
Abstract
The TonB-dependent Gram-negative bacterial outer membrane protein FepA actively transports the siderophore ferric enterobactin (FeEnt) into the periplasm. We developed a high-throughput screening (HTS) assay that observes FeEnt uptake through FepA in living Escherichia coli, by monitoring fluorescence quenching that occurs upon binding of FeEnt, and then unquenching as the bacteria deplete it from solution by transport. We optimized the labeling and spectroscopic methods to screen for inhibitors of TonB-dependent iron uptake through the outer membrane. The assay works like a molecular switch that is on in the presence of TonB activity and off in its absence. It functions in 96-well microtiter plates, in a variety of conditions, with Z factors of 0.8–1.0. TonB-dependent iron transport is energy dependent, and the inhibitory effects of the metabolic inhibitors carbonyl cyanide m-chlorophenylhydrazone, 2,4-dinitrophenol, azide, cyanide, and arsenate on FeEnt uptake were readily detected by the assay. Because iron acquisition is a determinant of bacterial pathogenesis, HTS with this method may identify inhibitors that block TonB function and constitute novel therapeutics against infectious disease caused by Gram-negative bacteria.
Introduction
Iron acquisition is vital for bacteria from its role in energy production, DNA synthesis, intermediate metabolism, oxygen detoxification, and nitrogen fixation. In the host environment, bacterial pathogens must acquire iron to proliferate. Their primary mechanism is secretion of siderophores that chelate ferric iron. 1 Enterobactin, 1 the native siderophore of Escherichia coli and several other Gram-negative bacteria, is a prototypic iron chelator with the strongest known affinity for Fe+++ (K = 1052 M–1). 2
Ferric enterobactin (FeEnt; 719 Da) utilizes FepA, an outer membrane (OM) protein that binds and internalizes it by active transport. 3 The detailed mechanism of FeEnt transport through FepA remains uncertain, but the process is controlled by TonB. 4 Active transport across the OM poses an energetic challenge: How does accumulation occur in a membrane containing open porin channels (e.g., OmpF) that preclude an electrochemical gradient? TonB presumably overcomes this problem by transferring energy from the inner membrane (IM) to the OM, which facilitates FeEnt passage through FepA. The TonB C-terminus adsorbs to a conserved site known as the TonB box at the N-terminus of TonB-dependent transporters (TBDTs) such as FepA, and this association is the basis of energy transfer. 4 After FeEnt passes into the periplasm, it binds to FepB, 5 which delivers it to the FepCDG ABC transporter 6 in the IM, which hydrolyzes ATP as it transports FeEnt into the cytoplasm.
We developed and optimized spectrofluorimetric methods for high-throughput screening (HTS) to measure TonB-dependent FeEnt iron uptake through fluorescently labeled FepA in microtiter plates. The assay may screen for inhibitors, with high Z factors that demonstrated sensitivity, flexibility, and reproducibility. We observed FeEnt uptake with this microtiter test and validated its ability to identify inhibitors of TonB action.
Materials and Methods
Bacterial Strains, Plasmids, and Media
E. coli strain OKN3 (ΔfepA) 7 was the host for derivatives of the low-copy plasmid pHSG575 8 : pITS23 carries the wild-type E. coli fepA gene; pITS23FepAS271C is its derivative that encodes the substitution mutation A271C. 7 All fepA genes were controlled by their native repressible Fur promoter.
Instrumentation
We originated and optimized fluorescent spectroscopic assays in an SLM/OLIS spectrofluorometer that has an SLM 8000 chassis (Aminco, Lake Forest, CA), upgraded to automatic control by an OLIS cpu, operating system, and analysis software (OLIS, Inc., Bogart, GA). For typical operations, the excitation and emission wavelengths were 490 and 520 nm, respectively. We adapted and performed HTS assays on a Tecan GENios Pro (Tecan, Switzerland) with black round-bottom 96-well plates (Corning, Lowell, MA). The excitation and emission filters were 485 and 535 nm, respectively.
Fluorescence Labeling
After overnight growth in Luria broth (LB 14 ) we subcultured OKN3/pITS23FepAS271C into 25 mL of MOPS minimal media 9 with amino acids (at 100 µg/mL), streptomycin (50 µg/mL), and chloramphenicol (20 µg/mL), and shook the flasks 5.5–6 h at 37 °C, until the midlog phase (~5 × 108 cells/mL). We collected the cells by centrifugation at 8000 × g for 10 min, resuspended the pellets in 10 mL of 50 mM NaHPO4, pH 6.5, repelleted the cells by centrifugation, and resuspended them in 10 mL of the same buffer. We exposed the bacteria to 5 µM fluorescein maleimide (FM) for 5 min at 37 °C, quenched the labeling reaction with 1 mM 2-mercaptoethanol, collected cells by centrifugation, and washed and resuspended them in phosphate buffered saline (PBS). After labeling, we adjusted the cell concentrations from their optical density at 600 nm.
FeEnt Quenching and Uptake
When FeEnt binds to fluoresceinated FepAS271C in living E. coli, fluorescence is quenched. But as the bacteria transport the iron complex and deplete it from solution, fluorescence rebounds. Because OM transport of FeEnt is TonB dependent, this assay may identify inhibitors of TonB action that block iron uptake and thereby prevent fluorescence recovery. E. coli OKN3/pITS23FepAS271C-FM, stored on ice, was warmed to 37 °C for 30 min in PBS plus 0.4% glucose in the absence or presence of inhibitors (see below) and then diluted to 2.5 × 107 cells/mL in 2 mL of the same buffer in a 3 mL quartz cuvette with stirring in the SLM/OLIS spectrofluorometer. We collected initial readings of fluorescence intensity, added FeEnt to the indicated final concentration, and monitored changes in fluorescence emissions during its binding and transport. We normalized the data to account for variations in labeling efficacy, cell number, and so forth, and analyzed the results with GraFit 6.011 (Erithacus Ltd., Middlesex, UK).
Inhibitors
To observe the effects of metabolic inhibitors (carbonyl cyanide m-chlorophenylhydrazone [CCCP], 2,4-dinitrophenol [DNP], azide, cyanide, and arsenate) in the assay, we preincubated fluoresceinated cells for 30 min at 37 °C with serial dilutions of different compounds in PBS plus 0.4% glucose, prior to initiation of the assay; we included the inhibitor at the same concentration in the cuvette or microplate. Bacterial cell concentrations, addition of FeEnt, sample mixing, fluorescence measurements, and data analysis were the same as above. Based on data collected in the presence of sequential concentrations of the inhibitors (five-point dose–response curves), 50% inhibitory values represent the final concentration of the test compound that allowed 50% recovery of fluorescence during the uptake time course; at the 100% inhibition level, we did not observe fluorescence recovery.
Cell and FeEnt Concentrations in Microtiter Plates
The rate of depletion of extracellular FeEnt, and consequently the rate of the fluorescence response, related to the concentrations of cells and FeEnt in the microplate wells. We observed the reaction over a range of cell densities and FeEnt concentrations in a Tecan GENois fluorescence microplate reader. We added labeled cells to a 96-well plate at decreasing densities (2.5–0.07 × 107 cells/mL), in a total well volume of 190 µL of PBS plus 0.4% glucose. After measuring unquenched fluorescence by readings with 485 and 535 nm excitation and emission filters, respectively, we added FeEnt (10 µL) to a final concentration of 200 nM in all the wells with automatic injectors. The high concentration of FeEnt ensured maximal quenching across all cell densities, but [FeEnt] is an adjustable parameter that can vary with cell concentration to achieve different extents and durations of quenching. After shaking the plate for 10 s, we measured the FeEnt transport time course over 110 min by plate readings at 10 min intervals (each read cycle was completed within ~1 min) and analyzed averaged triplicate measurements with GraFit 6.011.
Z′ Factor
Z factor calculations estimated the statistical significance of the HTS results and the viability of assay. 10 Positive and negative controls were used in the calculations to determine the statistical effect size.
Results
Fluorescence Spectroscopic Measurement of TonB-Dependent Transport
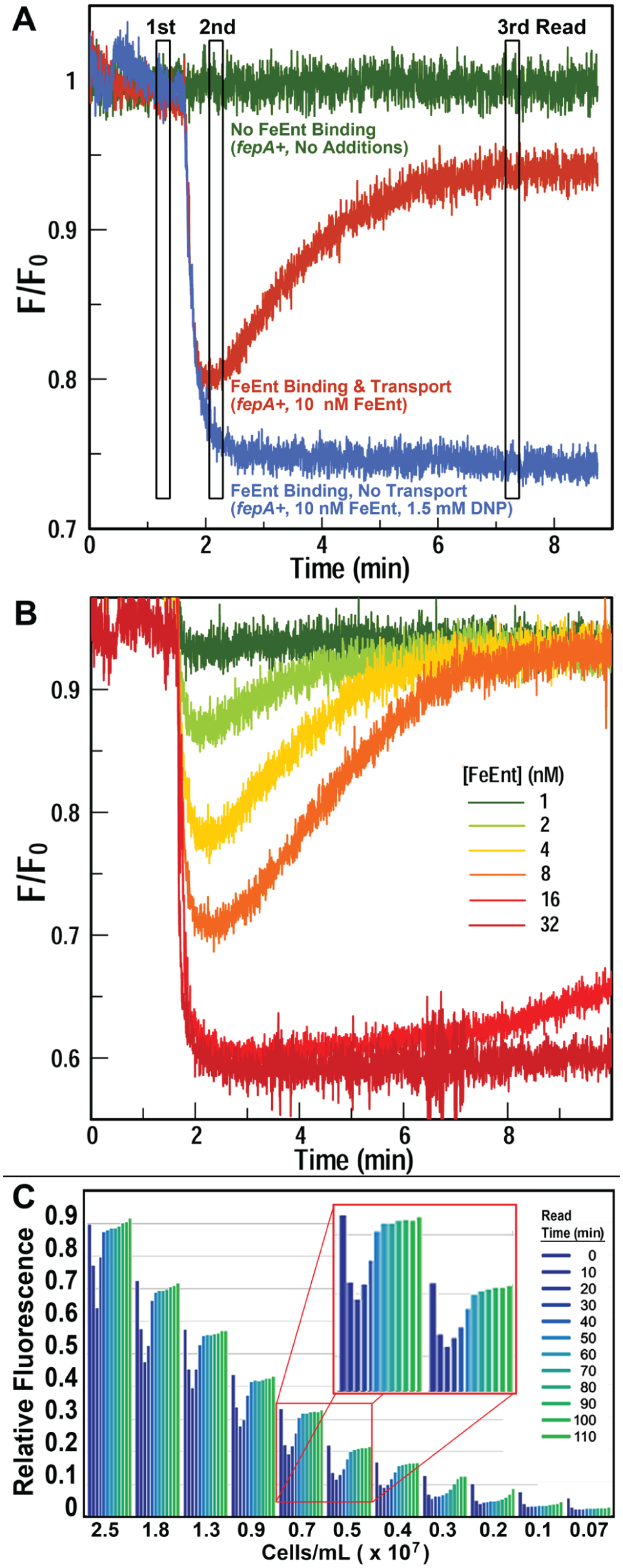
E. coli FepA 3 may be FM labeled in its surface loops, and loop closure around FeEnt during binding quenches emissions from the probe. When fluoresceinated bacteria transport FeEnt it becomes depleted from solution and fluorescence rebounds ( Fig. 1A ). Control experiments in energy- and TonB-deficient bacteria3,11 confirmed the interpretation of these observations. Thus, transport appears as real-time quenching and unquenching of fluorescence intensity, and the extent and duration of quenching depend on the concentrations of bacteria and FeEnt in the cuvette, the temperature, and the incubation time. Variation of these parameters allowed modification of the assay to encompass a spectrum of read times and the inclusion of potential inhibitors.
(
The primary obstacle to accurate measurements with this spectroscopic test is light absorption/scatter by the bacterial cells, which may obscure light emissions by the extrinsic fluorophore. In practice, only dilute bacterial suspensions (i.e., low turbidity) produced usable data. The assay therefore required high-intensity, specific labeling of FepA, which allowed utilization of dilute cell solutions (i.e., less than 2.5 × 107 cells/mL) without loss of the fluorescence signal. We previously defined conditions that maximize specific labeling of the engineered Cys residues in FepA surface loops: 5 µM FM in 50 mM NaHPO4, pH 6.5, 5 min at 37 °C. 3
To observe the dose–response behavior of FeEnt in the fluorescence assay of iron transport through FepA, we tested a range of concentrations (1, 2, 4, 8, 16, and 32 nM) of the ferric siderophore ( Fig. 1B ) in the SLM/OLIS fluorometer. The extent of quenching directly related to the concentration of FeEnt added up to 16 nM; both 16 and 32 nM caused maximum fluorescence quenching. The rate of depletion of extracellular FeEnt was inversely proportional to the amount of FeEnt added; fluorescence recovery occurred within a few minutes for all samples except those exposed to 16 and 32 nM FeEnt (that did not unquench over 8 min).
The SLM/OLIS fluorescence platform established a framework for similar studies in the microtiter format of a Tecan GENios plate reader. Instead of continuous measurements of dynamic fluorescence as seen in the SLM/OLIS, we designed an assay that involved three fluorescence reads ( Fig. 1A ) that allowed evaluation of FeEnt uptake, and its inhibition, in a high-throughput format. The three measurement points (marked in Fig. 1A with vertical bars) occur before addition of FeEnt, after addition of FeEnt, and after incubation that allowed transport of FeEnt. The third read time is flexible and may occur any time after fluorescence recovery. Three time courses ( Fig. 1A ) illustrated the assay concept: addition of PBS, FeEnt, or FeEnt plus an energy poison (DNP) to the cells. The inhibitor did not affect binding but prevented transport, thereby allowing quenching of fluorescence but not unquenching (recovery). The assay acted like a molecular light switch that was turned on by TonB activity and off in its absence.
Fluorescence Measurements in Microtiter Format
We miniaturized the assay in the Tecan Genios fluorometer by optimizing the [FeEnt], the number of cells in the wells, and the transport time. We deposited a 190 µL suspension of OKN3/pITS23FepAS271C-FM in PBS plus 0.4% glucose into microtiter plates, measured starting fluorescence at 535 nm, added 10 µL FeEnt by automatic injection, and monitored its uptake time course. The basic process involved three emission readings at 535 nm: a first reading of fluoresceinated bacteria alone (in the absence or presence of inhibitors), a second reading following automated injection of 5 nM FeEnt, and a third reading after incubation for 30 min at room temperature (RT) to allow FeEnt uptake. With bacterial cells at ~107 cells/mL, we saw transport of 5 nM FeEnt as the expected quenching and recovery of fluorescence intensity in the 96-well plates. Thus, the test correctly functioned in a 200 µL volume, creating a high-throughput platform for identification of compounds that block TonB-dependent FeEnt uptake.
Relative to the SLM/OLIS instrumentation, the high-throughput Tecan instrumentation allowed simultaneous testing of more methodologies and samples in 96- or 384-well black plates. To determine the optimal cell concentration for HTS, we evaluated a decreasing range of cell densities from 2.5 to 0.07 ×107 cells/mL ( Fig. 1C ). When FeEnt was added to the wells at 200 nM, the fluorescence of all samples was quenched; cell density determined the rate of fluorescence rebound. At 2.5 × 107 cells/mL, fluorescence recovered within 40 min, but a 10-fold dilution of the bacteria delayed recovery for an additional hour. Signal intensity also proportionally decreased with the concentration of the fluoresceinated bacterial cells, from 40 at 2.5 × 107 cells/mL to 5 at 0.19 × 107 cells/mL; even in the latter dilute suspensions, FeEnt transport was apparent as quenching/unquenching in sequential plate reads.
Bacterial Viability, Reproducibility, and Optimization of Cell and FeEnt Concentration
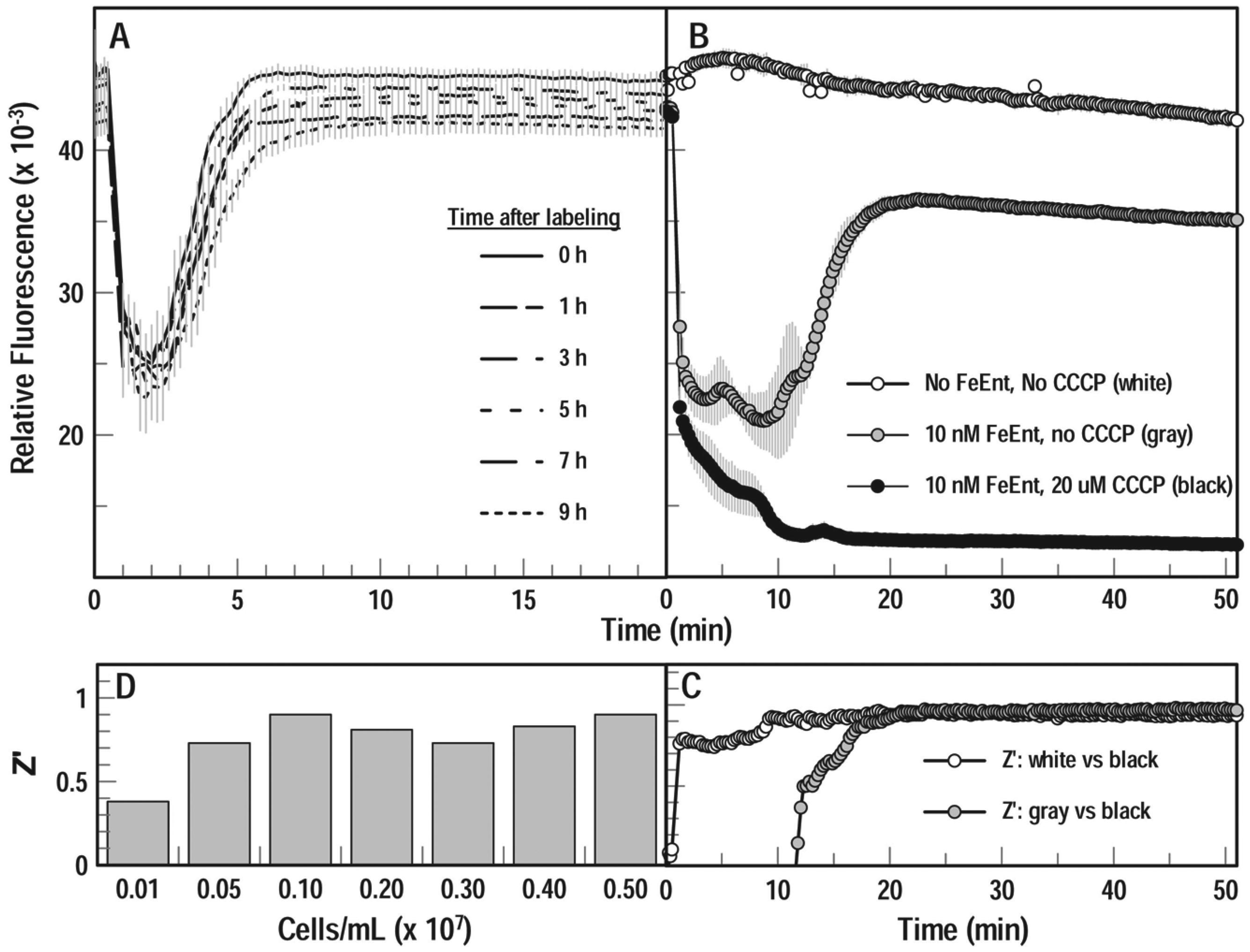
High-throughput screening often lasts hours, so we determined the ability of fluoresceinated bacteria to transport FeEnt after storage in the cold. We labeled cells as described and stored them on ice for 0, 1, 3, 5, and 9 h, and then reconstituted aliquots in PBS plus 0.4% glucose to 107 cells/mL in microplates at RT. After measuring unquenched fluorescence, we dispensed FeEnt into the microwells with automatic injectors to a final concentration of 20 nM, agitated the plate for 10 s, and observed the FeEnt transport time course over 95 cycles. The results showed the resilience of the bacterial cells, with <10% decay in cellular fluorescence (probably from cell lysis) over the 9 h at 0 °C, and otherwise no noticeable effect on FeEnt uptake. FeEnt quenched all samples to ~40% of original fluorescence intensity, and little variation occurred in the speed of fluorescence recovery: all test samples regained their original intensity within 8 min ( Fig. 2A ).
Assay durability, reproducibility, and Z factors. (
Even after extended storage on ice for 11 h, FeEnt uptake by the fluoresceinated cells in microwells recapitulated the concentration dependence of quenching/unquenching ( Fig. 2B ), as previously seen for freshly grown and labeled bacteria ( Fig. 1C ). Furthermore, Z factor calculations from the stored cells were acceptable and amenable to HTS. We added FeEnt (to 10 nM) by automatic injection to cells in microplates, agitated the plate for 10 s, and measured the transport time course. For each condition (no additions, plus FeEnt [10 nM], or plus FeEnt [10 nM] and CCCP [20 µM]), we followed fluorescence in three separate wells of a 96-well plate, made sequential plate reads over 50 min, and averaged the triplicate values ( Fig. 2B ). The statistical significance as determined by Z factors ( Fig. 2C ) validated this extended protocol, which allowed storage of fluoresceinated bacteria overnight on ice prior to use in the assay. Lastly, cell density was an influential parameter of the HTS procedure. For bacterial concentrations from 5 × 106/mL to 5 × 107/mL, we found Z factors in the range 0.7–0.9 ( Fig. 2D ). These results demonstrate the resilience of the cells and reproducibility of the measurements in an HTS microplate format. Some day-to-day variability occurred from one batch of labeled cells to another, but quenching and recovery were as consistent in microplates as in cuvettes.
Effects of Inhibitors
We initially studied the effects of metabolic inhibitors in the SLM/OLIS fluorometer, which revealed the following order of sensitivity: CCCP > DNP > cyanide > azide > arsenate. Preincubation of the bacteria with 0.005 mM CCCP reduced the extent of fluorescence recovery 50% (IC50), and 0.01 mM CCCP prevented any recovery (IC100). We determined IC50 and IC100 for each of these compounds ( Table 1 ). Proton ionophores (CCCP, DNP), electron transport inhibitors (cyanide, azide), and even the phosphate analog arsenate blocked active iron transport.
Inhibitory Concentrations of Energy Poisons.
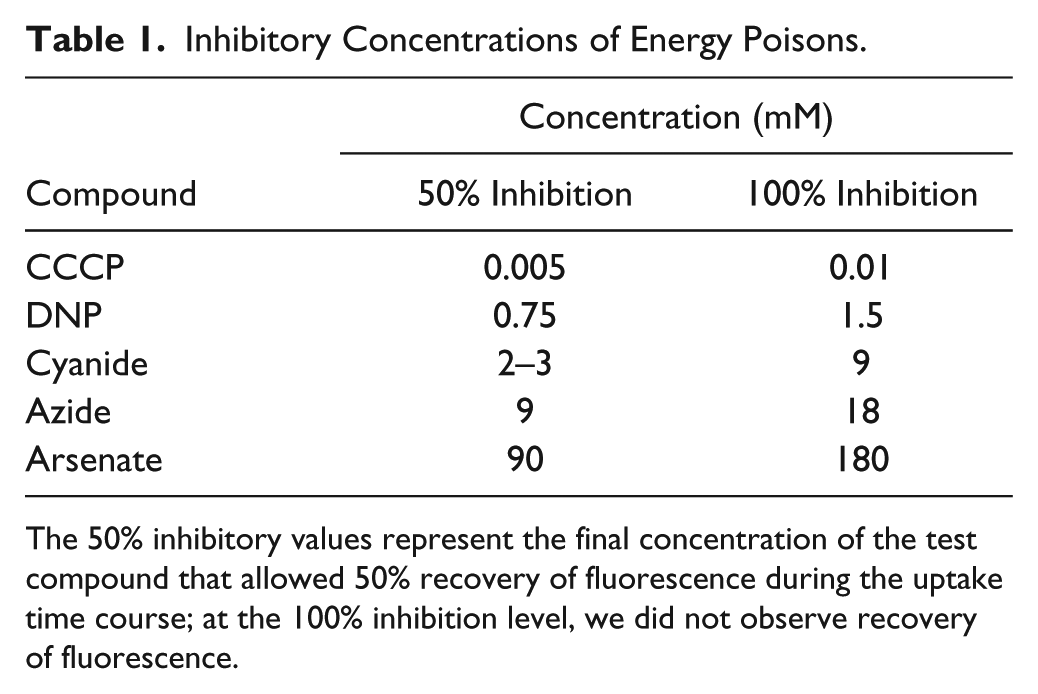
The 50% inhibitory values represent the final concentration of the test compound that allowed 50% recovery of fluorescence during the uptake time course; at the 100% inhibition level, we did not observe recovery of fluorescence.
The microtiter fluorescence assay recapitulated the inhibitory effects of CCCP on TonB-dependent FeEnt uptake ( Fig. 2B ). The fluorescence of the unaltered bacterial cells was stable over the 50 min time course with minimal variation; FeEnt binding by FepAS271C-FM quenched fluorescence, its depletion from solution by transport unquenched fluorescence, and the inclusion of CCCP (20 µM) fully prevented fluorescence recovery. The high Z factors calculated from these samples at the third time point validate the fluorescence HTS approach to identification of inhibitors of FeEnt uptake: compounds that blocked TonB-dependent transport (in this case an energy poison) had Z factors of ~1.0 any time after 20 min ( Fig. 2C ).
Discussion
Fe+++ uptake by Gram-negative bacteria requires TonB-dependent transport through the OM,3,4,11 so inhibitors of TonB action have the potential to inhibit bacterial growth and thereby thwart pathogenesis. The fluorescence spectroscopic microplate HTS assay we report will find compounds that inhibit the activity of TonB. It utilizes live E. coli, and cell-based assays have specific advantages over in vitro tests with purified proteins or reconstituted membrane systems. 12 The fluorescence system is sensitive, durable, and robust: modifications of concentrations, time, and temperature tailor the procedure for accuracy and convenience in 96-well microplates. No conceptual or practical obstacles exist to its operation in 384- or 1536-well plates. The test identified control inhibitors in both cuvettes and microtiter plates, with Z factors near 1.0 in the latter format. Another HTS method that involved bacterial growth inhibition by iron deprivation identified an inhibitor of TonB function, 13 but our test detects chemicals that block the physiologically indispensable TonB-dependent OM stage of iron acquisition. Such compounds are candidates for development as antimicrobial therapeutics. Except for the terminal enzymes of peptidoglycan biosynthesis, the most common antibacterial targets are cytoplasmic, so most antibiotics must pass through both the OM and the IM, which contain efflux pumps that may expel them from the cell. Inhibitors of the TonB-FepA system, on the other hand, may act in the periplasm, may not need to penetrate the IM, and may even act from the outside. These are positive aspects of TonB-dependent iron transport systems as targets for new therapeutics.
The technical details of this HTS assay required optimization in several ways:
Bacterial concentrations. Z factors over a range of cell concentrations showed that 106–107 cells/mL was ideal. At this level, turbidity was minimal, the complete uptake of 10 nM FeEnt occurred within 20–30 min, and Z factors tests were ~0.9. Z factors were lower in tests with more dilute cell concentrations (106/mL); mixing of FeEnt with bacteria in the wells may also become an issue at higher cell concentrations.
Timing of the fluorescence reads. The most basic test involves plate reads before and after addition of FeEnt and after fluorescence recovery (transport of FeEnt). At 107 cells/mL and 10 nM FeEnt, we made readings at t = 0, 5, and 50 min, with the timing adjustable based on the concentrations of the components. This is an end-point assay, in that inhibitors of TonB permanently prevent fluorescence recovery. So, the timing can be delayed for 12–24 h or more after recovery, by incubating the plates at 4 °C.
Stability and viability of fluorescently labeled E. coli. Labeled bacteria stored up to 11 h on ice showed consistent fluorescence intensity, quenching, and recovery in response to FeEnt uptake.
Interference and artifacts in the fluorescence assay. Autofluorescent substances, quenchers, and structural analogs of FeEnt may create artifacts or irrelevant responses in the assay. In general, such compounds are apparent at the outset by lower- or higher-than-expected fluorescence at time zero. However, we have not yet performed the assay in the presence of a wide variety of small molecules, and certain compounds may alter the fluorescence baseline of an HTS process by emitting or quenching fluorescence. A three-read assay will identify both categories of artifactual responses. Competitive, noncompetitive, and uncompetitive inhibitors of FeEnt binding may also prevent FeEnt-mediated quenching of fluorescence. But this effect will occur at the initial stage of FeEnt addition (e.g., no quenching) and create a different result profile than authentic inhibitors of TonB activity. Because FeEnt uptake through FepA is active transport, 4 primary screens will also identify poisons, so secondary screens of TonB function, such as phage and colicin sensitivity tests and siderophore nutrition assays, 14 are needed to exclude chemicals that block electron transport, deplete proton motive force (PMF) or compromise membrane integrity.
Besides creating a new screening assay for inhibitors of TonB activity, the experiments determined the relative potency of different metabolic inhibitors to block TonB-dependent FeEnt uptake. IC50 determinations for CCCP, DNP, cyanide, azide, and arsenate validated the ability of the HTS assay. Inhibitor efficacy related to mode of action: proton ionophores CCCP and DNP were most potent, with IC50 values of 0.05 and 0.75 mM, respectively; the former value confirms a previous study. 15 CCCP is often employed at 0.5–1 mM in physiological experiments, and potentially nonspecific effects may occur with such unnecessarily high excess. Electron transport inhibitors cyanide and azide were less potent (IC50 = 2–9 mM), and arsenate, a phosphate analog that blocks ATP metabolism, was least inhibitory (IC50 = 90 mM).
This is the first method that translates mechanistic knowledge of TonB-dependent systems into an automated assay of membrane transport by living cells, which allows screening for inhibitors of TonB-dependent iron transport. The test organism was E. coli K-12, but the assay is adaptable to any Gram-negative bacterium, assuming a correct genetic configuration that allows fluorescent labeling of the OM iron transporter in a genetic background that is deficient in siderophore biosynthesis. Finally, it’s noteworthy that the assay was not yet tested in practice for interference rates and false positives.
Footnotes
Acknowledgements
The authors thank Aritri Majumdar and Noah Long for helpful discussions.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.