
Abstract
Fidelity of glycan structures is a key requirement for biotherapeutics, with carbohydrates playing an important role for therapeutic efficacy. Comprehensive glycan profiling techniques such as liquid chromatography (LC) and mass spectrometry (MS), while providing detailed description of glycan structures, require glycan cleavage, labeling, and paradigms to deconvolute the considerable data sets they generate. On the other hand, lectins as probes on microarrays have recently been used in orthogonal approaches for in situ glycoprofiling but require analyte labeling to take advantage of the capabilities of automated microarray readers and data analysis they afford. Herein, we describe a lectin-based microtiter assay (lectin–enzyme-linked immunosorbent assay [ELISA]) to quantify terminal glycan moieties, applicable to in vitro and in-cell glycan-engineered Fc proteins as well as intact IgGs from intravenous immunoglobulin (IVIG), a blood product containing pooled polyvalent IgG antibodies extracted from plasma from healthy human donors. We corroborate our findings with industry-standard LC-MS profiling. This “customizable” ELISA juxtaposes readouts from multiple lectins, focusing on a subset of glycoforms, and provides the ability to discern single- versus dual-arm glycosylation while defining levels of epitopes at sensitivities comparable to MS. Extendable to other biologics, this ELISA can be used stand-alone or complementary to MS for quantitative glycan analysis.
Introduction
Biologics, predominantly recombinant immunoglobulins, make up a significant share of today’s pharmaceutical market. There has been an impetus to engineer or enrich for certain terminal glycan motifs, specifically sialylation, because of their effect on the stability and activity of therapeutic glycoproteins. Sialic acid has been reported to exert influence via prevention of serum proteins from degradation, masking antigenic epitopes, resistance on proteolytic degradation, and thermal stability. 1 For example, research on erythropoietin and tissue plasminogen activator has demonstrated the significance of sialylation for increased in vivo half-life.2–4 More recently, not just terminal sialylation but galactosylation5–7 and fucosylation 8 on recombinantly generated biologics as well as intravenous immunoglobulin (IVIG)6,9 have also been shown to have important roles in determining in vivo efficacy. Hence, the need to evaluate the levels of these glycans is a necessary first step for understanding how structure modulates activity in preparations with nuanced differences in motif distribution.
The ability to decipher the glycoform repertoire has benefited from the developments in high-resolution analytical tools such as mass spectrometry (MS), liquid chromatography (LC), capillary electrophoresis (CE), nuclear magnetic resonance (NMR), and combinations thereof.10,11 However, apart from requiring significant investment in specialized expertise, materials requirements, and equipment, data analysis for these analytical methods requires customization and is complex and labor intensive. Finally, obtaining topological information for terminal acidic carbohydrates via these methods entails additional derivatization, which adds to methodological complexity. 12
More accessible methods for glycan analysis have been developed recently but have several limitations. Microarray technology has been adapted to immobilize lectins as probes, on glass or nitrocellulose surfaces, to exploit their innate ability to recognize and bind sugars for in situ glycoprofiling of labeled protein or cells.13,14 Diverse binding specificities of lectins coupled with evanescent field-activated fluorescence and ratio metric/dual-color based detection have enabled mechanistic, organism-wide glycoprofiling and biomarker identification. 13 However, the weak monovalent lectin-glycan interactions demand either saturating concentrations of glycans on lectin microarrays or multivalent presentation of the carbohydrate structure. To overcome this, lectins have been presented in multimeric fashion in the in vitro assays of hemagglutinin, the influenza virus surface protein to glycan receptors, 15 and antibody-lectin sandwich assays wherein antibodies are immobilized on glass surfaces to selectively concentrate specific proteins from body fluids and multiplexed lectins added subsequently for profiling of the captured sample isolates.16–19 Geared toward biomarker discovery, sandwich-type assays require additional preparatory steps aimed to reduce the false positives due to lectin binding to the capture antibodies. This step specifically involves glycans on the capture antibodies to be chemically modified by the addition of a bulky dipeptide that effectively eliminates binding/recognition by lectins through an extensive routine.
We report here an elegant enzyme-linked immunosorbent assay (ELISA) approach that exploits the specificity of lectin-glycan interactions to obtain quantitative information on protein glycosylation using apposing readouts from a pair of lectins. This method uses an easy approach for attaching proteins of interest on microtiter plates, providing an expeditious, low-infrastructure, and minimal resource-requiring method of characterizing glycan epitopes on immunoglobulins. Moreover, the proposed method eliminates sample processing to allow for rapid and efficient scrutiny of multiple N-linked and O-linked glycan structures by the choice of cognate lectins. The assay not only provides linkage information on terminal glycans but has also been demonstrated to be able to distinguish single- versus dual-arm sialylation on complex N glycans on IgGs. This assay thus potentially can provide revealing insights previously considered affordable only via the use of MS/chromatographic analyses. 20
A panel of five plant lectins ( Table 1 ) was chosen to quantify sialylation, fucosylation, mannosylation, and galactosylation of therapeutic immunoglobulins. To demonstrate the practical applicability of the lectin-ELISA assay, we focused on IVIG, a heterogeneous therapeutic glycoprotein mixture, as well as homogeneous recombinant Fc proteins. The quantitative elucidation of differences in glycan expression between differentially glycan-engineered IgG/Fc preparations has been demonstrated herein, in a manner similar to that described by Cao and colleagues, 17 wherein the clustering of lectins using a biotin-avidin architecture enabled presentation in a fashion that closely mimicked the avidity seen physiologically. Neutravidin was specifically chosen to minimize background lectin interaction with glycans on streptavidin. The affinity of glycoproteins to hydrophilic surfaces was used to directly attach the proteins to 384-well plates while the neutravidin–horseradish peroxidase (neutravidin-HRP) complexation before probing the immobilized glycans on the titer plate paved the way for a short (3-h) assay duration. When applied to commercial-grade IVIG, this yielded quantitative data interpretable just as a data set generated by a LC/MS paradigm would have been, showing the same degree of clarity in subtle changes in the terminal glycan epitopes in rapid and high-throughput fashion.
Binding Specificities of the Lectin Panel Used for Glycoform Analyses of Fc and Intravenous Immunoglobulin.

In the current era of antibody therapeutics, the approach presented here highlights the need for integration of information garnered from orthogonal measures; lectins and LC/MS are both usable for robust glycan structure-function relationships, paving the discovery of novel ways in which glycans impinge on therapeutic efficacy.
Materials and Methods
IVIG
Three commercial-grade IVIG preparations, Octagam (Octapharma AG, Lachen, Switzerland), Privigen (CSL Behring, Bern, Switzerland), and Gammagard (GG, Baxter Healthcare, Westlake Village, CA), were tested.
Lectin-Based Microtiter Assay (Lectin-ELISA)
Black 384-well Maxisorp Nunc microtiter plates (Fisher Scientific, Waltham, MA) were coated with recombinant Fc proteins (rFc) or IVIG in phosphate-buffered saline (PBS) (1 µg/mL; 50 µL/well) for 30 min at room temperature (RT). The plates were washed to remove unbound proteins and blocked with PBSAT 1% bovine serum albumin (BSA) and 0.5% Tween-20 in PBS for 20 min. Biotinylated lectins (2–10 µg/mL; Vector Labs, Burlingame, CA) were precomplexed with neutravidin-HRP (Pierce, Rockville, IL) on ice for 20 min and 50 µL of the lectin-neutravidin-HRP complex added to each well; plates were incubated at room temperature for 2 h and washed six times with PBSAT to remove unbound or weakly bound lectins. HRP activity was measured using Amplex Red Peroxidase Assay Kit (Life Technologies, Carlsbad, CA) according to the manufacturer’s instructions. Appropriate negative and positive controls were included, and the tests were run in quadruplicate.
Cloning, Expression, and Purification of the Recombinant Fc (rFc)
For generation of in-cell glycan-engineered rFc, DG44-Fc cells (ProBioGen, Berlin, Germany) were transfected using two GalT vectors, pICHHygroGalT1 and pICHPuroGalT1. Selection medium constituted 10% Ultra Low IgG FBS, with trypsin including either hygromycin B at 300 µg/mL or puromycin at 8 µg/mL (Life Technologies, Carlsbad, CA). In total, 500 mM sodium butyrate, 1 M galactose, and 200 mM ManNAc were used as additives in media and were purchased from Sigma (St. Louis, MO).
The Fc cells transfected with GalT vector (both pICHHygroGalT1 and pICHPuroGalT1) were transferred into six-well plates with FBS and cultured overnight. Selection medium was added after transfection, and cells were monitored for viability (trypan blue exclusion) and clumping (digital imaging) with the Vi-cell instrument (Beckman Coulter, Fullerton, CA). The culture supernatant was purified by protein A column, and protein content was quantified by Nanodrop (Thermo Scientific, Wilmington, DE) before being analyzed by MS and lectin assay.
Subsequently, both stable pools were cloned out by limited-dilution cloning into 6*96-well plates with serum containing media. Twenty-one single-cell colonies were picked from the hygromycin pool and scaled up to six-well plates. Culture supernatant was purified by protein column purification for analysis by mass spectrometry and lectin-ELISA assay. In-cell α-(2, 6) sialylated Fc clones were generated by transfection with human α-(2, 6) sialyltransferase (ST6) constructs followed by purification by the protein A column before analyses by mass spectrometry and the lectin-ELISA assay.
MS for IgG Glycosylation Analysis
IVIG/Fc at 100 mg/mL was diluted into 6 M guanidine (Sigma) to a final concentration of 2 mg/mL. Dithiothreitol was added to a concentration of 10 mM, and the protein was denatured by reducing the disulfide bonds at 65 °C for 30 min. After cooling on ice, the samples were incubated with 30 mM iodoacetamide for 1 h in the dark to carbamidomethylate the cysteine residues. Guanidine, dithiothreitol, and iodoacetamide were all purchased from Sigma. The protein was then dialyzed across a 10-kDa membrane into 25 mM (pH 7.8) ammonium bicarbonate buffer. Trypsin (Promega, Madison, WI) was added to the sample for proteolysis, which was carried out in a Barocycler (NEP 2320; Pressure Biosciences, South Easton, MA). The pressure was cycled between 20 kpsi and ambient pressure at 37 °C for a total of 30 cycles in 1 h.
LC-MS/MS analysis of the tryptic digests was performed on a Dionex Ultimate 3000 chromatography system and a LTQ-XL mass spectrometer (Thermo Fisher Scientific, San Jose, CA). Peptides were separated on a Waters BEH PepMap column (Waters, Milford, MA) using 0.1% formic acid in water and 0.1% formic acid in acetonitrile as the mobile phases.
Glycosylation was quantified for the different isotypes by the area from the extracted ion chromatogram (XIC) for the tryptic glycopeptides as described by Zauner et al. 20
Fc Generation from IVIG
Human IVIG was papain digested to cleave the IgG at hinge region to generate the Fc dimer and Fab monomers. Briefly, commercial-grade IVIG was dialyzed against PBS using Slide-A-Lyzer G2 Dialysis cassettes (Thermo Scientific, Waltham, MA) for 24 h. The buffer-exchanged IVIG (50 mg/mL) was digested with papain (Worthington Biochemical, Lakewood, NJ) for 25 min at 37 °C and the reaction stopped by addition of iodoacetamide (Sigma). The undigested IgG and Fc/Fab monomers were separated by size exclusion chromatography (SEC) using a HiPrep 26/60 S-200HR column (GE Healthcare, Piscataway, NJ). The Fc/Fab peak was further purified on a protein A column (Pierce) to remove the Fab fragments.
In Vitro Glycan Engineering of IVIG and Recombinant Fc
Commercial IVIG or, following the aforementioned purification, Fc fragments were incubated at 37 °C for 48 h with 50 mM UDP-galactose and 20 mU bovine milk β-(1, 4) galactosyltransferase per milligram of substrate. Galactosylated Fc/IVIG were further incubated for 72 h at 37 °C with addition of 8 mM CMP–sialic acid and α-(2, 6) sialyltransferase for sialylation. Recombinant human α-(2, 6) sialyltransferase enzyme was expressed in 293T cells after transient transfection.
Results
Quantitative Analysis of In Vitro Glycan-Engineered rFc Using Lectin-ELISA Assay
Complex biantennary Fc glycan structures on IgGs shown to have pivotal roles in modulating activity of the molecules to which they are attached have been considered refractory to in situ analysis in contrast to the more easily accessible Fab glycans.2,21 Consequently, to understand whether Fc glycans could indeed be captured by the lectin-ELISA, we designed a series of glycoforms of Fc fragments as a first step.
Based on prevalent literature 22 on the structural epitopes recognized by lectins and their binding specificities, a panel of six lectins was chosen for our study. The lectins Erythrina cristagalli lectin (ECL), Ricinus communis agglutinin (RCA), Canavalia ensiformis (ConA), and Aleuria aurantia lectin (AAL) were used to recognize glycan motifs as shown in Table 1 . In addition, Sambucus nigra agglutinin (SNA) and Maackia amurensis lectin (MAL) were used to discriminate between the 2, 6 and 2, 3 glycosidic linkages, respectively. Moreover, we locked the spatial presentation of lectins in a multimeric fashion by precomplexing biotinylated lectins to neutravidin ( Fig. 1 , schematic) to overcome the weak lectin-glycan interaction. This presentation of multimeric Sambucus nigra lectin (SNA) to immobilized Fc glycans provided a dose-dependent signal that exhibited saturating behavior above lectin concentrations of 10 µg/mL ( Fig. 1 , bar graph). All the lectins showed a similar saturating behavior at concentrations above 10 µg/mL; hence, working concentrations for all lectins in the study were collectively restricted to 10 µg/mL or less.
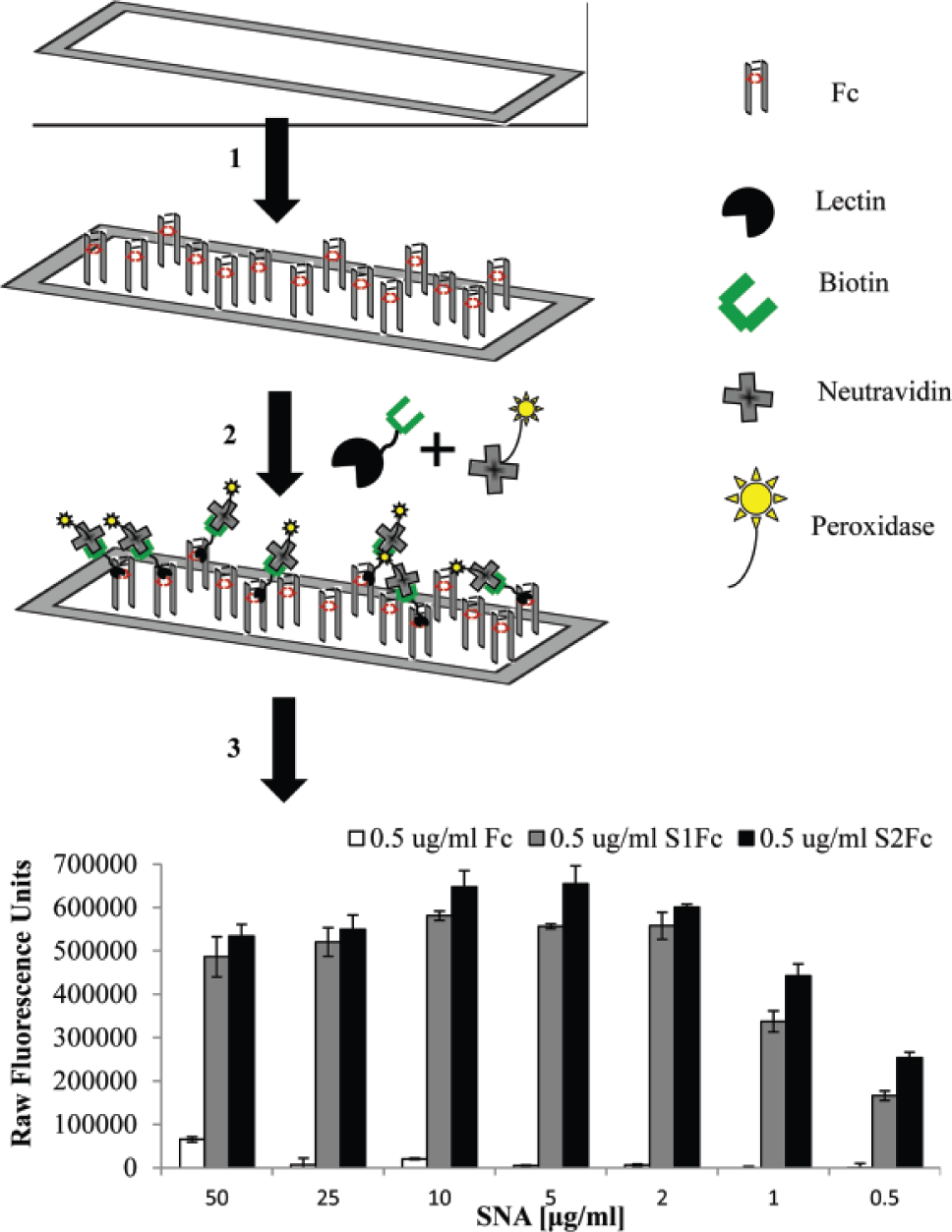
Lectin–enzyme-linked immunosorbent assay (ELISA). A schematic drawing of the microtiter assay for protein glycosylation. The 384-well plates (1) are coated with Fc/intravenous immunoglobulin (IVIG) (2). Precomplexed lectins (biotinylated lectins incubated with peroxidase-conjugated neutravidin) are added to facilitate the multivalent presentation. (3) After unbound lectins are washed off, binding to the plate is visualized after addition of amplex red. The obtained relative fluorescence unit (RFU) values can be plotted against lectin concentration to compare samples with varying glycosylation levels. The shown graph exemplifies the different concentrations of Sambucus nigra lectin (SNA) tested wherein 10 µg/mL, as clearly indicated, was the saturating concentration.
Presentation of lectins as part of a scaffold to improve in vitro binding to cognate receptors described herein is similar to attachment of lectins to agarose (lectin affinity chromatography)23,24 and streptavidin.24,25 Such multimeric presentation 17 has been shown to vastly improve binding, paving the way for lower concentration of lectins amenable for better signal discernment from background, and this was exactly what we observed in our experiments. In addition, the use of neutravidin precluded the engagement of lectins to epitopes in the complexed state as lectins have no recognition sites on neutravidin, allowing an unbiased signal generated exclusively due to recognition of glycan on the protein analyte.
It has been previously reported that saturating concentrations of glycan binding proteins may mask the quantitative measure of relative affinities of protein-glycan interactions. 15 This further lends credence to our proposed range for the precomplexed lectin concentration to be limited within 2 to 10 µg/mL.
Figure 2A
shows the quantitative glycosylation (sialylation, galactosylation, and mannosylation) analysis of rFc, monosialylated rFc (S1Fc), and disialylated rFc (S2Fc) using the lectin assay. Although one would expect that the relative SNA signal from S1Fc would be half that of S2Fc, this is not the case due to the steric hindrance posed by the multimeric presentation of SNA in the assay. Hence, to evaluate the differences in the terminal glycan composition of S1Fc and S2Fc, the lectin ECL that recognizes terminal asialogalactose was used. The difference in relative ECL signals between S1Fc (72%) and S2Fc (5%) is because S1Fc contains one terminal asialogalactose to which the ECL-NA complex binds compared with the absence of free terminal asialogalactose in S2Fc (
Fig. 2A
).
Figure 2C
lists the overall fractional distribution of the different glycan moieties in rFc, S1Fc, and S2Fc based on LC-MS/MS analysis (
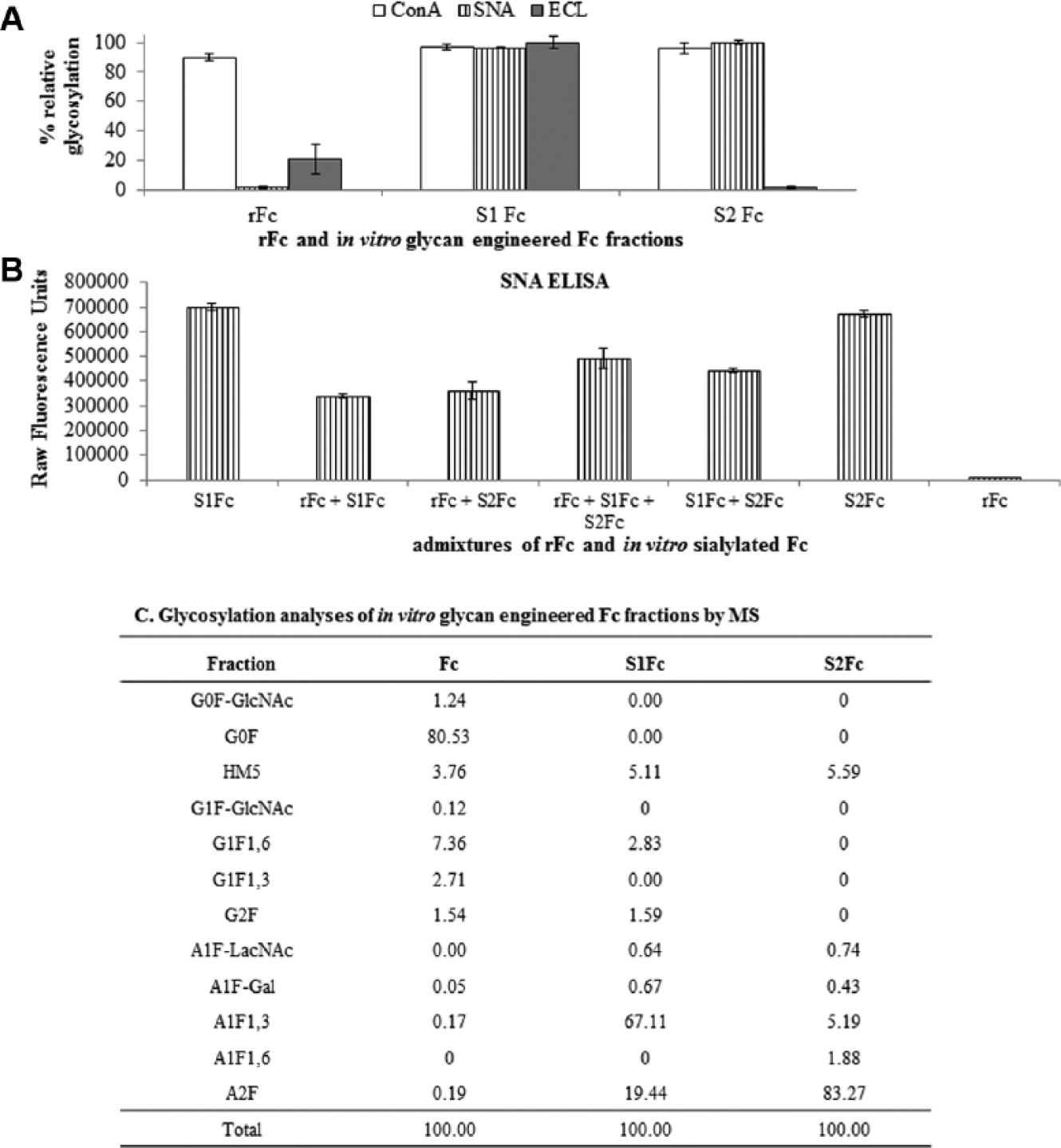
Lectin enzyme-linked immunosorbent assay (ELISA) on recombinant Fc (rFc) and in vitro glycan-engineered Fc. (
Analyses of Fc Glycoform and Admixtures Thereof.

ELISA, enzyme-linked immunosorbent assay.
The calculated contributions to Sambucus nigra lectin (SNA) binding from single- and double-arm sialylated complex Fc N glycans (in vitro glycan engineered) via mass spectrometry (MS) is compared with that obtained from the lectin (SNA) assay.
Potentially, in an engineered biologic product produced in a recombinant setting, the presence of IgGs with varied levels of sialylation and discerning intermediate species would be essential when parsing out glycoforms. This assay would support testing “unknown” glycoforms by obtaining MS signatures for select species—“standards” (rFc, S1Fc, and S2Fc in this case) and admixtures of these standards to provide an entire spectrum of glycoforms, allowing discrimination of all glycoforms embedded with clarity comparable to that of MS. To evaluate the linearity of the lectin assay, appropriate admixtures of rFc, S1Fc, and S2Fc were generated, and the extent of sialylation in the resulting mixtures was measured by lectin-ELISA ( Fig. 2B ). The calibration curve was generated using results obtained by MS analytics along with the calculated and observed levels of sialylation from the SNA lectin assay, and a correlation coefficient of 0.996 was calculated ( Table 2 ).
These results clearly indicated that the lectin-ELISA detected signals from Fc glycans with a level of sensitivity of the assay that was comparable to that of an orthogonal MS route that needed complete cleavage and release of glycans from parent peptide backbone.
Monitoring In-Cell Glycan-Engineered rFc Using Lectin-ELISA Assay
Given that a majority of immunoglobulin-derived biologics are produced in a small number of mammalian cell lines (e.g., Chinese hamster ovary [CHO] the most common; Per.C6, mouse myeloma NS0, baby hamster kidney, and the human embryonic kidney cell line, to name a few others), 26 we wanted to examine the utility of the lectin-ELISA assay in quick assessment of glycoform distribution obtained from cell culture supernatant. To this end, we chose a CHO cell culture system and clonal selection to simulate a biopharmaceutical setting of cell line development.
CHO DG44 cells transfected with Fc and β-(1, 4) galactosyltransferase were cultured in the presence of either hygromycin or puromycin to produce galactosylated rFc (G2Fc). Based on the ECL lectin assay, the hygromycin selection was observed to produce galactosylation levels similar to that of in vitro engineered S1Fc (possessing one terminal galactose) ( Fig. 3A ). As in the case of SNA analysis, the multivalent presentation of ECL (and the consequent steric hindrance) precludes the differentiation between monoantennary and biantennary G2Fc produced from the CHO DG44 cells. To maximize the yield of in-cell engineered S1Fc, the hygromycin culture was scaled up and 21 different clones picked. The ECL lectin assay was used to screen the clones selected for relative levels of asialogalactosylation produced ( Fig. 3B ). An independent evaluation of the S1Fcs produced by these colonies using MS analytics demonstrated >80% similarity in the ranking of the individual clones ( Fig. 3B ). Based on this, a single clone was chosen, purified, and transfected with ST6 to generate a sialylated rFc (rSFc) product. The lectin-ELISA was in sync with MS analytics and yielded identical results for the highest rSFc producer ( Fig. 3C ).
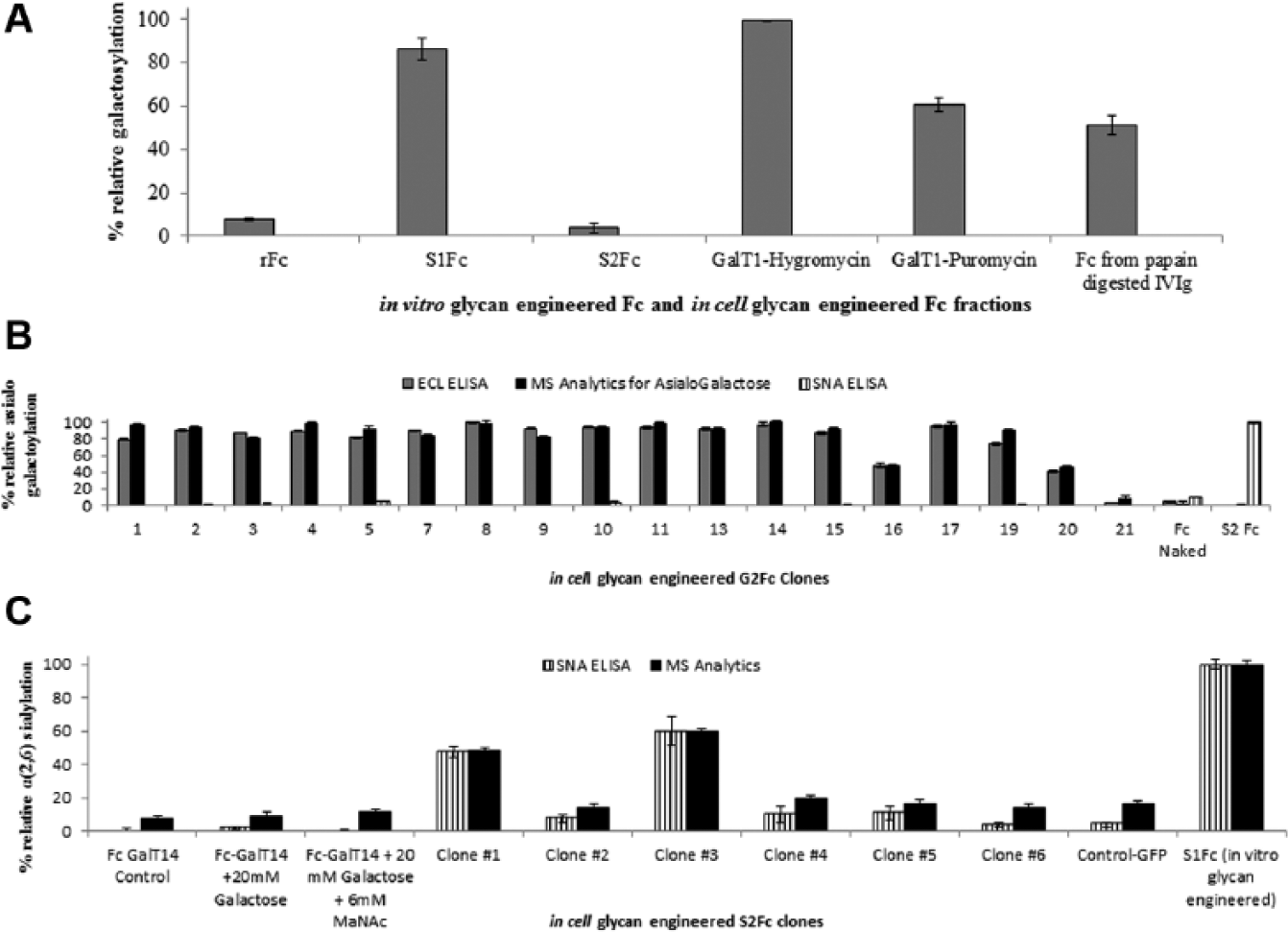
Application of lectin enzyme-linked immunosorbent assay (ELISA) toward screening of in-cell glycan-engineered Fc proteins. (
The similarity in the clonal selection of the highest rSFc producer using the lectin-ELISA and MS thus validates that the plate-based assay can be adapted for monitoring glycoform distribution of therapeutics produced in a cell culture system. Moreover, the assay was performed in quadruplicate and intraplate assay coefficients of variation (CVs) of ~8% and ~4% obtained for the ECL and for SNA ELISAs, respectively.
Quantitative Analysis of In Vitro Glycan-Engineered IVIG Using Lectins and Characterization of IVIG from Commercial Vendors
While both the aforestated applications were on proteins generated in a controlled laboratory setting, the next extension of the assay was on IVIG, a mélange of immunoglobulins and hence glycan epitopes. This biologic was tested at two levels, by generating in-house glycan-engineered versions of commercial IVIG (just as glycan-engineered versions of Fc proteins screened earlier), following which admixtures of these engineered IVIGs were generated and tested. In addition, IVIG samples from several commercial vendors were compared in a concurrent experiment to check the feasibility of this assay for routine screening of glycoforms in a biotech setting.
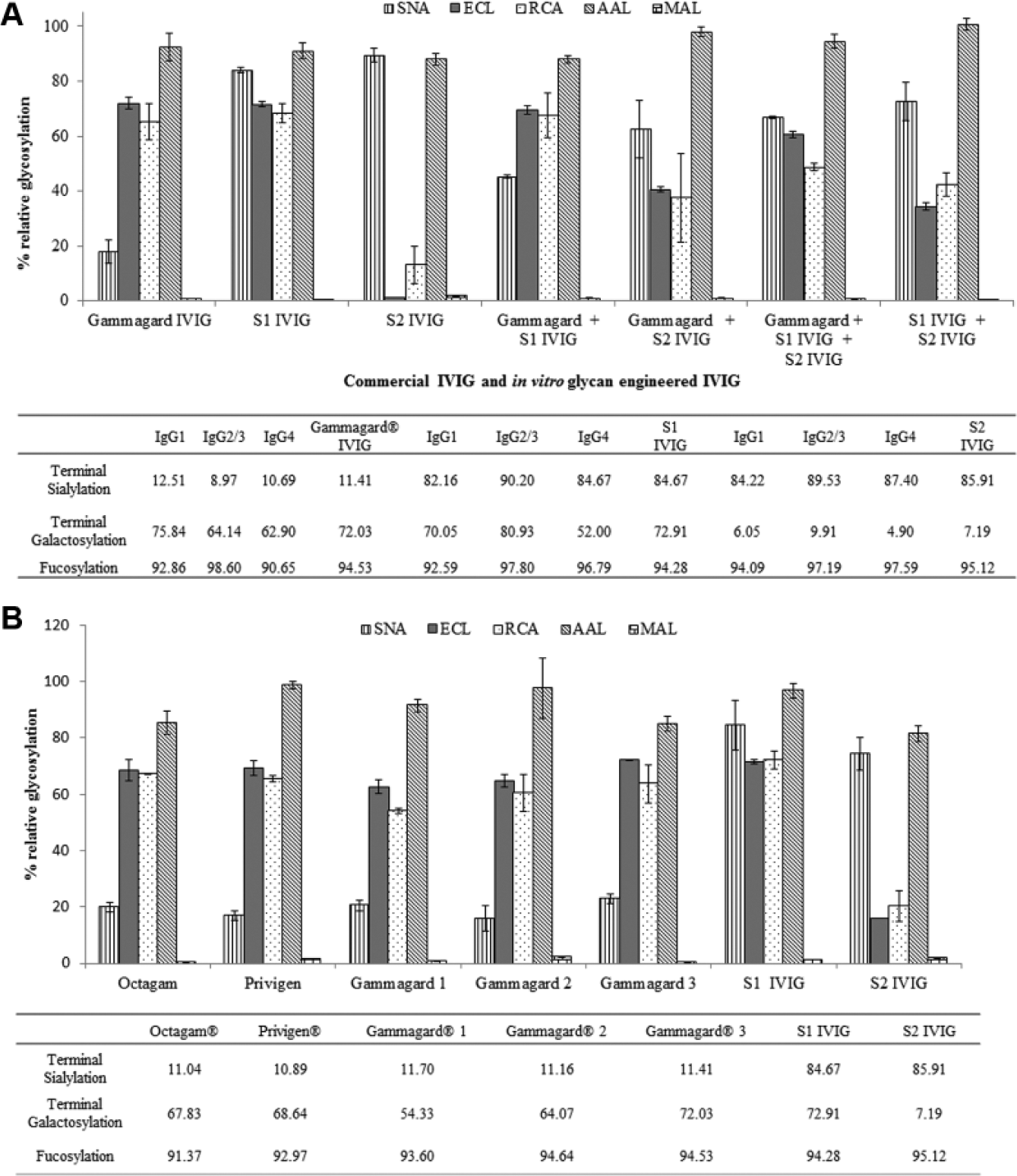
The utility of lectins to serve as highly specific probes to the terminal glycans of interest in a plate assay (from results above) allowed quantitative analysis of commercial IVIG and in vitro engineered IVIG. Specifically, SNA, ECL, RCA, and AAL were used to glycoprofile IVIG (Gammagard, GG) as well its engineered monosialylated (S1 IVIG) or disialylated (S2 IVIG) analogues. Lectin analyses using SNA revealed that GG IVIG was ~20% α-(2, 6) sialylated, while S1 IVIG and S2 IVIG normalized to ~90% sialylation. The difference between mono- and disialylation was revealed by the difference in ECL and RCA binding between S2 IVIG (15%) and S1 IVIG (~75%–80%). Combined together, these results indicate that GG was primarily terminally galactosylated (~70%–75%) while S1 IVIG was monosialylated and S2 IVIG was sialylated on both arms of the biantennary complex glycan. This result was corroborated by MS analytics and by performing lectin analyses on equimolar mixtures of GG, S1 IVIG, and S2 IVIG (
Fig. 4A
). The accuracy of the assay in detecting differences in levels of galactosylation was maintained within 3 to 50 ng IVIG immobilized (
(
Next, we compared batches of commercial-grade IVIG from Octapharma (Octagam), CSL Behring (Privigen), and two batches of Baxter (Gammagard). Analyses using the lectin assay ( Fig. 4B ) revealed no significant differences in their glycan composition, as corroborated by MS analysis ( Fig. 4B ). Specifically, the Octagam and Privigen batches exhibited a glycan signature similar to GG—namely, predominant asialogalactosylation (55%–70%) accompanied with minimal terminal sialylation (~20%). As expected, heavy core fucosylation with levels >90% was observed across all lots tested. SNA and MAL ELISAs indicated the substantial presence of exclusive α-(2, 6) sialylated glycans and negligible α-(2, 3) sialic acids on terminal galactose.
Interplate assay CVs between 8% and 10% were obtained while the intraplate assay CVs ranged between 5% and 9% for the lectin-ELISAs on IVIG (both glycan engineered and commercially sourced), proving the robustness of the ELISA and the reproducible signatures that were generated each time. The bottom panel in Figure 4B details the LC-MS/MS glycosylation analyses across the samples tested and is in sync with the ELISA data. This further substantiated the utility of the lectin-ELISA in screening both commercial and laboratory-generated biologics.
Discussion
Complete glycoprofiling of complex biologics has been made possible by the use of well-established analytical tools.11,28,29 While exhaustive glycan analyses afforded by these techniques are useful, they have limitations. Customized commercial glycan profiling services, although available, are often not accessible to the average researcher.
The present study evaluated a unique strategy to unravel relative levels of carbohydrate compositions, focusing on smaller subsets of glycan structures grouped based on the biological activity they orchestrate. The microtiter plate assay, corroborated by independent MS analytics, takes advantage of the specificity of plant-derived carbohydrate binding proteins to terminal sugar moieties in N-linked Fc glycans such as sialic acid, galactose, mannose, and fucose ( Table 1 ) for the relative quantitation of these functionally relevant carbohydrates. 6 The true merit of the assay developed lies in leveraging the information garnered by the use of pairs of terminal glycan-binding lectins to afford a quantitative interpretation of the same epitopes and as a tool that is an amalgam between LC/MS analytics (used to generate “standards” of proteins with known levels of glycans) and the ELISA for acquisition of details on “unknowns” without the need to run LC/MS on each sample.
Lectin-glycan interactions are weak, with affinity constant (KD) values in the 10−3 to 10−7 M range. 13 Lectin-glycan interactions observed in vivo in the context of bacterial and viral pathogens as well as the galectin family of mammalian lectins repeatedly reinforce multivalency and avidity as core requirements for transmission of downstream effector functions. 30 Consequently, lattice-like arrangements of lectins that allow simultaneous engagement of multiple binding pockets and concentration of lectin-glycan interactions in the in vitro assay systems have shown to improve the apparent KD to picomolar levels,15,31 which is being proposed in our current assay system.
To improve the avidity, multiple biotinylated lectins were linked to neutravidin and used in a concomitant manner to measure sialylation and galactosylation content of recombinant Fcs and engineered Fcs. Admixtures of in vitro glycan-engineered rFcs showed linearity of the assay platform and a high degree of correlation to the MS composition analytics.
Binding of glycan epitopes to each lectin are independent measures, and the panel of lectins is carefully chosen based on the epitopes of interest in the given setting. Comparison of signals from two different lectins with affinities for the same target glycan is not intended. With each lectin, a panel of samples (admixtures) is used to discern differences in signals garnered from the same lectin binding to a different amount of epitopes exposed in each sample, thereby allowing relative quantitation of the epitope in each sample set. Pleiotropic binding to multiple targets is a known facet of lectins, and to get around this caveat, the epitope of interest was quantitated by juxtaposing readouts from a rationally chosen panel of lectins, no two of which have been reported to have the same affinities for the same target.
Furthermore, this ELISA assay can be customized for screening targets of interest, accordingly. Our interest in sialylation, a motif whose role has long been of interest in biologics,12,32,33 led us to select this particular set of lectins. Each lectin in the panel is used to measure signal from a specific epitope, their primary target. Thus, SNA was used to quantitate the levels of α-(2, 6) sialylation, in conjunction with ECL; the latter, for discernment of signal from the penultimate sugar (asialogalactose) and MAL, was chosen for differentiating signals from α-(2, 3) sialylation. AAL and ConA were used to quantify unchanged motifs such as fucosylation and mannosylation and thus served as positive controls.
Recognizing that most therapeutic biologics are produced in recombinant expression systems, the application of the lectin assay was extended to screen expression systems for in-cell galactosylation and sialylation. Again, SNA and ECL were used in an intersecting manner to test levels of sialylation/galactosylation for each of the Fc clones produced. In each case, testing the different recombinant Fc clones yielded a maximal glycosylation ranking, which matched closely with that obtained using MS analysis.
Therapeutic preparations of IVIG obtained from pools of healthy blood donors contain numerous flavors of IgG glycans. The inherent heterogeneity of the product poses a tough challenge for the maintenance of clinical equivalence. There are currently nine IVIG products being produced by six different manufacturers in the US market, and lots from three different manufacturers were selected for testing at random. To assess this complex glycoprotein, the lectin RCA, a lectin with reported specificity for terminal asialogalactose,
22
was added to the lectin panel to further validate the ECL quantitation. IVIG obtained from different suppliers when analyzed using this particular lectin panel showed about 15% to 25% of α-(2, 6) sialylation—an observation that aligns with what has been reported for IVIG composition by other groups and one that we believe predominantly reflects Fc sialylation.27,33 Furthermore, it was demonstrated that the assay was sufficiently sensitive to detect differences in levels of galactosylation and core fucosylation between different batches of commercial IVIG that were independently verified using MS techniques and also described elsewhere20,32,34 (
The profiles of different vendor-sourced IVIGs examined were remarkably similar and likely mirror IgG Fc glycosylation in normal humans given the nature of preparation of this biologic. However, a glycan-engineered version was clearly differentiated and the utility of this assay in picking up nuanced changes between glycoforms noticeably apparent. It is envisioned that the lectin assay can be used for rapid, in-process monitoring of the homogeneity of engineered biologics and ensuring glycoform fidelity in the event of a switch to a modified preparation of IVIG.
In summary, an assay system using apposing reads from multiple lectins specific for terminal and subterminal Fc glycans allowed examination of attributes with a much greater degree of finesse than has been reported using any other commercial lectin-based platforms without any need for sample preprocessing. The role of Fc-conjugated oligosaccharides in defining the biological activity of recombinant therapeutics is now well accepted, with sialic acid taking the lion’s share of interest owing to its pleiotropic cellular effects. At the other end of the spectrum, with most therapeutic antibodies now generated in mammalian hosts, the need for tools to objectively look for differences at the gene, protein, and posttranslational levels has never been greater. Use of plant lectins in the fashion described, opens doors to quickly evaluating homogeneity/heterogeneity of glycoforms of Fc or intact IgGs, with a high level of granularity. Given the vast number of well-characterized plant lectins with a wide range of epitope specificities, it is conceivable to develop such ELISAs for numerous other protein pharmaceuticals in the developmental pipeline.
We recommend that when using this assay system, a panel/library of lectins be chosen with high levels of affinity for terminal and penultimate epitopes of interest, allowing a complete picture to emerge that does not rely on a single lectin for levels of a particular epitope. Continued advances in glycomics leading to the discovery of lectins with novel specificities 35 could potentially be incorporated into the assay described herein, allowing rapid screening of glycan epitopes in high throughput with no additional caveats associated with respect to sample preprocessing while keeping costs minimal.
Footnotes
Acknowledgements
We thank Stan Lee for generation of in-cell glycan-engineered Fc; Amy Medeiros, Naveen Bhatnagar, and Maurice Hains for generation of the in vitro glycan-engineered Fc; Sakshi Bhargava for technical assistance; Carlos Bosques, Thomas Prod’homme, Enrique Arevalo, and Paul Miller for critical review of the manuscript; and Ishan Capila and Amanda Weyers for useful feedback.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by Momenta Pharmaceuticals, Inc.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.