
Abstract
A novel methodology is presented to investigate the organic solvent tolerability of cytochrome P450 monooxygenase BM3 (CYP BM3) mutants. A fluorescence-based continuous-flow enzyme activity detection (EAD) setup was used to screen the activity of CYP BM3 mutants in the presence of organic solvents. The methodology is based on the CYP BM3–mediated O-dealkylation of benzyloxyresorufin to form the highly fluorescent product resorufin. The assay setup not only allows detection of the formed resorufin, but it also simultaneously monitors cofactor depletion online. The EAD setup was used to test the activity of a small library of novel CYP BM3 mutants in flow-injection analysis mode in the presence of the organic modifiers methanol, acetonitrile, and isopropanol. Mutants with enhanced tolerability toward all three solvents were identified, and the EAD setup was adapted to facilitate CYP BM3 activity screening against a gradient of an organic modifier to study the behavior of the small library of CYP BM3 mutants in more detail. The simple methodology used in this study was shown to be a very powerful tool to screen for novel CYP BM3 mutants with increased tolerability toward organic solvents.
Keywords
Introduction
Cytochrome P450 monooxygenases (CYPs) are a family of proteins that catalyze a multitude of reactions. 1 These reactions include hydroxylations, epoxidations, dealkylations, reductions, and other oxidative reactions and are performed on substrates that range from alkanes to complex endogenous molecules. CYPs can introduce atomic oxygen into allylic positions, double bonds, and nonactivated C-H bonds. 2 CYPs are the most important enzymes in human drug metabolism, and it is estimated that approximately 75% of all drugs are metabolized by one or more CYPs. 3 During development of novel drugs, investigation of the metabolic fate of drug candidates is crucial because drug metabolites produced by CYPs may have improved pharmacological activities or may be responsible for toxicity or unwanted pharmacological side effects associated with the parent drug. Therefore, systems are needed that enable facile (bio)synthesis of sufficient quantities of drug metabolites for structural elucidation and pharmacological and toxicological evaluation.
One of the most promising CYPs to be used as a biocatalyst for metabolite production is the microbial CYP BM3 (CYP102A1) from Bacillus megaterium. CYP BM3 is the most active CYP identified thus far, and it is catalytically self-sufficient because it is a natural fusion protein containing the heme domain and reductase on a single polypeptide chain.4,5 Wild-type CYP BM3 is involved in metabolism of long-chain fatty acids and cannot metabolize xenobiotics such as drugs. Rational design and directed evolution approaches have been employed to generate CYP BM3 mutants that can be used for production of human relevant metabolites.6–8 These mutant enzymes can perform regio- and stereoselective bioconversions very efficiently, especially in buffer, at room temperature and under atmospheric pressure. However, many substrates in industrial processes have low water solubility; therefore, organic solvents are often supplemented to enhance solubility and to increase the enzyme-turnover rate. Addition of organic solvents can lead to destabilization of the enzymes or to a strong reduction of enzymatic activity.9,10 Therefore, there is a need to improve organic solvent tolerability of CYP BM3 mutants and to understand underlying mechanisms of effects of organic solvents on CYP BM3. In 2004, a study was published describing creation and identification of CYP BM3 mutants with improved organic solvent tolerability. 11
This study presents a novel setup to investigate the organic solvent tolerability of CYP BM3 mutants. A recently published online screening method to screen for diversity within CYP BM3 mutant libraries 12 has been adapted to create a platform that allows online screening of CYP BM3 activities at different organic solvent concentrations. Furthermore, the setup was altered in such a way that activity of CYP BM3 mutants against an organic solvent gradient can be investigated. The different setups were used to investigate the organic solvent tolerability of a small library of already existing and novel CYP BM3 mutants. The combination of site-directed mutations T235A, R471A, E494K, and S1024E was selected because this was previously shown to improve organic solvent tolerance of CYP BM3 F87A toward DMSO, acetone, acetonitrile (ACN), ethanol, and tetrahydrofuran. 11 These mutations were applied to drug-metabolizing mutants M01, M02, M05, and M11 as templates. We demonstrate here that online screening can be used to study the organic solvent tolerability of CYP BM3 mutants.
Materials and Methods
Chemicals
Benzyloxyresorufin and allyloxyresorufin were synthesized as described previously.12,13 All other reagents and chemicals were of analytical grade and were purchased from standard commercial suppliers unless stated otherwise.
Enzymes and Plasmids
Bacterial CYP BM3 mutants M01, M02, M05, and M11 in the pET28a+ vector were described previously.
14
The first series of triple mutants (MT11, MT112, MT113, and MT114) each contained the R471A, E494K, and S1024E mutations introduced in M01, M02, M05, and M11, respectively. The second series of quadruple mutants (MT133, MT134, MT135, and MT136) contained an additional T235A mutation introduced in the triple mutants.
M01, M02, M05, and M11 mutant proteins were expressed as described previously.
16
pET28a+ constructs of all other mutants used were transformed into Escherichia coli BL21 (DE3) cells using standard procedures. For expression, 600 mL Terrific Broth (TB) medium (24 g/L yeast extract, 12 g/L tryptone, and 20 mL/L glycerol) with 30 µg/mL kanamycin was inoculated with 15 mL of an overnight culture. Cells were grown at 175 rpm and 37 °C until OD600 reached 0.6. Protein expression was then induced by the addition of 0.6 mM isopropyl-β-
Instrumentation
Two Knauer K-500 high-performance liquid chromatography (HPLC) pumps (Berlin, Germany) were used to control 120-mL superloops, made in house, containing enzyme (CYP BM3) and cofactor/substrate (reduced form of nicotinamide-adenine dinucleotide phosphate [NADPH]/alkoxyresorufin), respectively. Two Knauer K-500 HPLC pumps were used for control of the organic solvent concentration in the carrier flow and for delivery of injected samples. CYP BM3 and NADPH/alkoxyresorufin solutions were prepared in potassium phosphate buffer (100 mM; pH 7.4). Both superloops were kept on ice. Flow restrictors were inserted between the pumps and superloops to ensure proper operation of the pumps at low flow rates. Flow restrictors consisted of natural peek tubing and gave back pressures of approximately 50 bar. Pressure limits of the pumps were set 20 bar higher than the working pressure, and VICI Jour backpressure regulators (Schenkon, Switzerland) were inserted after the superloops to prevent damage as a result of possible clogging of the system. For injections of samples, a Gilson 234 autoinjector (Villiers-le-Bel, France) equipped with a Rheodyne (Bensheim, Germany) six-port injection valve (injection loop, 10 μL) was used. To maintain reaction coils at a constant temperature, a Shimadzu CTO-10AC column oven (Duisburg, Germany) was integrated into the system. An Agilent 1100 (Waldbronn, Germany) series fluorescence detector (λex 530 nm; λem 580 nm) was used for monitoring fluorescence. An Agilent 1100 series ultraviolet (UV) detector (340 nm) was used for monitoring NADPH consumption. All hardware was integrated into one system by Kiadis B.V. (Groningen, The Netherlands) and was controlled by software developed by Kiadis B.V.
Metabolism of Alkoxyresorufins by CYP BM3 Mutants
Enzyme activities of the CYP BM3 mutants toward benzyloxyresorufin and allyloxyresorufin were measured according to the method of Burke and Mayer 13 with modifications. These two alkoxyresorufins were selected because they generally showed the highest rate of resorufin production when incubated with CYP BM3 mutants.12,20 Specific activities were determined by measuring resorufin formation off-line on a Shimadzu RF-1501 spectrofluorometer set at λex 530 nm and λem 586 nm. All measurements were performed in duplicate at 24 ºC in a total volume of 1 mL, and 100 mM potassium phosphate buffer (pH 7.4) was used for all experiments. CYP BM3 mutants were introduced in the assay in buffer at a final concentration of 20 nM. Alkoxyresorufin stock solutions were prepared in DMSO and alkoxyresorufins were introduced into the assay for 30 s of preincubation at a final concentration of 10 µM (2% DMSO). Reactions were initiated by addition of 100 µL of 1 mM NADPH, resulting in a final concentration of 100 µM. The increase in resorufin fluorescence was measured over 2 min and the initial linear increases in resorufin fluorescence were used to quantify enzymatic activities.
CYP BM3 Enzyme Activity Screening in Flow-Injection Analysis Mode
A schematic overview of the enzyme activity detection (EAD) system in flow-injection analysis (FIA) mode is depicted in
To test the activity toward the substrate benzyloxyresorufin at fixed percentages of organic solvent, a slightly different EAD setup was used. The superloop containing CYP BM3 was replaced by a loop containing only potassium phosphate buffer (100 mM; pH 7.4). Different CYP BM3 mutants (10 pmol) were introduced by flow injection into a carrier solution consisting of a fixed percentage of organic solvent (the total flow rate was 200 µL/min). Activity of the different CYP BM3 mutants toward benzyloxyresorufin (10 µM) was determined by quantifying the fluorescent signal of the corresponding resorufin product peak formed. NADPH consumption was determined by quantifying the decrease of the UV signal measured at 340 nm.
CYP BM3 Enzyme Activity Screening in Gradient Mode
Activity toward benzyloxyresorufin at varying organic solvent concentrations was investigated by screening benzyloxyresorufin O-dealkylating activity against a gradient of methanol (MeOH). Instead of using a carrier solution with a fixed percentage of MeOH, the two pumps controlling the carrier flow were used to run a gradient of MeOH at a total flow rate of 200 µL/min. The gradient applied was as follows: constant for 5 min at 1% of MeOH, linear increase from 1% to 49% of MeOH, and constant for 5 min at 49% of MeOH. Activity of the different CYP BM3 mutants (50 nM) toward benzyloxyresorufin (10 µM) was determined by monitoring the fluorescent signal of the resorufin product formed. NADPH (100 μM) consumption was monitored by following the decrease of the UV signal measured at 340 nm.
CYP BM3–Mediated Metabolism of the Drug Amitriptyline
Metabolic incubations for the drug amitriptyline were performed in 100 mM potassium phosphate buffer (pH 7.4) with the cytosolic fraction containing 250 nM of the different CYP BM3 mutants at a 100 µM substrate concentration. The final volume of the incubations was 250 µL and the final DMSO concentration varied from 1% to 30%. Reactions were initiated by the addition of 50 µL NADPH, resulting in a final concentrations of 100 μM. The reaction was allowed to proceed for 30 min at 24 °C and was terminated by the addition of 500 µL ice-cold ACN. After centrifugation at 14,000 rpm for 10 min, 5 µL supernatant was analyzed by liquid chromatography (LC)–mass spectrometry (MS). All reactions were performed in duplicate.
Extracts were separated by reversed-phase chromatography using a C18 column (Luna C18[2], 5 μm, 4.6 × 150 mm i.d.; Phenomenex, Amstelveen, The Netherlands) at a flow rate of 0.4 mL/min and a temperature of 25 °C. The gradient was composed of solvent A (0.1% formic acid in water) and solvent B (0.1% formic acid in ACN). The gradient applied was as follows: constant at 5% B for 1 min, linear increase from 5% B to 40% B in 19 min, linear increase from 40% B to 50% B in 0.1 min, constant for 2.8 min, and linear decrease to 5% B in 0.1 min. The column was allowed to re-equilibrate for 12 min at 5% B. Samples were analyzed on an Agilent 1200 Series Rapid resolution LC device equipped with a Time-of-Flight (TOF) Agilent 6230 mass spectrometer. The mass spectrometer was operated at a capillary voltage of 3500 V with nitrogen as the drying gas (12 L/min) and nebulizer gas (pressure 60 psig). The gas temperature was 350 ºC during operation. The TOF mass spectrometer was used in the positive mode and data were acquired using Mass Hunter workstation software (version B.06.00; Agilent Technologies). Standard curves of the substrates were linear between 1 and 250 µM (data not shown).
Results and Discussion
This study aimed to develop an alternative approach to screen CYP BM3 mutants for organic solvent tolerability. Therefore, a recently developed continuous-flow bioassay setup that has been successfully used to screen CYP BM3 libraries for diversity was adapted.
Expression and Activity of Novel CYP BM3 Mutants
The first step was to create novel CYP BM3 mutants with increased organic solvent tolerability by introducing mutations previously described by Wong et al. 11 Although expression yields varied among the different mutants (data not shown), at least 50 nmol of CYP BM3 protein was expressed per 600 mL of TB culture for each mutant. The CO-difference spectra did not show a peak at 420 nm, indicating that the mutations introduced did not affect the stability of the CYP BM3 mutants. Cytosolic fractions were used to prepare 5 μM of CYP BM3 stock solutions in KPi-glycerol buffer for all mutants.
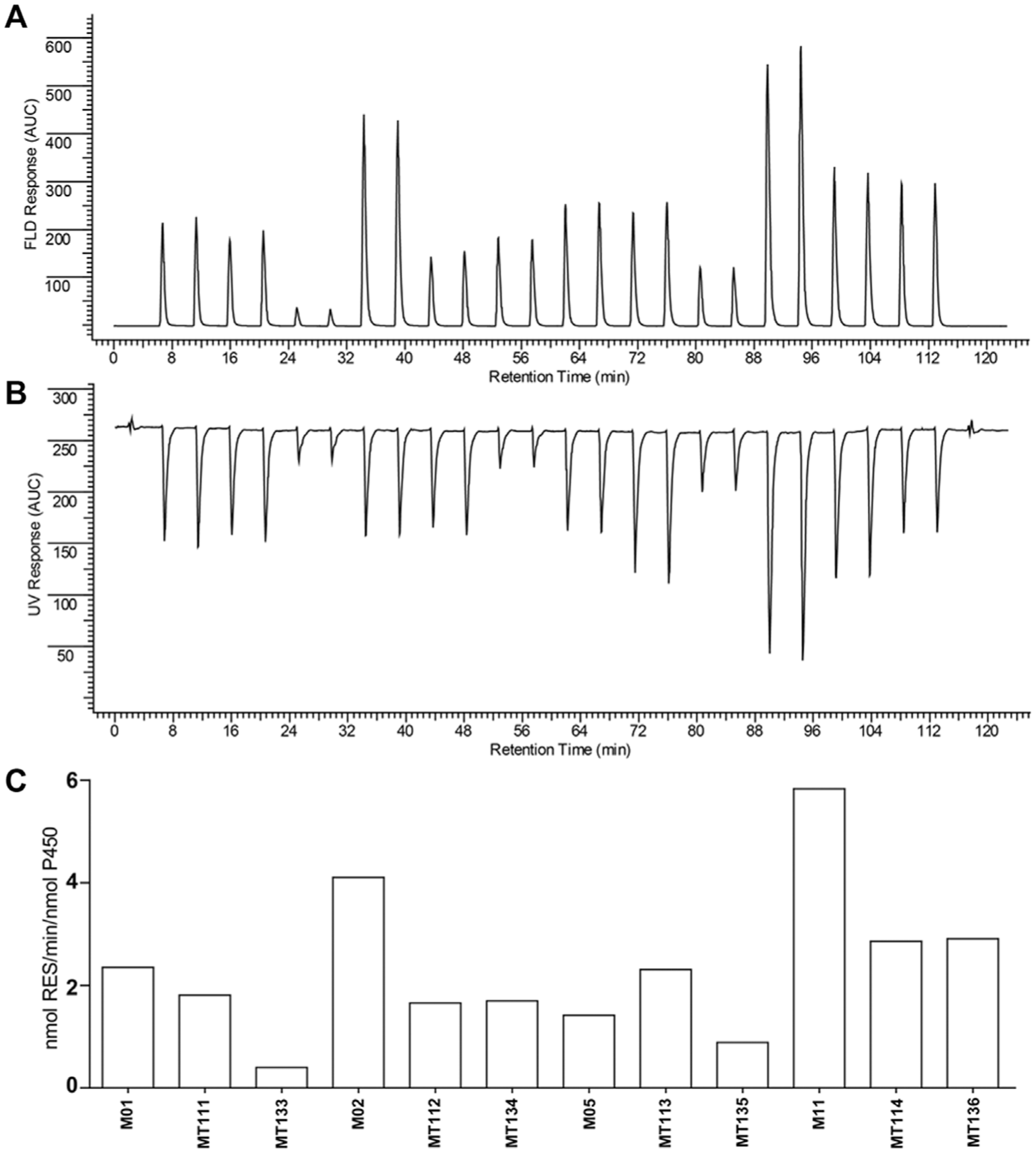
Activities of the four templates and the eight novel mutants toward benzyloxyresorufin and allyloxyresorufin were determined in an off-line fashion to determine which of the two alkoxyresorufins showed the highest O-dealkylation activity. All mutants displayed activity toward the two substrates, but benzyloxyresorufin O-dealkylation (BROD) activity was generally higher than allyloxyresorufin O-dealkylation activity (
For M11, a BROD activity of 5.83 nmol resorufin/min per nmol CYP was detected, which corresponds very well with the value reported previously by Vottero et al. 18 (5.43 nmol resorufin/min per nmol CYP). For M01, M02, and M11, introduction of the combined R471, E494K, and S1024E mutations resulted in a decrease in BROD activity. For M05, introduction of these mutations resulted in only a slight increase in activity. Introduction of the additional T235A mutation resulted in a significant decrease in activity for MT133 and MT135 compared with their templates MT111 and MT113, respectively. For MT134 and MT136, no significant effect on activity was observed compared with their templates MT112 and MT114, respectively.
CYP BM3 Activity Screening in FIA Mode
It was shown previously that the FIA EAD system provided in
(
CYP BM3 Organic Solvent Tolerability Screening in FIA Mode
After having established that the FIA EAD setup could be employed to accurately measure BROD activity and NADPH consumption in parallel, the assay was used to screen the activity of the CYP BM3 mutant library in the presence of different percentages of MeOH, ACN, and isopropanol (iPrOH).
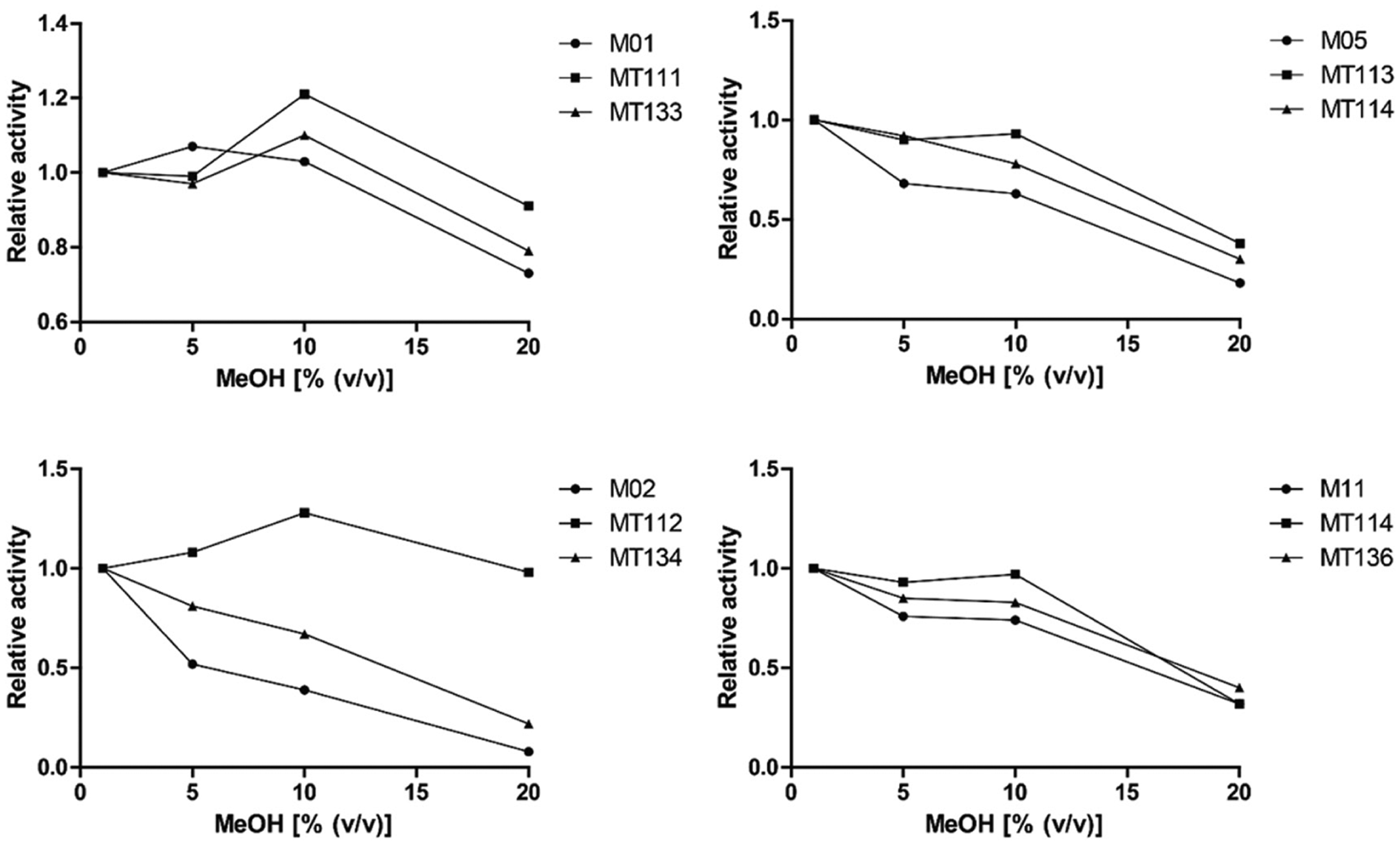
Relative activities as a function of methanol (MeOH) concentration as measured by flow-injection analysis into the enzyme activity detection setup. Relative activities at 1%, 5%, 10%, and 20% of MeOH are calculated as the ratio of the corresponding specific activities to the specific activity in the presence of 1% of MeOH.
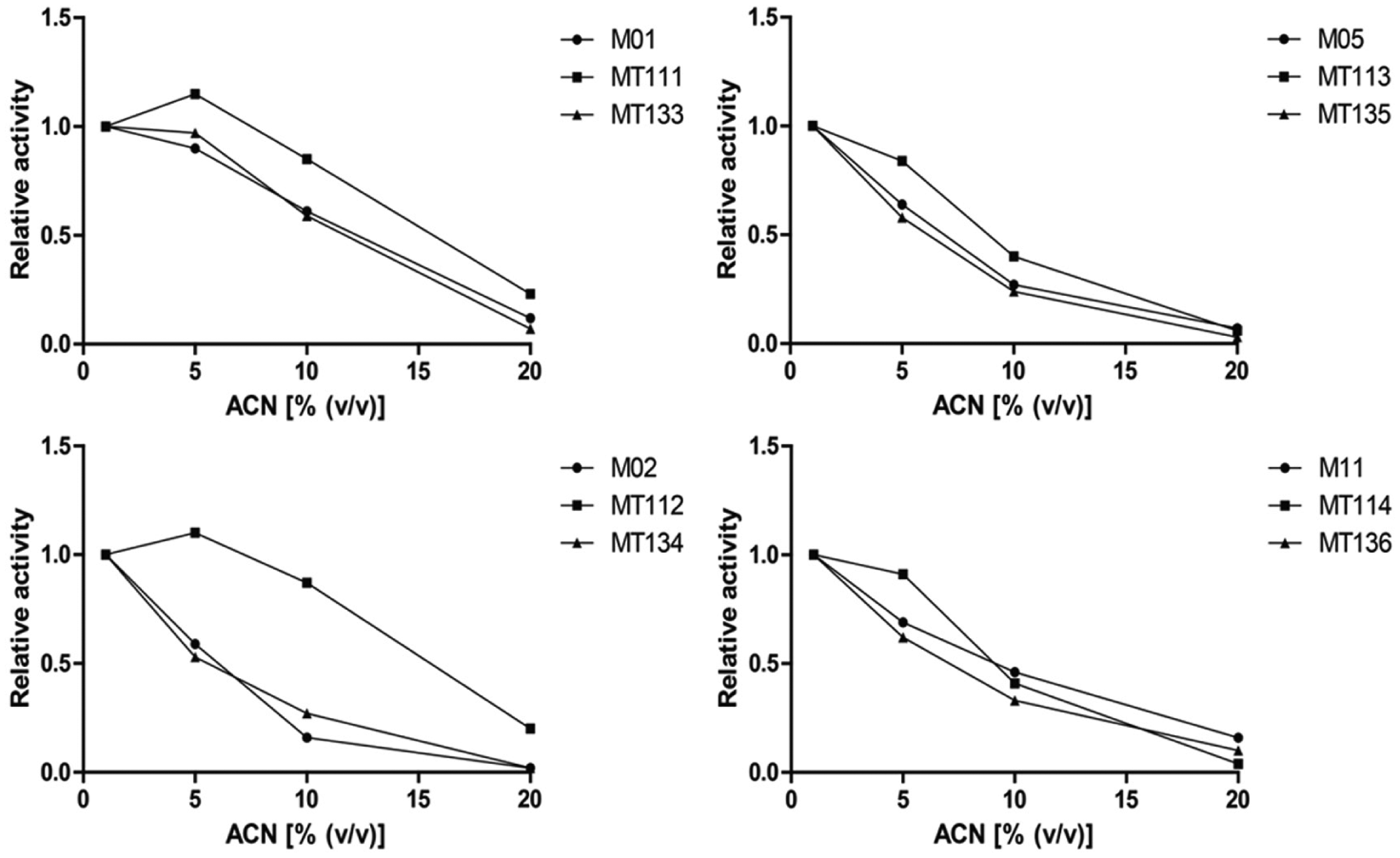
Relative activities as a function of acetonitrile (ACN) concentration as measured by flow-injection analysis into the enzyme activity detection setup. Relative activities at 1%, 5%, 10%, and 20% of ACN are calculated as the ratio of the corresponding specific activities to the specific activity in the presence of 1% of ACN.
As shown in Figure 2 , some of the novel mutants displayed increased tolerability toward MeOH. Although a downward trend for templates M02, M05, and M11 is seen in relative activity with an increasing percentage of MeOH, the corresponding triple mutants (MT112, MT113, and MT114, respectively) display an increase in relative activity at 5% and 10% of MeOH. The increase in tolerability toward MeOH is highest for MT112, the triple mutant of M02, which appeared to be active at 20% MeOH. For the triple mutants of M01, M02, and M05 (MT111, MT112, and MT113, respectively), the activity at 20% of MeOH is higher than that of their corresponding templates. This effect is not observed for the triple mutant of M11. Addition of the T235A mutation to mutants MT111, MT112, MT113, and MT114 (MT133, MT134, MT114, and MT136, respectively) appeared to reverse the tolerability to MeOH to a large extent ( Fig. 2 ).
Figure 3 shows that the triple mutants of M01, M02, and M05 also display an increased tolerability toward ACN compared with their templates. It is again observed that this effect is strongest for the triple mutant of M02. Overall, the negative effect on the activity of all tested CYP BM3 mutants caused by increasing the amount of ACN is much stronger compared with the effect of increasing the amount of MeOH. In the study by Wong et al., 11 the effect of MeOH on CYP BM3 mutants was not tested. However, a stronger inhibition of ACN was observed compared with ethanol.
As shown in
When looking at the effects of increasing the amounts of organic modifier on BROD activities and NADPH consumption, it was observed that a change in activity is always accompanied by a similar change in NADPH consumption (see
Furthermore, consistent with the results of Wong et al., the effects of the stabilizing mutations appear to be dependent on the organic solvent used. However, the specific mechanisms underlying the solvent dependence of effects are still unknown.
CYP BM3 Organic Solvent Tolerability Screening in Gradient Mode
As described above, a number of mutants display an increased organic solvent tolerability toward MeOH, ACN, and iPrOH. Because this effect was most significant for MeOH, it was decided to investigate this in more detail. In the FIA EAD setup, the activity of the different mutants was tested at only four discrete MeOH concentrations. To construct activity curves at a large, continuous range of organic modifier concentrations, the setup was adapted to allow for measurement of parallel BROD activity and NADPH consumption against a gradient of an organic modifier. To test the applicability of the novel gradient EAD setup, the BROD activity and BROD-induced NADPH consumption of the 12 CYP BM3 mutants was screened against a gradient of MeOH. Instead of using a carrier solution with a fixed percentage of MeOH, the two pumps controlling the carrier flow were used to run a gradient of MeOH from 1% up to 49%.
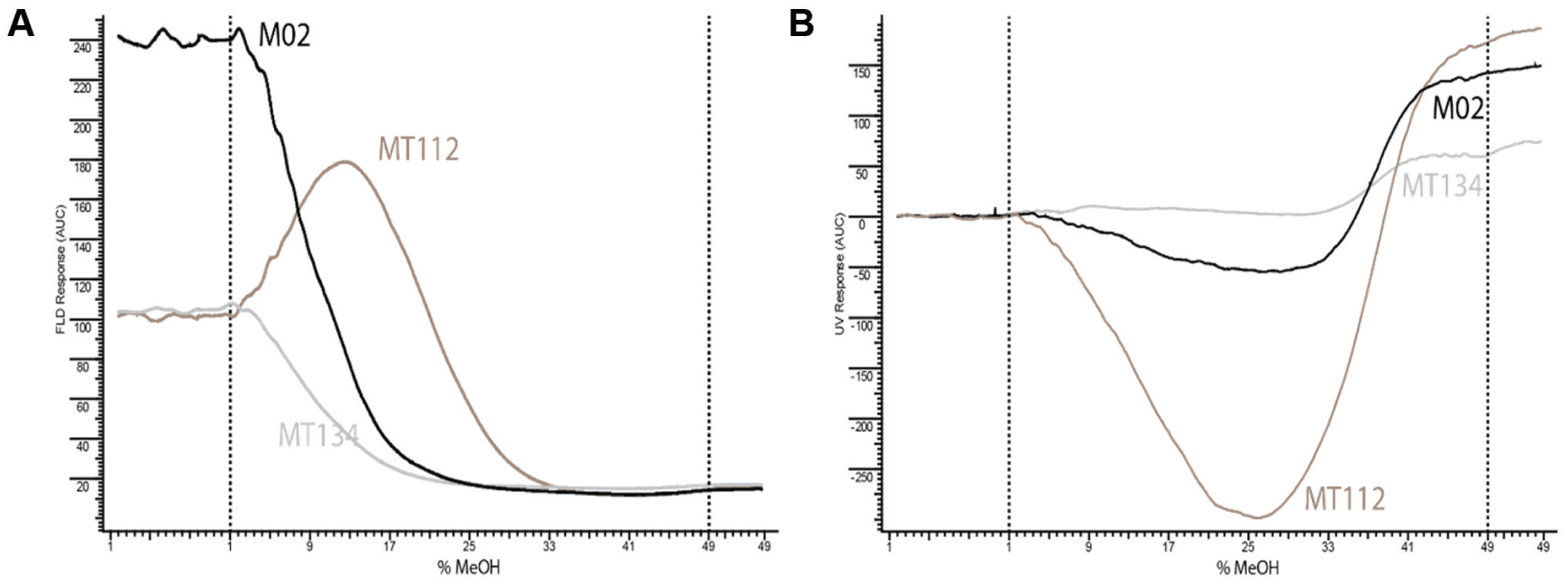
Figure 4A
displays BROD activity profiles of M02 and its triple (MT112) and quadruple (MT134) mutants.
Figure 4B
displays the corresponding BROD-induced NADPH consumption traces. Similar profiles were generated for the other nine CYP BM3 mutants (data not shown). Activity of each mutant was tested in duplicate (two gradient runs were performed) and the raw data files of gradient EAD screening of M02 and MT112 are displayed in
(
BROD activity traces measured were subsequently used to calculate relative activities of the CYP BM3 mutants over a range of MeOH concentrations. Resulting activity curves are depicted in Figure 5 ; compared with their templates, the triple mutants of M01, M02, and M05 display an enhanced tolerability toward MeOH. This effect is greatest for MT112. When comparing results obtained for MeOH in the FIA EAD setup ( Fig. 2 ) with the results obtained for MeOH in the gradient EAD setup ( Fig. 5 ), it is clear that they correlate well. M02, M05, and M11 again display a downward trend in their relative activities with an increasing percentage of MeOH, whereas M01 shows a small initial increase followed by a decrease in relative activity. For M11 and its triple and quadruple mutants, it is again found that the introduction of the mutations has not resulted in an increase in tolerability toward MeOH. To confirm the results obtained in the two EAD setups, the BROD activity of M01, M02, M05, and M11 and their triple mutants was also tested in the traditional microplate reader-based assay format 12 at various concentrations of MeOH (data not shown). The results of this experiment confirmed that the triple mutants of M01, M02, and M05 displayed an enhanced tolerability toward MeOH, whereas this effect was not observed for the triple mutant of M11.
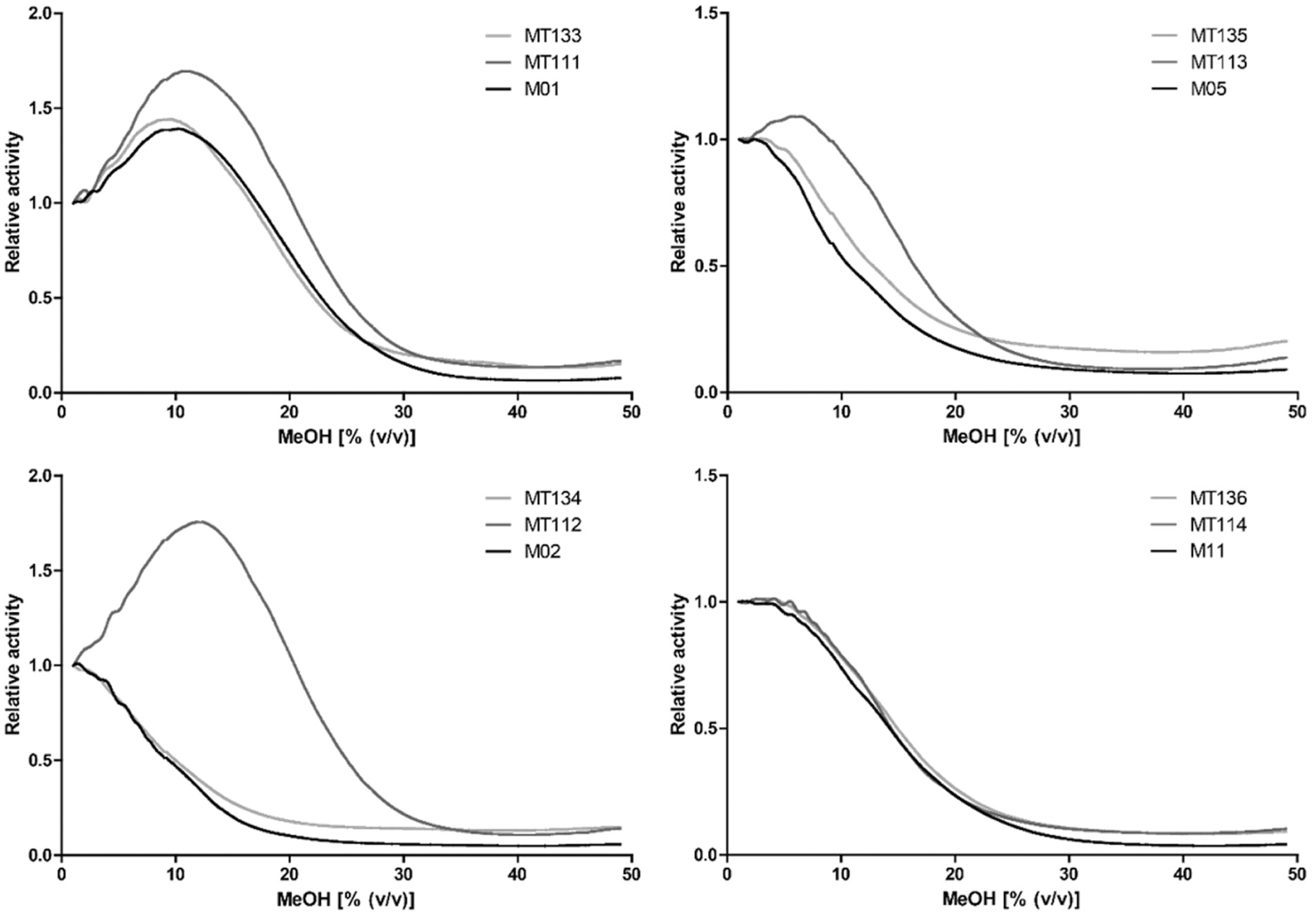
Relative activities as a function of methanol (MeOH) concentration as measured in the enzyme activity detection setup in gradient mode. Relative activities at a given percentage of MeOH are calculated as the ratio of the corresponding specific activities to the specific activity in the presence of 1% of MeOH.
The results presented here clearly demonstrate that the novel gradient EAD setup is an ideal method to study organic solvent tolerability of CYP BM3 mutants. As a proof of principle, the effect of the organic modifier MeOH upon CYP-mediated BROD activity was determined. The setup of the assay can easily be adapted to test the effects of other organic modifiers, other CYP-mediated reactions that are suitable for an online screening assay, or other proteins that can be used in an online screening assay.
CYP BM3–Mediated Metabolism of the Drug Amitriptyline
The novel mutant that displayed the highest increase in organic solvent tolerability was MT112, the triple mutant of M02. To test whether MT112 also displayed an enhanced tolerability toward organic solvents while metabolizing substrates other than the drug-like alkoxyresorufins, an experiment was performed to investigate the M02- and MT112-mediated metabolism of the drug amitriptyline. Amitriptyline was previously shown to be efficiently metabolized by M02 20 in the presence of increasing concentrations of DMSO, because this is the most commonly used organic solvent for poorly soluble substrates. It was previously shown that the mutations applied in this study strongly increased tolerance of CYP BM3 F87A for DMSO. 11
Amitriptyline was metabolized to four products by both mutants (see the
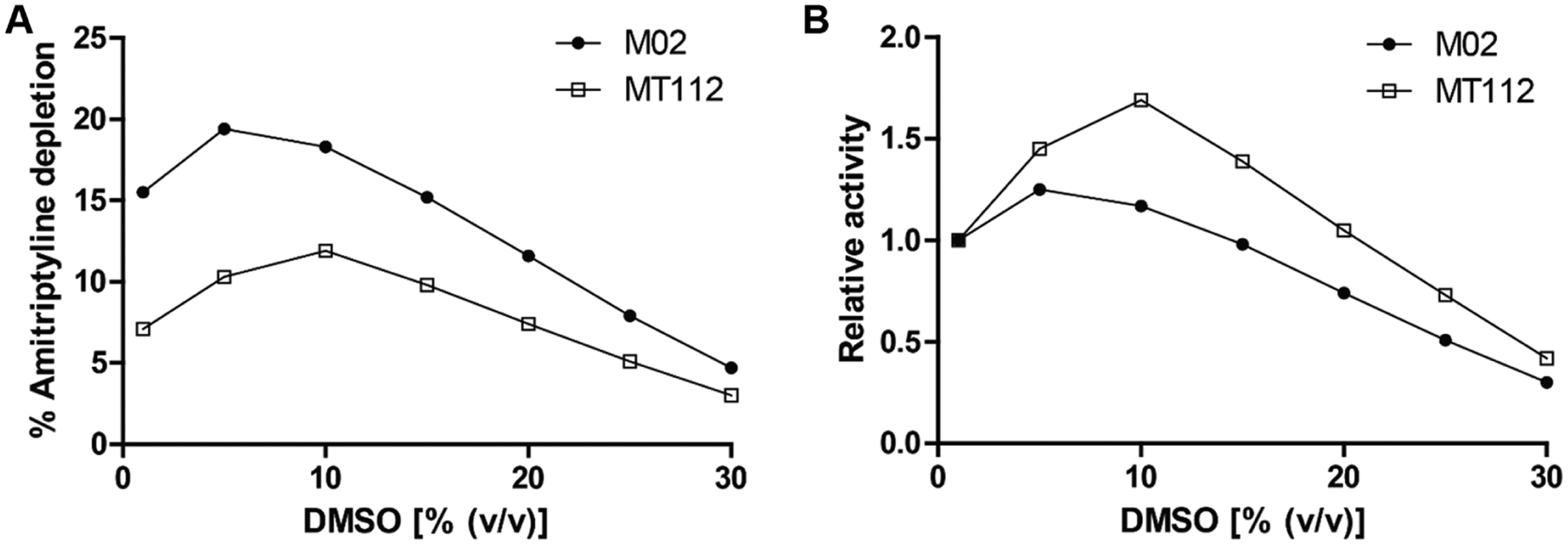
Metabolism of amitriptyline by M02 and MT112. (
In conclusion, the results of this study show that continuous-flow bioassays can be used to screen CYP BM3 mutants for their tolerability toward organic solvents, both at fixed concentrations of organic solvents and with gradients. The bioassay used in this study was based on the CYP BM3–mediated O-dealkylation of alkoxyresorufin to the highly fluorescent product resorufin. In principle, this setup will also be applicable for other fluorogenic substrates such as alkoxycoumarins, which have also been demonstrated to be converted to highly fluorescent products.21–23 Alternatively, chromogenic substrates that are converted to colorimetric products, such as p-nitrophenol ethers, will be applicable for the online activity methodology. 24 The fact that the presented methodology can also be used to measure NADPH consumption shows that the setup is not limited to fluorogenic and chromogenic substrates. However, although NADPH oxidation has been applied as a generic method for measuring CYP activities,25–27 it does not necessarily represent product formation because several substrate/CYP combinations show a high degree of uncoupling, as is the case with metabolism of alkoxyresorufins by CYP BM3 mutants. The most generic method therefore would be online detection of metabolites by MS, as has been successfully applied for analysis of products formed by CYP BM3–mediated metabolism of steroids and p38α kinase inhibitors.28,29 However, this methodology requires availability of expensive mass spectrometers, which will be less commonly available for screening purposes than fluorometric or spectrophotometric detectors.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.