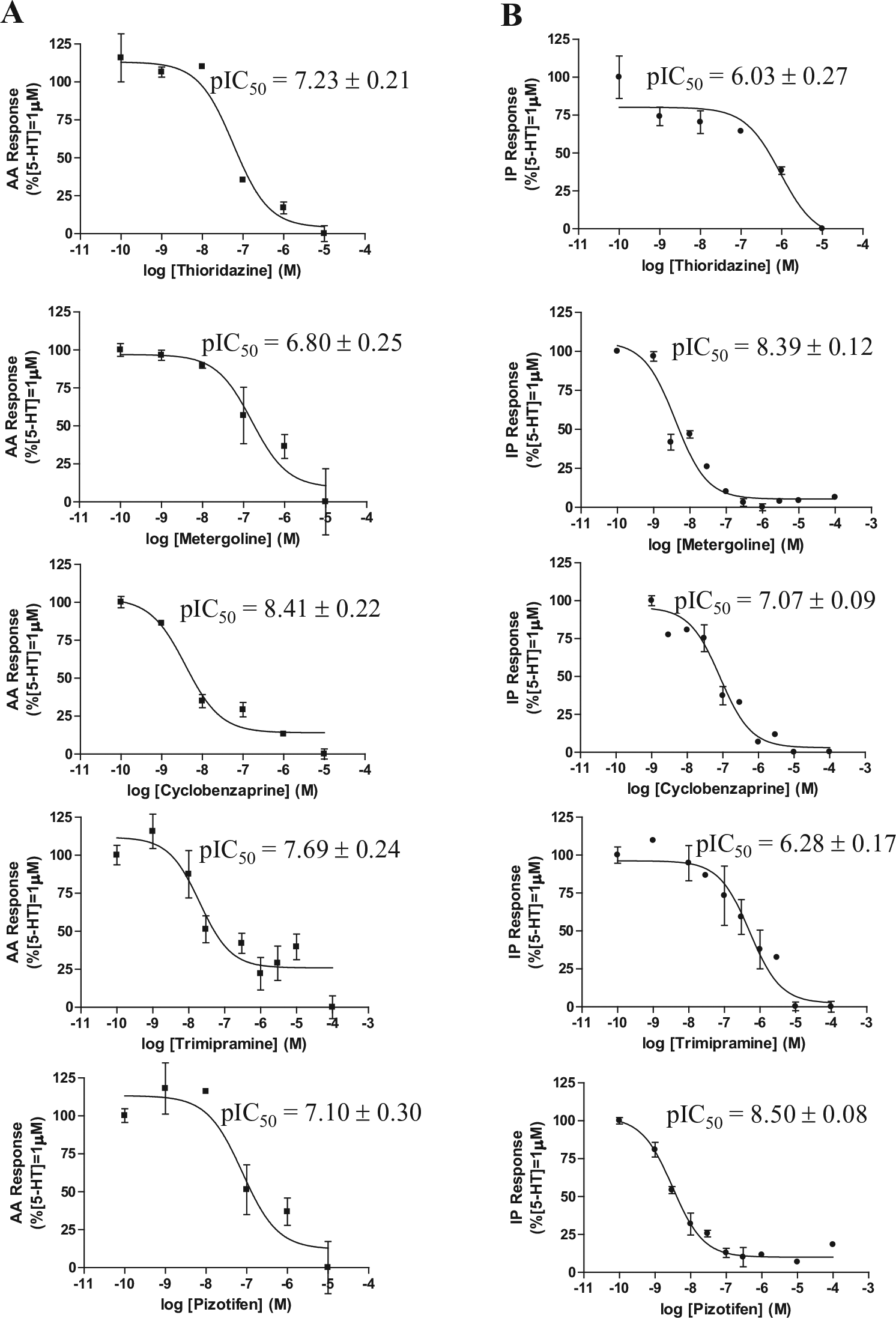
Concentration–response curves for different compounds demonstrating functional selectivity. (
pIC50 Values for Compounds That Demonstrated Activity in Both Signaling Pathways.
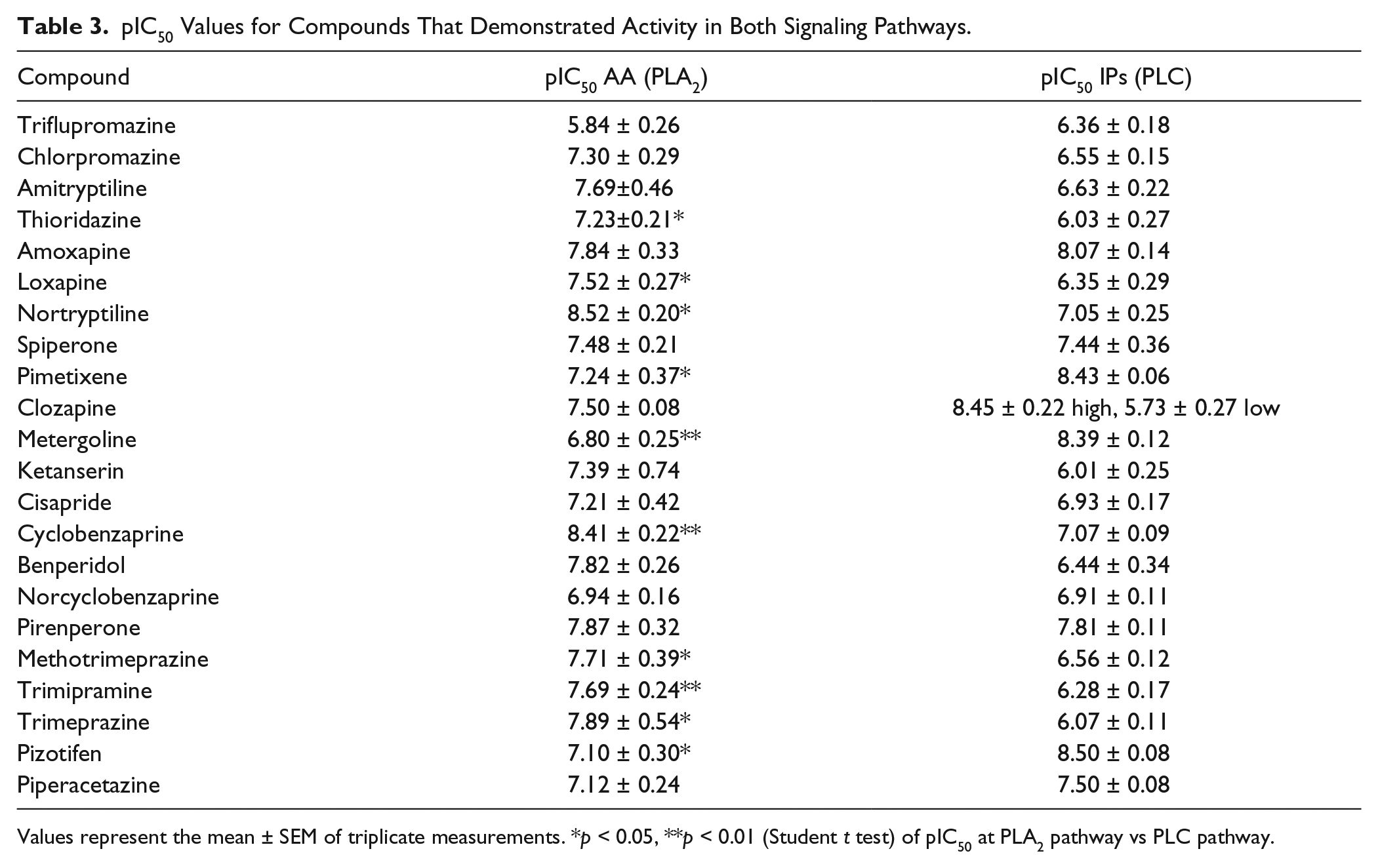
Values represent the mean ± SEM of triplicate measurements. *p < 0.05, **p < 0.01 (Student t test) of pIC50 at PLA2 pathway vs PLC pathway.
Discussion
The major finding of this work was the development of a multiplex assay in 96-well plates that allowed us to detect compounds exhibiting functional selectivity in an automated screening process. This method was suitable for high-throughput screening (HTS), as was evidenced by screening the Prestwick Chemical Library.
Previously, our group observed functional selectivity for human 5-HT2A receptors in cells seeded in 12-well plates. In that work, we proposed a model based on the existence of an equilibrium between the receptor in its inactive dimeric form (R2) and two distinct active ([R2]* and [R2]**) receptor states. This model was able to describe the differential functional antagonist profiles observed experimentally by assuming a different receptor active conformation for each pathway (e.g., [R2]* for IP accumulation and [R2]** for AA release). 6
The low throughput of this assay hindered the discovery of novel compounds demonstrating functional selectivity for the human 5-HT2A receptor, which is one of the current challenges in pharmacology. 7 Because of this hurdle, we aimed to develop a 96-well multiplex assay that could be employed for hit identification and also detect compounds exhibiting functional selectivity. As a first step, the [3H]AA release induced by 5-HT in 24-well plates was characterized finding potencies similar to those obtained using 12-well plates in previous studies conducted by our group (pEC50 = 6.60 ± 0.25). 6 Subsequently, the protocol to measuring [3H]AA release in 96-well plates was optimized. For this purpose, different concentrations of cells were tested, and 20,000 cells/well was established as the optimal compromise between cell number and signal (data not shown).
When the assay volume was reduced, a decrease in [3H]AA release was observed, resulting in a diminished assay window. The BSA concentration in the assay medium was optimized to increase the [3H]AA signal/background ratio. BSA interacts with fatty acids (FAs), forming a BSA-FA complex. Therefore, the protein interacts with AA in the cell membrane, inducing its release into the medium and allowing its detection. The percentage of BSA in the experimental medium was fixed at 2%, with a signal/background ratio of 15.16. An increase in basal AA levels was observed with 2% BSA, evidencing that the AA flow from cells to supernatant is favored with higher BSA concentrations.
Miniaturization to the 96-well format led to a reduction in radioactive counts for [3H]AA release, which could interfere with [3H]myo-inositol analysis. To avoid this potential interference, [3H]AA was replaced with [14C]AA that is detected in a different window to improve assay reproducibility.
When the assays were carried out in either 12- or 24-well formats, IPs were isolated by ion exchange chromatography following the method described by Berridge et al. 12 A similar approach was first employed by using AG 1-X8 resin in 96-well filtration plates. However, this experimental approach was not useful for automated screening. Thus, we utilized positively charged RNA binding YSi beads with scintillation liquid; the beads interact with the negative charges of IP groups and thus function as an ion exchange matrix. Both, ion exchange chromatography and scintillation proximity assay methods demonstrated the same functional profiles in terms of efficacy and potency for both 5-HT and clozapine.
Additional optimization was performed for the washing steps carried out to remove unincorporated [3H]myo-inositol and [14C]AA. In the 12-well assay, three washes of 5 min were employed, which were replaced by a single wash of 10 min in the 96-well plates. Thus, the risk of cell loss (and therefore the obtained signal) was minimized.
Using our methodology, clozapine was found to demonstrate functional selectivity, inhibiting 5-HT stimulation with a monophasic profile through the PLC pathway and a biphasic profile for the PLA2-mediated pathway. This biphasic profile was already observed in previous studies conducted by our group using both binding and functional assays in 12-well nonautomated measurements. Oligomeric GPCRs characteristically exhibit negative cooperativity between the binding sites of both protomers, in which the binding of a ligand to an orthosteric center hinders the binding of the second ligand to another orthosteric center of the dimer. 6
The novel automated methodology suitability for running screening campaigns was demonstrated by screening the Prestwick Chemical Library. This chemical library was selected as a proof-of-concept screening as we aimed to ascertain if we were able to detect compounds with known activity at these receptors and their functional selectivity profile. Assay showed to be robust at both signaling pathways, with an average Z′ value of 0.7 ± 0.08 for IP formation and 0.5 ± 0.1 for AA release, and 38 hits with greater than 90% activity (mean + 3 SD) detected in both signaling pathways. There were several ligands with activities of less than 90% that would behave as 5-HT2A receptor antagonists; however, we fixed the common accepted criterion for hit threshold in order to filter only a limited number of compounds, showing the higher activities of the whole chemical library for further studies. Most of the identified 5-HT2A antagonists were antipsychotics or antidepressants with known activity at this receptor, thus validating the methodology’s sensitivity. Moreover, similar to clozapine, certain compounds demonstrated functional selectivity between the signaling pathways studied. From the 22 hits that were confirmed by the construction of concentration–response curves for both signaling pathways, 10 compounds produced statistically significant differences between PLA2 and PLC signaling pathways ( Table 3 ). Thioridazine, loxapine, methotrimeprazine (all antipsychotics); nortryptiline, cyclobenzaprine, and trimipramine (antidepressants); and trimeprazine (antihistamine) demonstrated greater potency in inhibiting the PLA2-mediated signaling pathway than the PLC-mediated signaling pathway ( Table 3 ). In contrast, metergoline (used to treat hyperprolactinemia), pimethixene (antihistamine), and pizotifen (antidepressant) exhibited higher potency for the PLC pathway than the PLA2 pathway ( Table 3 ). The therapeutic relevance of this functional selectivity is still unknown; as the signaling bias is cell dependent, the transference of in vitro bias to complex systems is a challenge to receptors’ pharmacology. The availability of biased ligands has been proposed, which would allow us to assess this potential problem; therefore, methods like the one reported here would allow identification of novel biased ligands and contribute to advancing the knowledge of their therapeutical/physiological relevance. 13
Even though there are available nonradioactive kits for measuring either inositol phosphate formation or arachidonic release from different providers, these kits do not allow multiplexing for simultaneously measuring these effectors. Therefore, two independent assays should be carried out in order to quantify both PLC and PLA2 signaling, doubling the time needed for obtaining the same results and leading to putative artifacts, depending on the cells’ response in each of the assays. It has been reported that some 5-HT2A agonists can activate Gi/o proteins, with this activation being responsible for a hallucinogenic effect, 14 but it was later reported that metabotropic glutamate mGlu2 receptors’ coexpression is needed for 5-HT2A-mediated activation of Gi/o proteins. 15 The host cell line employed in this work (CHO cell line) is devoid of endogenous expression of metabotropic glutamate mGlu2 receptors. 16 Therefore, 5-HT2A receptors did not activate Gs or Gi/o proteins, hampering a new multiplex for measuring cAMP, but this could be achieved by detecting cAMP levels from cell lysates by using any of the different approaches available (homogeneous time resolved fluorescence [HTRF]), alphalisa, enzyme-linked immunosorbent assay [ELISA], RIA radioimmunoassay [RIA]).
In summary, we have developed a miniaturized and robust multiplex 96-well plate assay that allowed us to simultaneously analyze the PLA2 and PLC effector pathways. The suitability of the method to be employed in HTS campaigns was tested using the Prestwick Chemical Library. The identified hits are known to interact with the target receptor studied, and the assay appears to have detected previously undiscovered functional selectivity for some of the hits. The assay also enabled the detection of previously reported negative cooperativity for clozapine. This novel multiplex methodology will allow the detection of 5-HT2A ligand functional selectivity in the early drug discovery process and provides a new avenue for finding pathway-selective compounds that could serve as pharmacological tools to unravel the pathophysiological relevance of the PLA2 and PLC signaling pathways.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Ministerio de Ciencia e Innovación (SAF2009-13609-C04-01), Ministerio de Economía y Competitividad (SAF2014-57138-C2-2R), and Innopharma project (PI12/00742; Ministerio de Economía y Competitividad–FEDER). A.I. was recipient of an FPI fellowship from Ministerio de Ciencia e Innovación. S.L. was recipient of an FPU fellowship from Ministerio de Educación.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.