
Abstract
Stem cell research has been progressing rapidly, contributing to regenerative biology and regenerative medicine. In this field, small-molecule compounds affecting stem cell proliferation/differentiation have been explored to understand stem cell biology and support regenerative medicine. In this study, we established a multiparametric screening system to detect bioactive compounds affecting the cell fate of human neural stem/progenitor cells (NSCs/NPCs), using human fetal hippocampal NSCs/NPCs, HIP-009 cells. We examined effects of 410 compounds, which were collected based on mechanisms of action (MOAs) and chemotypes, on HIP-009’s cell fate (self-renewal, neuronal and astrocytic differentiation) and morphology by automated multiparametric assays and profiled induced cellular phenotypes. We found that this screening classified compounds with the same MOAs into subgroups according to additional pharmacological effects (e.g., mammalian target of rapamycin complex 1 [mTORC1] inhibitors and mTORC1/mTORC2 dual inhibitors among mTOR inhibitors). Moreover, it identified compounds that have off-target effects under matrix analyses of MOAs and structure similarities (e.g., neurotropic effects of amitriptyline among tri- and tetracyclic compounds). Therefore, this automated, medium-throughput and multiparametric screening system is useful for finding compounds that affect the cell fate of human NSCs/NPCs for supporting regenerative medicine and to fingerprint compounds based on human stem cells’ multipotency, leading to understanding of stem cell biology.
Keywords
Introduction
Cytological phenotypic profiling based on a multiparametric high-content assay system is a powerful tool for characterization of compounds.1,2 The phenotypes can be employed to classify bioactive compounds based on their mechanisms of action (MOAs) and also to detect or predict novel targets of compounds of which MOAs have not been clarified. By using human stem cells, we can make a novel physiologically relevant screening system that characterizes human cell-fate modulating profiles of compounds, because stem cells are pluripotent/multipotent, being different from using conventional cell lines. In the profiling assays with stem cells, detection of plural cell types by using phenotypic multicolor imaging with differentiation markers fits for this purpose. The results of the cell-fate analysis combined with high-content assays will be useful to understand basic stem cell biology and help regenerative medicine.3,4
In the brain, neural stem/progenitor cells (NSCs/NPCs) are unique, lifelong sources of both neurons and glia. Human NSCs/NPCs hold a great promise as a model for studying human neural biology and diseases, as a source of differentiated neural cells for regenerative medicine, and for bioassays for therapeutic efficacy and neuronal toxicity assessment during drug development. There are several types of NSCs/NPCs, such as fetal NSCs, adult NSCs, NSCs derived from embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs), and mesenchymal stem cells. We previously reported useful characteristics of HIP-009 cells, which are human fetal hippocampal NSCs/NPCs. 5 The cells can differentiate into functional neurons and astrocytes: they express functional glutamate receptors, such as N-methyl-D-aspartate (NMDA) receptors, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionate receptors, and kainate receptors; neurons function electrophysiologically.
HIP-009 cells have the following unique features superior to the other human NSCs/NPCs. First, differentiated HIP-009 cells are considered to be a more physiologically relevant model as hippocampal neural cells (in other words, more region specific) than those derived from human ESCs/iPSCs, because HIP-009 cells themselves are originated from human hippocampal NSCs/NPCs. Second, HIP-009 cells can differentiate into both neurons and astrocytes simultaneously at an about equal ratio in the same culture system, while it is difficult for the other NSCs/NPCs. In this study, using these HIP-009 cells’ features as NSCs/NPCs at maximum, we developed a multiparametric phenotypic assay system to screen compounds affecting cell fate (self-renewal of bipotent cells and differentiation into neurons and astrocytes) and morphology (e.g., neurite length and branch number) of HIP-009 cells. The results indicate that the multiparametric screening system can detect compounds having specific cell-fate modulating effects (e.g., neurotropic) and provide a novel fingerprinting method of compounds based on induced cellular phenotypes.
Materials and Methods
Chemicals
We constructed a bioactive compound library consisting of compounds whose MOAs have been known, collecting different chemotypes of compounds on the same MOA series to enable us to identify off-target effects. They were obtained by purchasing from Axon Medchem (Groningen, the Netherlands), Sigma-Aldrich (St. Louis, MO), Selleck Chemicals (Houston, TX), Wako (Osaka, Japan), and Shanghai Haoyuan Chemexpress (Shanghai, China). In addition, some compounds were synthesized in our laboratories (Tsukuba, Japan). We assayed 410 compounds of this library, which were numbered as 1 to 410. The mechanisms of compounds included enzyme (e.g., kinases) inhibitors and activators, agonists, antagonists, and modulators of receptors or channels. All compounds were dissolved in dimethyl sulfoxide (DMSO; Wako).
HIP-009 Cell Culture for Expansion
HIP-009 cells were purchased from GigaCyte (Branford, CT; now PhoenixSongs Biologicals, Branford, CT). Cells were expanded as described in the manufacturer’s instructions. Dishes of 100 mm were coated with 10 µg/mL laminin (Sigma-Aldrich) in DMEM/Ham’s F-12 (1:1; DMEM/F12; Wako). Cells were seeded and grown on these dishes in Neural StemCell Growth Medium (PhoenixSongs Biologicals) added with Neural StemCell Growth Supplement, 10 ng/mL basic fibroblast growth factor (bFGF), 20 ng/mL epidermal growth factor (EGF), 3 µM CHIR-99021, 1 µg/mL laminin, and 30 µg/mL gentamicin (Life Technologies, Carlsbad, CA). The medium was changed every 2 or 3 d, and confluent cells were split for expansion every 4 or 5 d. Cell culture was carried out at 37 °C in a humidified atmosphere of 2% O2, balanced with N2, and 6% CO2. HIP-009 cells at passages 10 were used in assays. This study was performed with the approval of the Eisai Research Ethics Committee.
Validation of Cell-Fate Changes of HIP-009 Cells by Using Isoxazole-9
Creating the cell-fate modulator screening system using HIP-009 cells, we examined if we were able to detect phenotypic cell-fate changes of the cells by a reference compound in the assays. We used isoxazole-9 (N-cyclopropyl-5-[thiophen-2-yl]-isoxazole-3-carboxamide, Isx-9; Tocris Bioscience, Bristol, United Kingdom), which was reported to induce neuronal differentiation in rat hippocampal NSCs/NPCs. 6 Cell culture, immunostaining, imaging, and analysis procedures were basically the same as in the cell-fate screening described below.
Preparation of HIP-009 Cells for High-Content Screening
Expanded HIP-009 cells were harvested and suspended in Neural Transition Medium (PhoenixSongs Biologicals) added with Neural Transition Supplement, 10 ng/mL bFGF, 20 ng/mL EGF, 1 µg/mL laminin, and 30 µg/mL gentamicin and seeded on poly-D-lysine–precoated 96-well plates at 6 × 103/100 µL/well. One day after, the medium was exchanged into assay medium (DMEM/F12 supplemented with 1× N2 supplement [Wako], 1 µg/mL laminin, and 30 µg/mL gentamicin), and compounds were added at four concentrations of 10-fold serial dilution in reference to their cell toxicity (n = 2). DMSO-treated cells were used as a control (n = 6). The final concentration of DMSO was adjusted to 0.1% in all samples. Cells were treated with compounds for 7 d, and an additional 7 d culture was performed without compounds in the assay medium. Medium replacement was conducted every 3 or 4 d.
Immunostaining
After the 14 d assay treatment of cells, they were fixed with 1% paraformaldehyde (Wako) for 30 min at room temperature. After fixation, they were washed with phosphate-buffered saline (PBS; Wako) and permeabilized with 0.2% Triton X-100 (MP Biomedicals, Santa Ana, CA)/1× Blocking One (Nacalai tesque, Kyoto, Japan)/PBS (PBS-B-T buffer) for 30 min at room temperature. Then, cells were incubated with primary antibodies (goat anti-SOX1 [as a neural progenitor marker, 1:1000; R&D Systems, Minneapolis, MN], rat anti-GFAP [as an astrocyte marker, 1:1000; Life Technologies], and mouse anti-MAP2 [as a neuron marker, 1:1000; Sigma-Aldrich]) in PBS-B-T buffer overnight at 4 °C. After washing with 0.1% Tween-20 (Wako)/PBS, cells were treated with secondary antibodies (anti-goat IgG [H+L] Alexa Fluor 488 [1:500; Life Technologies], anti-rat IgG [H+L] Alexa Fluor 594 [1:500; Life Technologies], and anti-mouse IgG [H+L] Alexa Fluor 647 [1:500; Life Technologies]) and Hoechst-33342 (5 µg/mL; Sigma-Aldrich) in PBS-B-T buffer for 60 min at room temperature.
Imaging Analysis
Image acquisition and analysis were performed by CellVoyager6000 (CV6000), an automated high-throughput cytological discovery system with laser-scanning confocal microscopes and image analysis (Yokogawa Electric Corp., Tokyo, Japan). Four channels of excitation laser wavelength (405, 488, 561, and 635 nm) were used. A total of 45 fields per well were scanned at 10× magnification to produce a tiled image for whole-well image acquisition. The CV6000 quantified numbers of cells positive for SOX1, MAP2, and GFAP among total cells (Hoechst-33342). We calculated ratios of SOX1+, MAP2+, and GFAP+ cell numbers to total cell numbers and used them in analyses. In addition, neurite arborization (neurite length and branching number) of MAP2+ cells was quantified using an algorithm of the CV6000 software. The algorithm segmented the cells into nucleus-adjacent neuronal somas and neurite traces, further determining neurite branching points. This analysis was also applied to GFAP+ cells. Then, the following morphological changes were calculated: neurite length per MAP2+ cell, and process length per GFAP+ cell; number of neurite branching per MAP2+ cell, and number of process branching per GFAP+ cell. Image analysis extracting the cell population and morphology is shown in
Data Analysis
In the preliminary study to validate effects of Isx-9 on phenotypic changes of HIP-009 cells in the assay procedures, one-way analysis of variance was performed, followed by Dunnett’s multiple comparison test as a post hoc test, comparing with the DMSO-treated group. Probability (p) values <0.05 were considered statistically significant.
Collected data of the screening were expressed as fold changes relative to those of the DMSO-treated control group. Criteria for selecting significant changes were set as ≥3× Z-score or ≤-3× Z-score. Data at concentrations showing <50% cell viability were excluded. When the same direction of parameter changes (increases [≥3× Z-score] or decreases [≤–3× Z-score]) was observed at two or more concentrations of a compound, this change was regarded as a specific phenotype of this compound. The changes were ranked as ≥3, ≥5 and ≥10 for increases and ≤-3, ≤-5 and ≤-10 for decreases. The maximum or minimum change of a compound was used in fingerprinting and phenotypic clustering. Dendrograms of compounds’ phenotypes were produced based on clustering and distance measurement by using the complete linkage and cosine correlation, respectively. A hierarchical chemical structure clustering was made based on the Tanimoto similarity between the chemical fingerprinting. The phenotypic and chemical clustering and dendrogram drawing were performed by using TIBCO Spotfire software (TIBCO Software Inc., Palo Alto, CA).
Results
Phenotypic Changes of HIP-009 Cells by Isx-9
Total cell numbers were significantly decreased by Isx-9 at 3.3 (p < 0.05) and 10 µM (p < 0.001; Fig. 1A ). SOX1+ cell number was significantly increased at 3.3 µM (p < 0.05; Fig. 1B ). A trend in an increase of SOX1+ cells was observed at 1.1 µM (p = 0.0932) and 10 µM (p = 0.0600). GFAP+ cell number was decreased concentration dependently ( Fig. 1C ). There was no significant change of GFAP+ cell process length, but a significant reduction of GFAP+ cell process branch number was seen at 10 µM (p < 0.01; data not shown). Isx-9 increased MAP2+ cell number ( Fig. 1D ), and MAP2+ neurite length ( Fig. 1E ) and branch number ( Fig. 1F ) in a concentration-dependent manner. Moreover, a trend in increases of MAP2+ cell neurite length and branch number was seen at 1.1 µM (p = 0.0531 for neurite length; p = 0.0507 for neurite branch number).
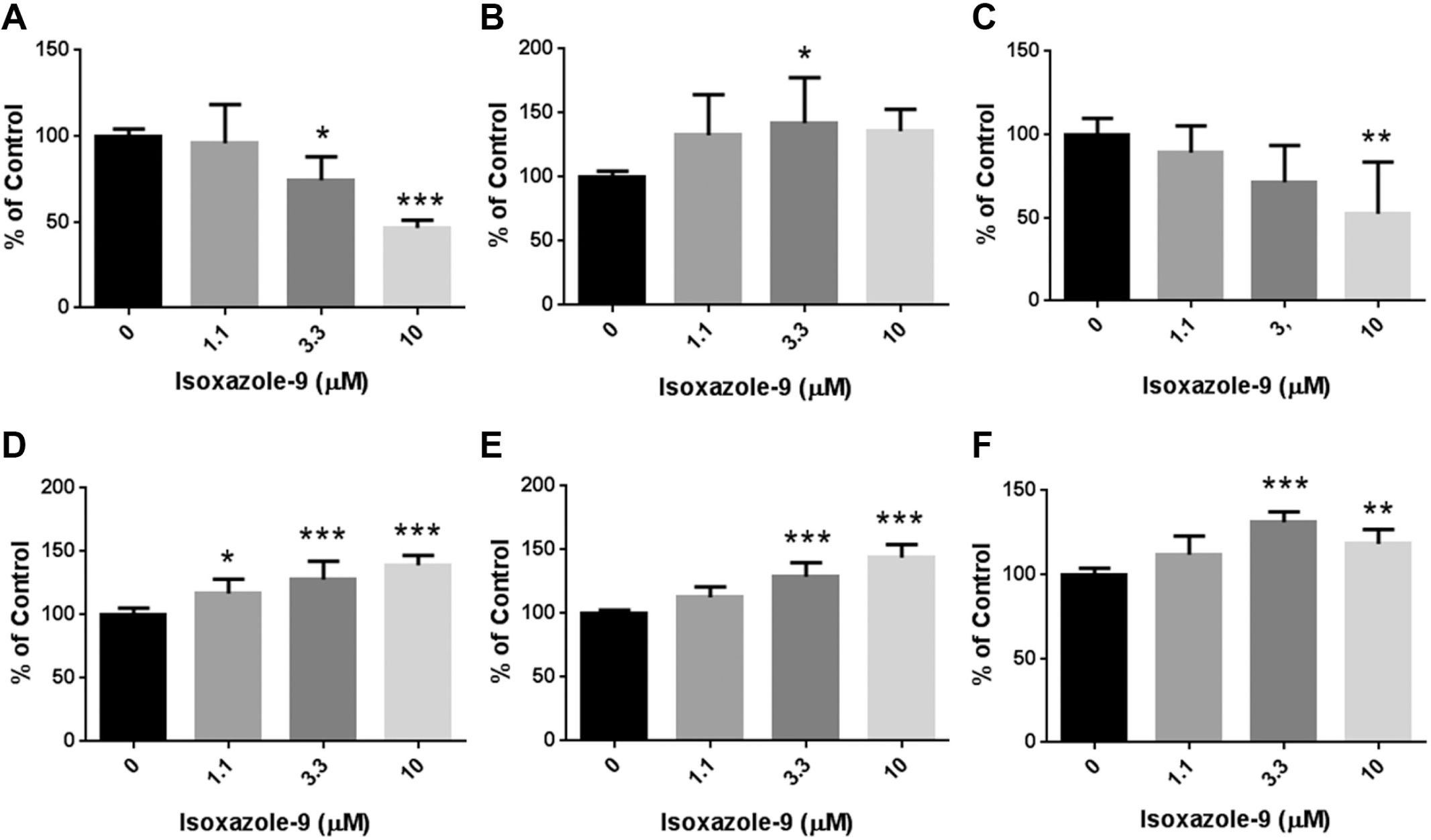
Validation of neurotropic effects of isoxazole-9 (N-cyclopropyl-5-[thiophen-2-yl]-isoxazole-3-carboxamide) on human neural stem/progenitor cells, HIP-009 cells, for the construction of cell-fate screening system. HIP-009 cells were treated with isoxazole-9 (1.1 µM, 3.3 µM, or 10 µM) or vehicle (dimethyl sulfoxide) according to the Materials and Methods section. We quantified total, SOX1+, MAP2+, and GFAP+ cell numbers; MAP2+ cell neurite length and branch number; and GFAP+ process length and branch number. Regarding SOX1+, MAP2+, and GFAP+ cell number, we compared ratios of numbers of cells positive for these markers to total cell numbers. (
Cell-Fate Screening: General Observations
We counted the total cell number as one phenotypic parameter in this screening. Twenty compounds showing <50% cell viability at three or four concentrations were excluded in further analysis. Of the remaining 390 compounds, 148 compounds manifested no phenotypic change.
Figure 2A
indicates a dendrogram of 242 compounds drawn based on the phenotypes of HIP-009 cells. The graph demonstrated that we were able to group compounds into several clusters according to the phenotypes. We analyzed correlations between two phenotypic parameters by drawing scatter plots (
(
Cell-Fate Screening: Specified Observation Examples
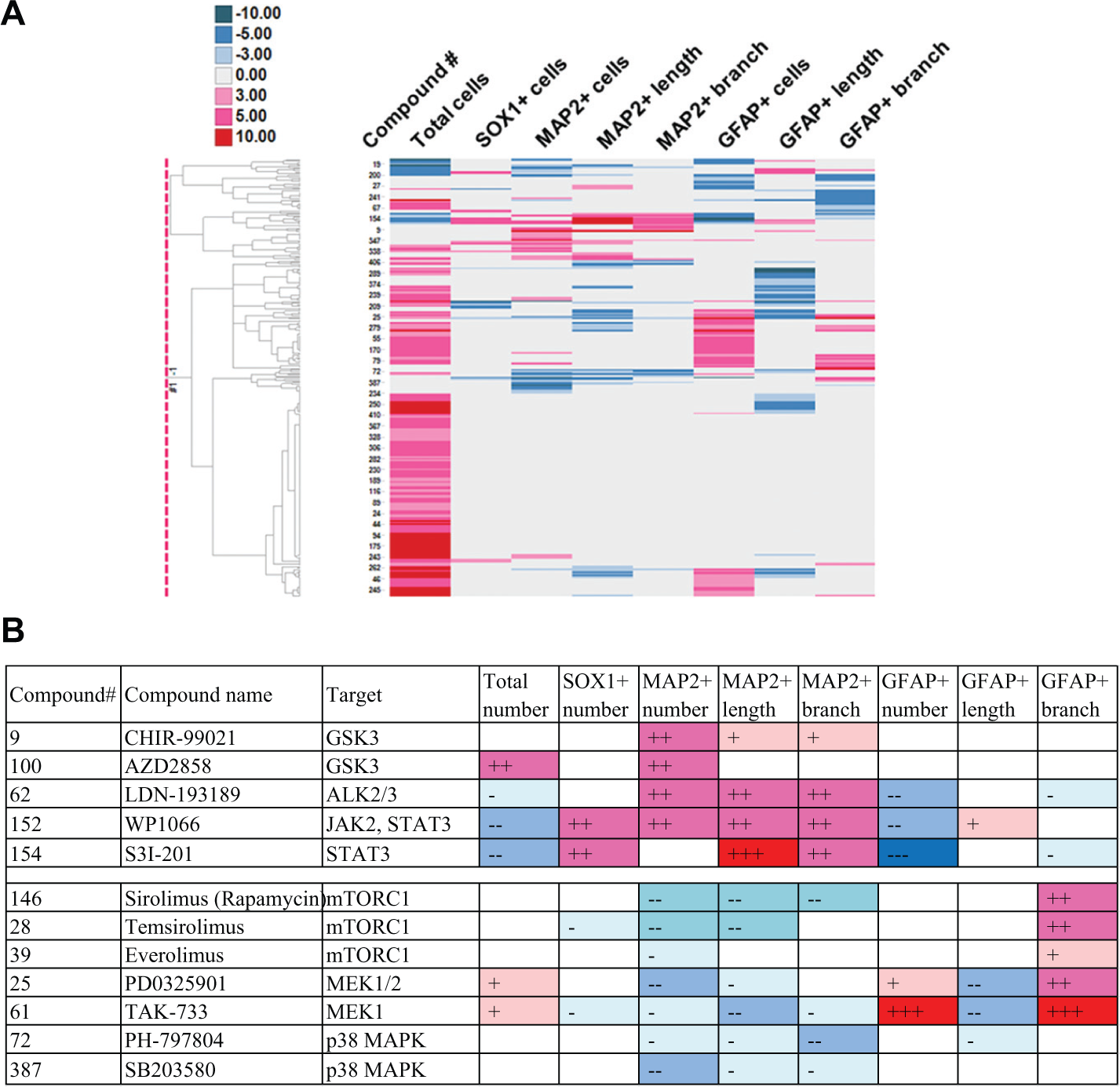
Applying the phenotypic clustering to specific subjects, such as compound groups with similar MOAs and chemical structures, we found several interesting observations, demonstrating further value as a unique screening system. We describe results of some examples hereafter. Compounds selected for specified observations are listed in
Profiling of Tri- and Tetracyclic Compounds Used for Mental Disorders
(
There are several MOAs of tricyclic and tetracyclic antidepressants, including inhibition of serotonin transporter and of norepinephrine transporter, antagonism at 5-HT2 (5-HT2A and 5-HT2C), 5-HT6, 5-HT7, α1-adrenergic and NMDA receptors, and agonism at σ receptors. AMT works as a serotonin-norepinephrine reuptake inhibitor (SNRI),
7
and also as an antagonist at 5-HT2A and 5-HT2C receptors.
8
Profiling of mTOR Inhibitors
Chemical structures of mTOR inhibitors are pictured in
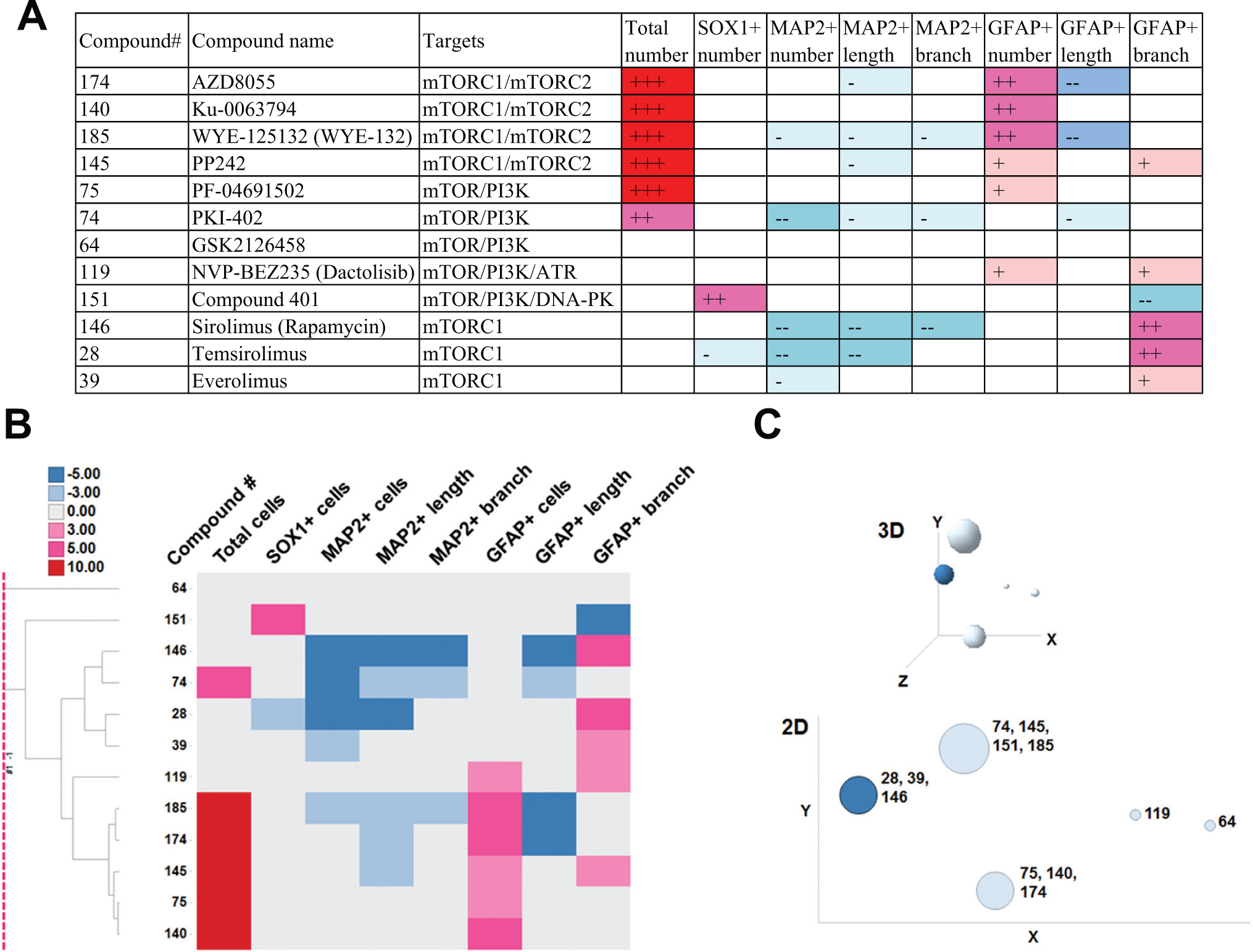
(
Profiling of Dopamine Receptor Agonists
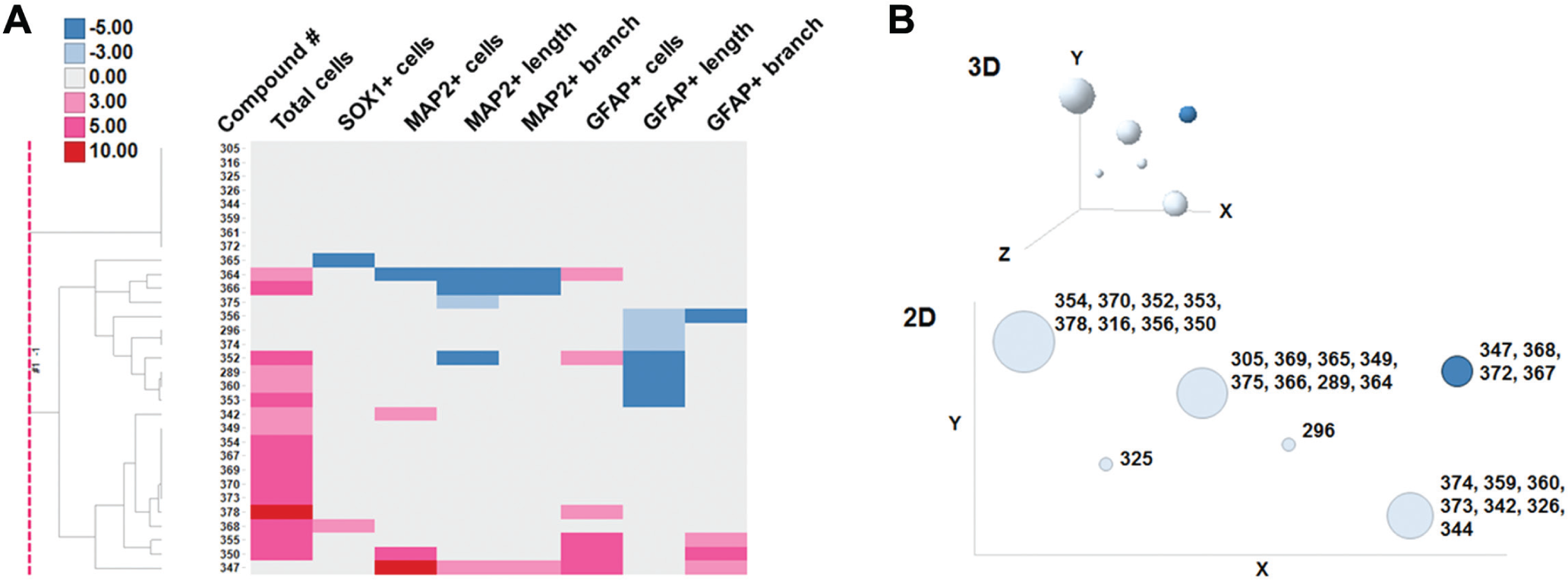
Among the bioactive compounds assayed in this screening, there were seven dopamine receptor agonists. These compounds were classified into two groups based on their structures: ergoline derivatives (#264 [pergolide mesylate], #278 [bromocriptine mesylate], #279 [lisuride maleate], and #298 [metergoline]), and nonergoline derivatives (#282 [ropinirole hydrochloride], #290 [talipexole dihydrochloride], and #291 [pramipexole hydrochloride])). The chemical structures of these compounds are in
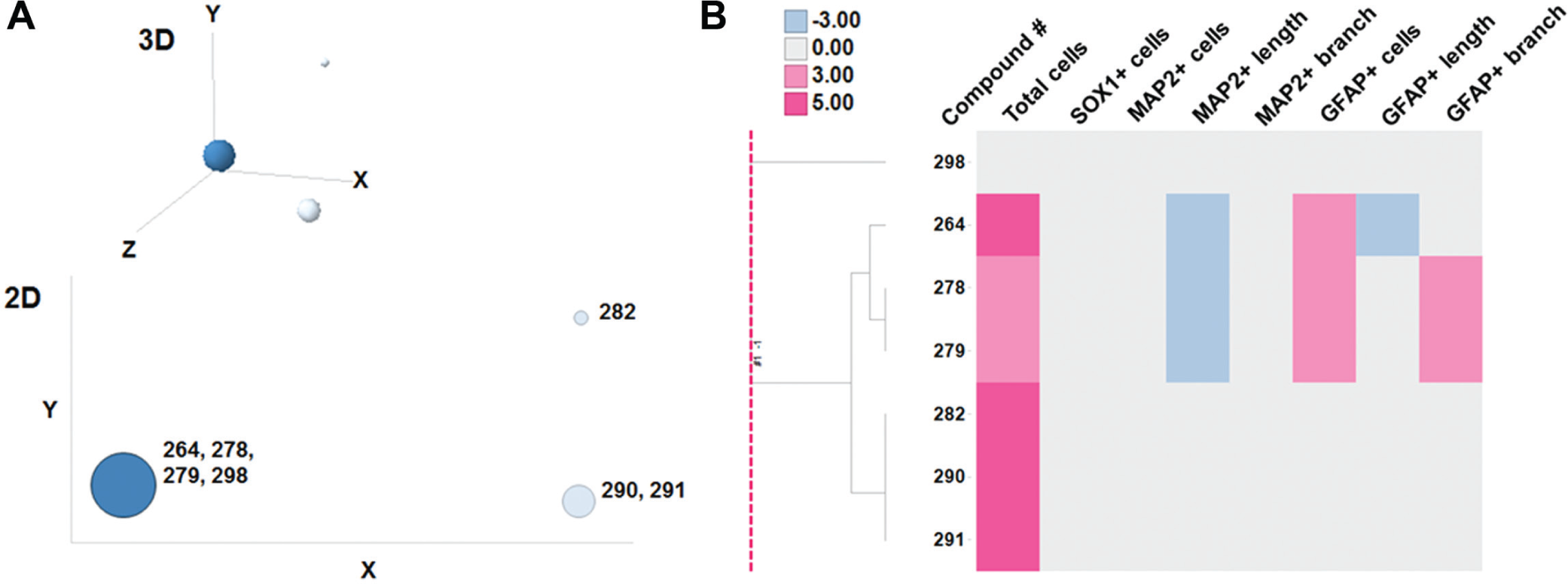
(
Discussion
In constructing the cell-fate multiparametric phenotypic screening, we used Isx-9 as a reference compound to examine if the assay was able to detect HIP-009 cells’ phenotypic changes by this compound as previously reported. Isx-9 was identified as a small-molecule compound to trigger neuronal differentiation in adult rat hippocampal NSCs/NPCs. 6 Moreover, mice treated with Isx-9 showed proliferation and neurogenesis in the hippocampal subgranular zone (SGZ), accompanied with enhanced dendrite arborization including dendrite length and branching. 9 In our study, Isx-9 induced concentration-dependent increases of the MAP2+ cell population and MAP2+ cell neurite length and branch number in HIP-009 cells. Thus, these phenotypes were matched with the in vitro and in vivo changes caused by Isx-9 in rodents, suggesting that Isx-9 is also neurotropic in humans. Furthermore, it was reported that Isx-9 inhibited astrocyte differentiation induced by leukemia inhibitory factor and BMP-2. 7 It appeared that GFAP+ cells were decreased in a concentration-dependent manner in our examination. These results of the validation experiment using Isx-9 would promise us to detect cell-fate and morphological changes of human NSCs/NPCs by bioactive compounds in the procedures of the screening.
We showed representative compounds having an influence on MAP2+ or GFAP+ cell numbers ( Fig. 2B ). These results were in a good agreement of the following published findings: a dominant negative Stat3 mutant increased MAP2 protein levels and MAP2+ cell numbers but decreased GFAP protein levels and GFAP+ cell number in rat fetal NSCs 10 ; cyclic adenosine monophosphate and nerve growth factor–induced neuronal differentiation and neurite outgrowth were inhibited by SB203580 at 1 and 10 µM in rat pheochromocytoma PC12 cells 11 ; CHIR-99021 treatment at 3 µM promoted neuronal differentiation from human iPSC-derived NPCs, 12 and other GSK3 inhibitors, SB216763 (3 µM) and kenpaullone (1 µM), accelerated neurogenesis of a human immortalized NPC line, ReNcell VM 13 ; rapamycin at 5 µM abrogated insulin-induced neuronal differentiation in rat neonatal NPCs 14 ; BMP-4 induced astrocyte differentiation of NSCs/NPCs collected from the subventricular zone (SVZ) of fetal mice 15 and rats 16 ; exposure of BMP-2 to mouse fetal telencephalic NPCs caused developmental fate change from neurons to astrocytes 17 ; microinjection of an MEK inhibitor, U0126, into the caudate-putamen and SGZ resulted in a decreases in NSCs and NeuN+ cells in the SGZ and SVZ of C57BL/6J mice aged 3 mo. 18 The scheme of the pathways with the inhibitors used in this screening is indicated in Figure 6 . As in the references cited above, these pathways seem to be common between humans and rodents. In addition, these results validated the screening system as cell-fate assays using neural stem/progenitor cells.
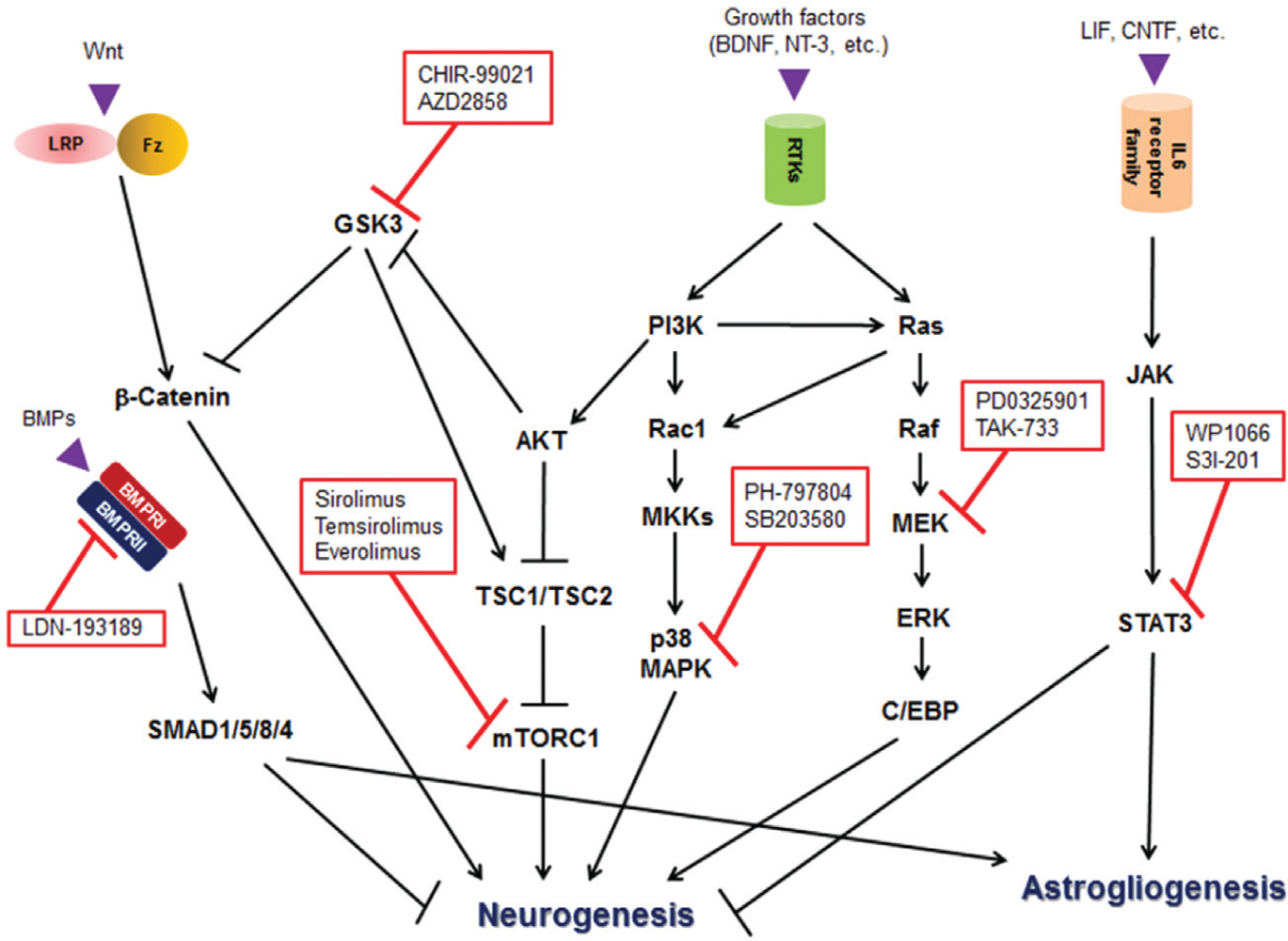
The diagram of the expected regulation of neurogenesis and astrogliogenesis based on the results of the compounds listed in Figure 2B . BDNF, brain-derived neurotrophic factor; BMP, bone morphogenetic protein; BMPR, BMP receptor; CNTF, ciliary neurotrophic factor; C/EBP, CCAAT-enhancer-binding protein; Fz, Frizzled; GSK3, glycogen synthase kinase 3; JAK, Janus kinase; LIF, leukemia inhibitory factor; LRP, lipoprotein receptor-related protein; MEK/MKK, mitogen-activated protein kinase kinase; mTORC1, mammalian target of rapamycin complex 1; NT-3, neurotrophin-3; RTK, receptor tyrosine kinase; SMAD1/5/8/4, SMAD family member 1/5/8/4; STAT3, signal transducer and activator of transcription 3; TSC1/2, tuberous sclerosis complex 1/2.
Results of scatter plots and correlation coefficients of the phenotypic parameters indicated that the correlations between two parameters were not so high each other, excluding a case between MAP2+ cell neurite length and branch number (
We found the unique phenotypic fingerprint of AMT among the tri- and tetracyclic compounds: a prominent increase of MAP2+ cell number with increases of MAP2+ neurite length and branch number (
Fig. 3A
). AMT is a tricyclic antidepressant, and its main MOA is serotonin-norepinephrine reuptake inhibition, also antagonizing activity against 5-HT2A and 5-HT2C receptors. Besides AMT, #349 (imipramine hydrochloride), #370 (dosulepin hydrochloride), and nortriptyline act as SNRIs, but their phenotypic profile was an increase of total cell number. Furthermore, we picked up phenotypic profiles of compounds used for mental disorders whose MOAs are related to serotonin and/or norepinephrine (SSRIs, SNRIs, and NRIs) and to antagonism at 5-HT2A and 5-HT2C receptors. But there were no compounds showing phenotypes similar to AMT (
The neurotropic effects of AMT were reported both in vitro and in vivo. Potentiated neurite formation and axonal growth of primary mouse hippocampal or cortical neurons by AMT at 10 nM were reported by Chadwick et al. 19 In addition, they showed cognitive enhancement with neurogenesis in transgenic Alzheimer’s disease model mice treated with AMT. Jang et al. 20 reported that AMT was a TrkA and TrkB receptor agonist, demonstrating significant neurite outgrowth in PC12 cells at 100 to 500 nM. They reported that neurite outgrowth was not induced by the following tricyclic antidepressants: imipramine, chlorimipramine, prochlorperazine, promazine, trimeprazine or quinacrine. Among them, imipramine, chlorimipramine (#369), promazine (#316), and prochlorperazine (#378) were characterized in our phenotypic screening, and no compounds increased MAP2+ cell number, neurite length, or branch number ( Fig. 3A ). Thus, it is possible that AMT manifests remarkable neurotropic effects via TrkA and/or TrkB receptor in the screening. In other words, this screening system is able to detect neurotropic compounds definitely, although it is a pinpointed, off-target action.
We presented here phenotypic fingerprinting results of compounds having inhibitory activity against mTOR. Sato et al. 21 reported that inhibition of either PI3K by LY294002 (5 µM) or mTOR by rapamycin (10-50 nM) reduced mouse fetal NSCs/NPCs and did not affect self-renewal potency. However, when both PI3K and mTOR were inhibited at the same time, the cells moved into astrocytic differentiation, judged by GFAP expression. An mTOR/PI3K dual inhibitor, PF-04691502, increased GFAP+ cell number in our screening ( Fig. 4A ), which is in agreement with the observation by Sato et al. Four mTORC1/mTORC2 dual inhibitors (AZD8055, Ku-0063794, WYE-125132, and PP242) displayed phenotypes common to those of PF-04691502 (increases of total and GFAP+ cell numbers), which were not induced by mTORC1 inhibitors (sirolimus, temsirolimus, and everolimus). Chronic rapamycin treatment at 100 nM to mouse NSCs resulted in complete abrogation of neuronal differentiation. 22 All mTORC1 inhibitors reduced MAP2+ cell number, which accords with this report. mTORC2 pathway regulation has not been fully elucidated, compared with mTORC1. 23 The present results suggest that target(s) of mTORC2 would be located upstream of mTORC1 in the PI3K pathway, such as AKT.
We can divide dopamine receptor agonists into two groups according to HIP-009 cells’ phenotypic profiles. One group increased total cells and GFAP+ cells, and decreased MAP2+ cell neurite length, whereas the other group increased total cell number only ( Fig. 5B ). Of interest, all compounds in the former group were ergoline derivatives, and the latter’s compounds were nonergoline derivatives. Pertz and Eich 24 described that bromocriptine and lisuride were partial agonists at dopamine D2 receptors, and pergolide showed mixed agonistic activity of dopamine D1 and D2 receptors. On the other hand, Tadori et al. 25 showed that ropinirole, talipexole, pramipexole, bromocriptine, and pergolide behaved as dopamine D2 and D3 receptor full agonists. Metergoline is categorized not only as a dopamine receptor agonist26,27 but also as a 5-HT2C/5-HT1B/5-HT1D ligand. 24 It was reported that bromocriptine had affinity for 5-HT1A receptors and lisuride had affinity for 5-HT1A, 5-HT1D, 5-HT2A, 5-HT2B, and 5-HT2C receptors. Pertz and Eich 24 then wrote that ergoline derivatives were “dirty” drugs. It is thus possible that phenotypic grouping results of these dopamine receptor agonists are due to these pharmacological characteristics derived from their structures.
Heilker et al. 28 described that there was no publication of a high-throughput screening campaign using expandable human NPCs, but they cited two pilot-scale screenings using human iPSC-derived NSCs/NPCs. As differentiation from NSCs/NPCs into neural cells is time-consuming, it is difficult to make cell-fate assays high throughput. We believe that how many valuable and unique parameters we can collect from one assay is very important. Thus, multiparametric screening is fit for this purpose. In the combination of multiparametric phenotyping and characteristics of HIP-009 cells as human expandable NSCs/NPCs, we developed the automated medium-throughput, multiparametric phenotypic screening in this study. This screening is a unique, powerful approach to detect compounds to modify the cell fate of human NSCs/NPCs for helping make progress in regenerative medicine and understand stem cell biology well. In addition, it is very useful as a phenotypic fingerprinting method to categorize compounds according to MOAs and to find compounds with novel, off-target effects among the compounds with the same MOA, using the compound library with a variety of chemotypes.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.