
Abstract
Endothelial cells forming the blood-brain barrier limit drug access into the brain, due to tight junctions, membrane drug transporters, and unique lipid composition. Passive permeability, thought to mediate drug access, is typically tested using porcine whole-brain lipid. However, human endothelial cell lipid composition differs. This investigation evaluated the influence of lipid composition on passive permeability across artificial membranes. Permeability of CNS-active drugs across an immobilized lipid membrane was determined using three lipid models: crude extract from whole pig brain, human brain microvessel lipid, and microvessel lipid plus cholesterol. Lipids were immobilized on polyvinylidene difluoride, forming donor and receiver chambers, in which drug concentrations were measured after 2 h. The log of effective permeability was then calculated using the measured concentrations. Permeability of small, neutral compounds was unaffected by lipid composition. Several structurally diverse drugs were highly permeable in porcine whole-brain lipid but one to two orders of magnitude less permeable across human brain endothelial cell lipid. Inclusion of cholesterol had the greatest influence on bulky amphipathic compounds such as glucuronide conjugates. Lipid composition markedly influences passive permeability. This was most apparent for charged or bulky compounds. These results demonstrate the importance of using species-specific lipid models in passive permeability assays.
Introduction
One major challenge hampering the success of neurotherapeutics is the difficulty in delivering drugs and diagnostic agents to the brain. 1 This is due, in part, to the complexity of the blood-brain barrier (BBB), the site of molecular exchange between the blood and the brain. The BBB is formed by endothelial cells that line cerebral microvessels and consists of both physical and metabolic barriers that work in tandem to maintain tight control of the brain microenvironment. The physical barrier consists of restrictive intercellular tight junctions between endothelial cells, greatly reduced vesicular transport, and a unique suite of proteins that transport nutrients, proteins, hormones, and drugs from the blood to the brain, as well as proteins that mediate the clearance of metabolic waste products and drugs in the opposite direction from the brain to the blood. The metabolic barrier, consisting of phase I and phase II drug-metabolizing enzymes, functions to prevent lipophilic compounds from crossing the BBB by increasing their hydrophilicity.2–4 In order for a drug-like molecule to enter into the brain, it must pass through the plasma membrane of endothelial cells, transcellularly rather than paracellularly. Thus, vascular capillary endothelial cells are the “gatekeepers” of the brain, restricting the entry of toxic as well as potentially therapeutic compounds.
Many potential drugs have failed to succeed therapeutically, however, because they are too bulky or lack the necessary lipophilicity to cross the BBB and thus are unable to reach their therapeutic target. Therefore, it is important to understand endothelial cell membrane permeability potential when developing drugs for central nervous system (CNS) diseases. Several in vitro and computational models have been used as putatively physiologically relevant models in the drug discovery process to predict the BBB penetration potential of drug candidates. Computational models of drug permeability are typically based on molecular descriptors and range in complexity from simple multiple linear regression to machine-learning methods and have varying degrees of prediction performance.5–7 Similarly, in vitro BBB models have a range of complexity, throughput, and performance. The simplest models use lipid immobilized on a bead or solid support.8–10 In contrast, more complex, cell-based models use brain microvessel endothelial cells cultured either alone or co-cultured with astrocytes and/or pericytes. 11
The parallel artificial membrane permeability assay (PAMPA) is an immobilized lipid model for measuring passive membrane permeability of small molecules. It was originally developed as a simple method for prediction of transcellular drug absorption and used a synthetic lipid immobilized on a filter support to mimic passive permeability across a cellular membrane. 12 The assay has since been modified to mimic specific BBB, 8 intestinal, 13 and skin 14 permeability barriers. The BBB-PAMPA assay as originally developed uses a lipid extract from whole pig brains (polar brain lipids). 8 The assay was able to discriminate between BBB permeable or nonpermeable compounds. 8 However, when the assay was used in a separate study to predict in vivo brain penetration, the correlation between the effective permeability (Peff) for the porcine polar brain lipids model compared with the log BB (logarithm of the brain to plasma concentration ratio) after oral administration in rats of 14 structurally diverse compounds was less than that determined using the synthetic lipid dioleoylphosphatidylcholine alone. 15 This demonstrates the importance of using a model membrane that closely resembles the tissue or cells of interest, to ensure that the behavior seen with the lipid model membrane reflects the behavior of cells in vitro and/or in vivo. 16
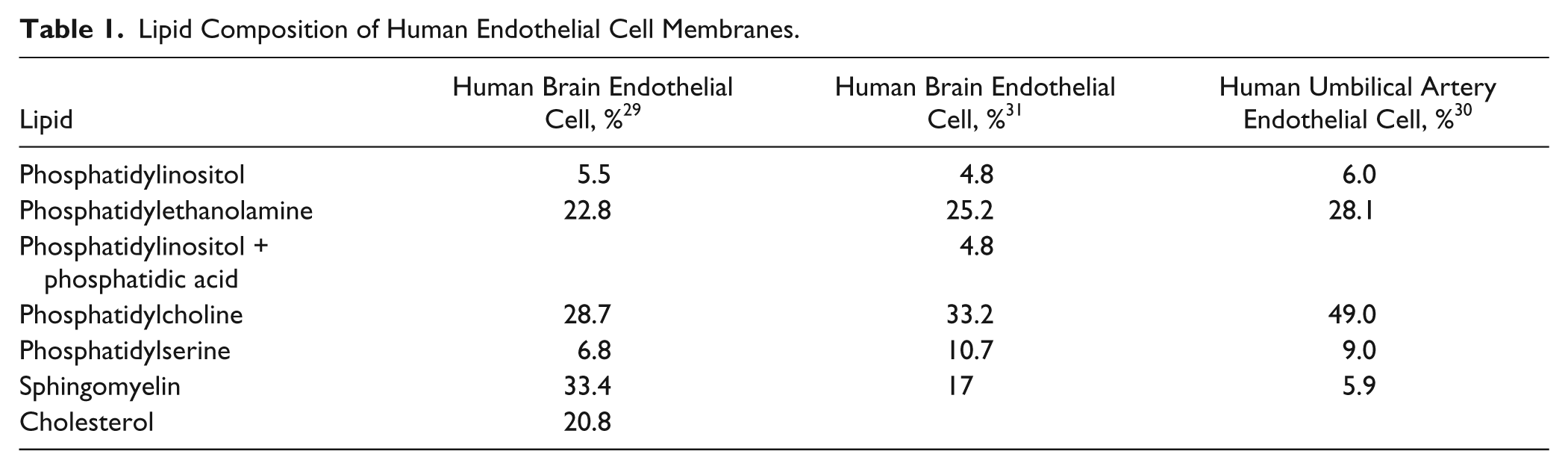
The lipid composition of human BBB endothelial cell membranes ( Table 1 ) differs from that of porcine polar whole-brain lipid (PBL), traditionally used for PAMPA-BBB assays ( Table 2 ). Such differences could potentially result in model misspecification when using pig brain lipid to predict human CNS drug disposition.
Lipid Composition of Human Endothelial Cell Membranes.
Lipid Composition of PAMPA Models. a
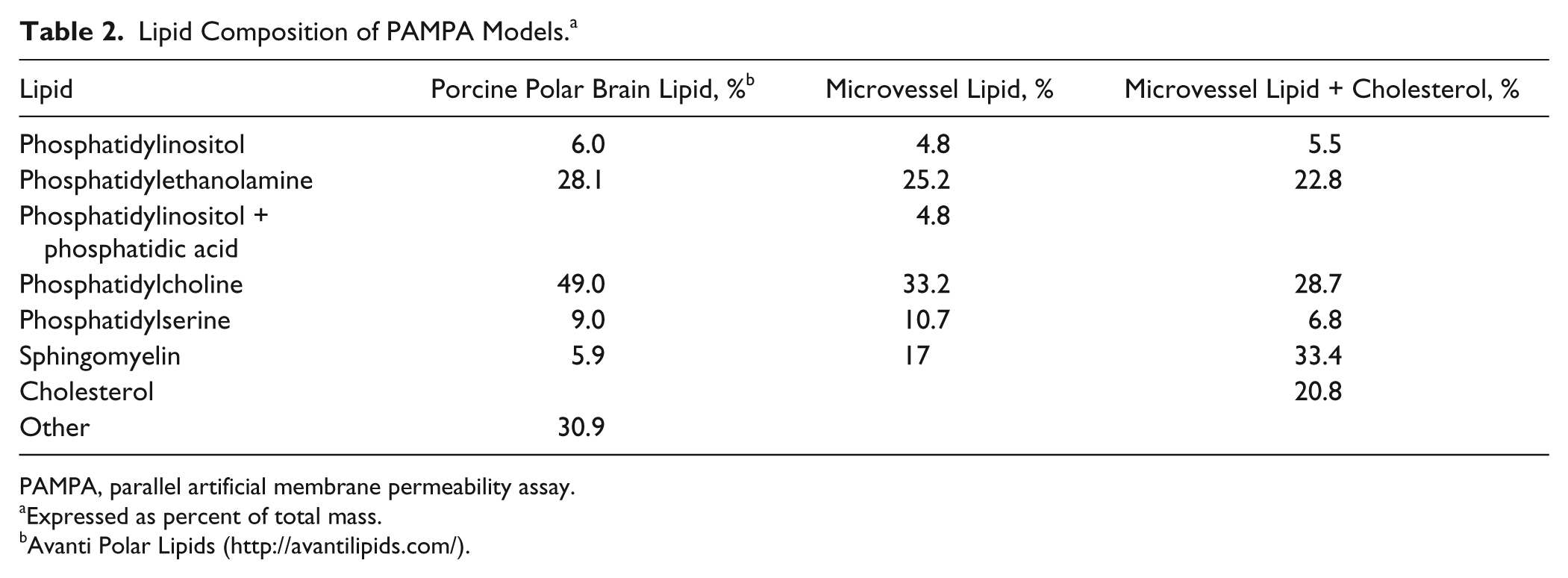
PAMPA, parallel artificial membrane permeability assay.
Expressed as percent of total mass.
Avanti Polar Lipids (http://avantilipids.com/).
The objective of the present study was to compare the effect of membrane lipid composition on the passive permeability of 18 compounds and metabolites active in the CNS using the PAMPA assay. In addition to traditional porcine polar whole-brain lipid, one model system was designed to reflect the lipid composition of human brain endothelial cells (microvessel lipid model, MVL) and the other model to reflect human brain endothelial cell lipid and cholesterol content (microvessel lipid plus cholesterol, MVLC). The hypothesis was that lipid and cholesterol composition in model lipid systems would influence passive permeability.
Materials and Methods
Materials
Multiscreen polyvinylidene difluoride (PVDF) 96-well plates and Transporter Receiver Plates were obtained from Millipore (Billerica, MA). Lipids were obtained from Avanti Polar Lipids (Alabaster, AL) with the exception of sphingomyelin, cholesterol, phosphatidylethanolamine, and phosphatidylcholine, which were obtained from Sigma-Aldrich (St. Louis, MO). Verapamil, loperamide, theophylline, (±) methadone HCl, fentanyl citrate, codeine, diazepam, hydromorphone HCl, alprazolam, caffeine, and atenolol were also obtained from Sigma-Aldrich. Alfentanil, norbuprenorphine, buprenorphine, buprenorphine-3-β-D-glucuronide, morphine-3-glucuronide, morphine-6-glucuronide, and oxycodone were obtained from the National Institute on Drug Abuse Research Resources Drug Supply Program. Oxazepam was obtained from Hoffman-La Roche. Sufentanil and dihydromorphine were obtained from Cerilliant (Round Rock, TX) and morphine sulfate from Mallinckrodt (St. Louis, MO). Tramadol and O-desmethyltramadol were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Norbuprenorphine-3-β-D-glucuronide was synthesized from norbuprenorphine using dog liver microsomes. 17 Hank’s buffered salt solution (HBSS), pH 7.4, was obtained from MediaTech (Manassas, VA).
Methods
With the exception of the polar brain lipid, which was solubilized as received from the manufacturer, the individual components for each lipid preparation were solubilized in anhydrous dodecane (Sigma-Aldrich). All lipid preparations had a total lipid concentration of 20 mg/mL and were prepared fresh prior to each assay. Each of the test compounds was diluted to 2 mM in DMSO (Sigma-Aldrich) and further diluted prior to testing to 2 µM in HBSS, pH 7.4. The assay was performed using a 96-well Multiscreen-IP PAMPA plate (Millipore). Each well was prepared by first adding 4 µL of the lipid suspension to the PVDF membrane of the filter plate, as described previously. 8 Immediately following the addition of the lipid to the membrane, the donor (upper) chamber was filled with HBSS containing the compound to be tested (200 µL). HBSS was also added to the receiver plate (300 µL), and the filter and receiver plates were then assembled and incubated for 2 h at room temperature in a moistened sealed bag to prevent evaporation. At the end of the incubation period, 100 µL was removed from the receiver plate, and the concentration was determined by HPLC–tandem mass spectrometry (MS/MS). An additional equilibrium plate was used, representing a theoretical, partition-free sample. The equilibrium plate was prepared by adding 300 µL HBSS and 200 µL HBSS plus 2 µM drug to a 96-well deep-well plate, mixing and transferring 100 µL of the diluted drug to a new plate for analysis. Analysis of each compound was performed in triplicate on 3 different days.
Analytes were quantified by HPLC-MS/MS. Samples were diluted 1:1 with purified water (MilliQ; Millipore) containing 0.1 µg/mL norfentanyl d5 (Cerilliant) as the internal standard and mixed thoroughly. Liquid chromatography–MS/MS analysis was performed on an ultra-fast liquid chromatography system from Shimadzu Scientific Instruments (Columbia, MD) consisting of a CMB-20A system controller, two LC-20ADXR pumps, a DGU-20A3 degasser, a SIL-20AC autosampler, and a CTO-20A column oven. An external switching valve was installed between the chromatography system and the mass spectrometer. The chromatography system was coupled to an API 4000 QTrap LC-MS/MS linear ion trap triple quadrupole tandem mass spectrometer from Applied Biosystems/MDS Sciex (Foster City, CA). The mass spectrometer was equipped with a turbo ion spray ionization source operating in a positive ionization mode. Chromatographic separation was performed on a Sunfire C18 HPLC column (50 × 2.1 mm, 3.5 µm) (Waters Corp., Milford, MA). The injection volume was 10 µL, the autosampler temperature was 15 °C, and the oven temperature was 40 °C. The HPLC mobile phase (0.3 mL/min) was (A) 4.5 mM ammonium acetate in water (pH 4.5) and (B) 4.5 mM ammonium acetate in acetonitrile. The gradient program was as follows: 0 to 0.5 min 5% B, increased linearly to 95% B at 1 min, held at 95% B until 2.5 min, linearly decreased to 5% B over 0.01 min, and reequilibrated at 5% B until 4 min. The mass spectrometer was operated in positive ion mode with an ion spray voltage of 5500 V. The source temperature was 450 °C, entrance potential was 10 V for all compounds, and curtain gas, ion source gas 1, ion source gas 2, and collision gas (all ultra-high-purity nitrogen) were 20 psig, 30 psig, 40 psig, and medium setting, respectively. Both Q1 and Q3 quadrupoles were optimized to unit mass resolution, and the mass spectrometer conditions were optimized for each analyte. Multiple-reaction monitoring transitions for each analyte and internal standard, as well as other mass spectrometric parameters, are provided in
Canonical SMILES strings for each compound were obtained from the PubChem Web site (pubchem.ncbi.nlm.nih.gov/), and molecular descriptors ( Table 3 ) were calculated using ACD/I-Lab (Advanced Chemistry Development, Toronto, ON, Canada).
Structural Features and Molecular Descriptors. a
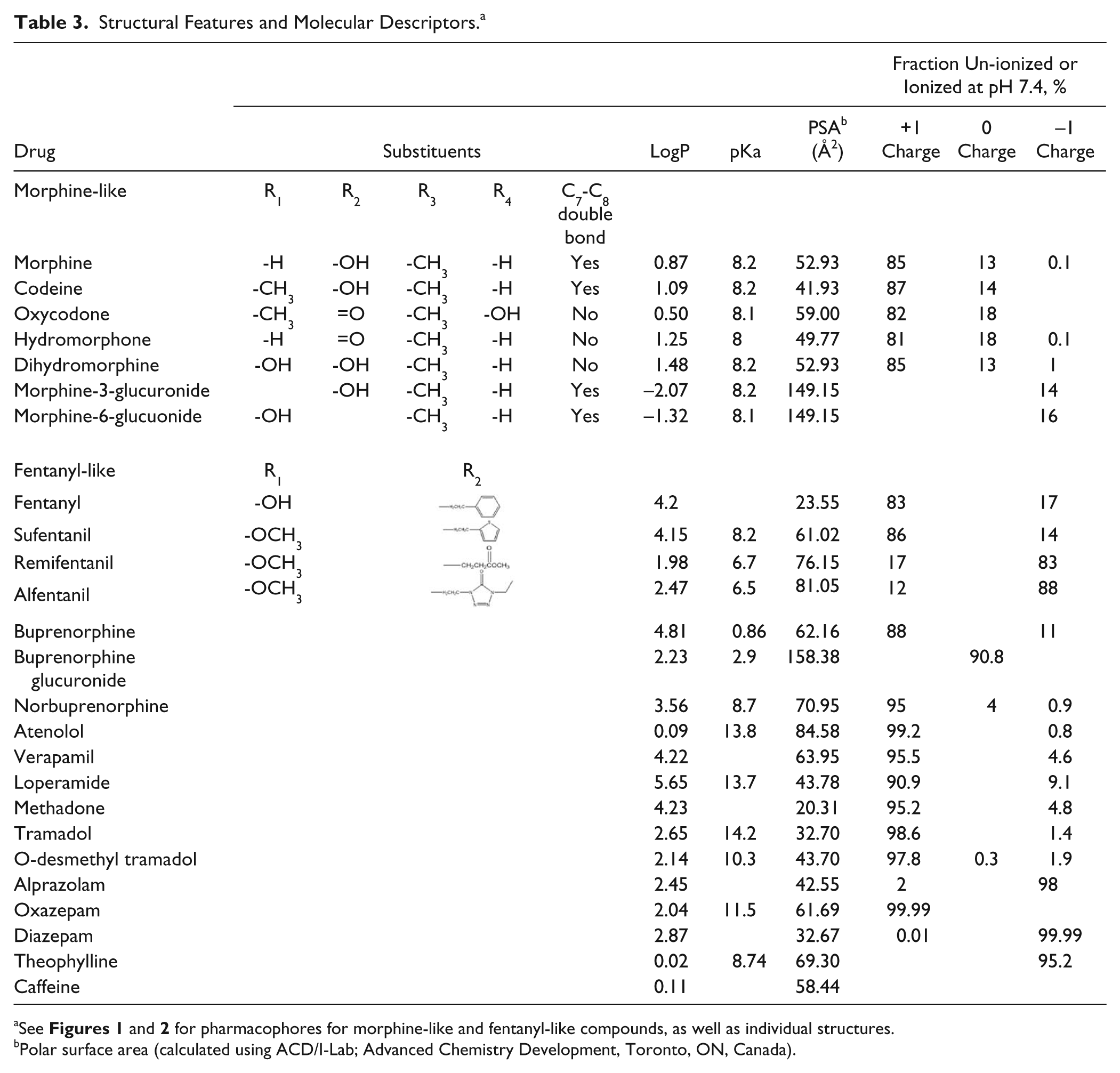
See Figures 1 and 2 for pharmacophores for morphine-like and fentanyl-like compounds, as well as individual structures.
Polar surface area (calculated using ACD/I-Lab; Advanced Chemistry Development, Toronto, ON, Canada).
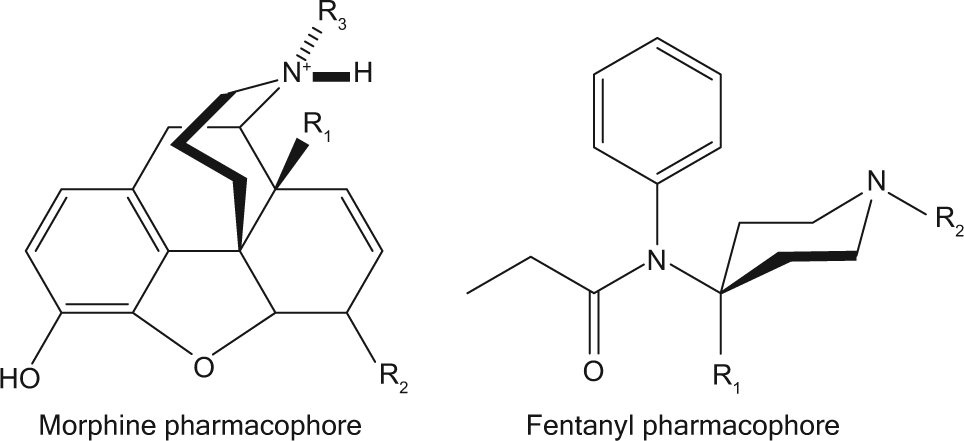
Morphine and fentanyl pharmacophores. Specific compounds and substituents are described in Table 3 .
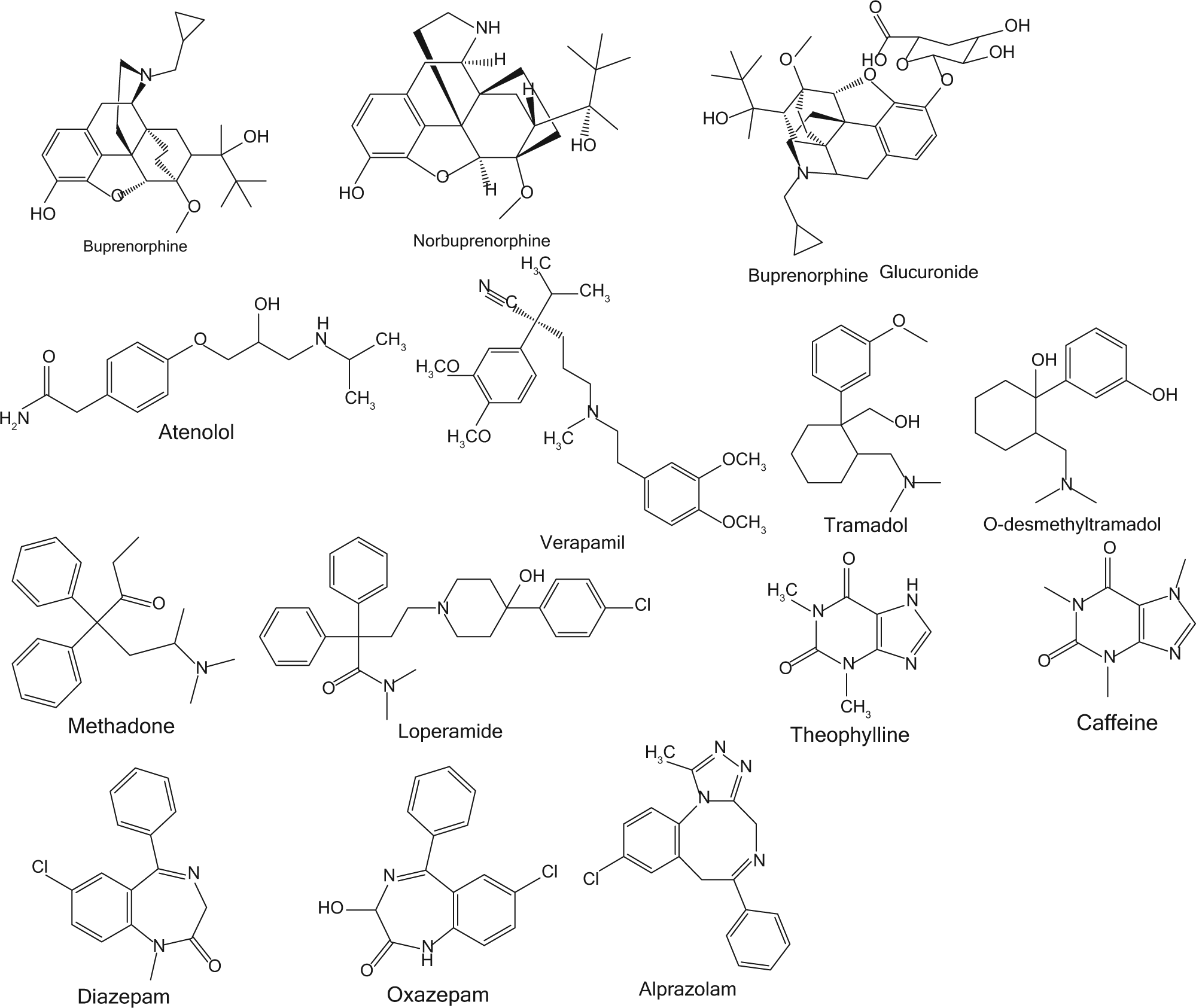
Structures of individual compounds.
Calculations
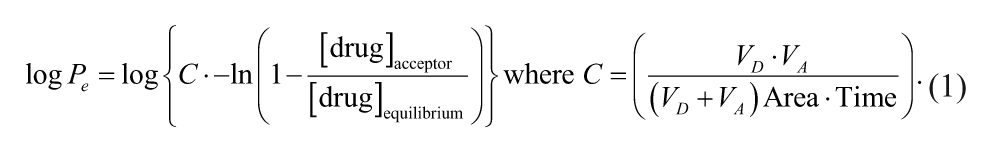
The log of the effective permeability was calculated using equation (1): 18
Each data point represents the mean and standard deviation of triplicate determinations performed on the same day.
Results and Discussion
The polar brain lipid-based PAMPA assay was developed to measure the passive permeability of compounds in a brain lipid environment. 8 However, the lipid typically used for this assay is from pig brain, rather than human brain, and, more specifically, is extracted from the whole pig brain, rather than from brain microvessel endothelial cells. These considerations confer substantial differences in the abundance of several component phospholipids in the brain ( Table 1 ). Therefore, the purpose of this study was to determine the effect of using a more physiologically relevant phospholipid composition (human and microvessel endothelial lipid) on the effective permeability of CNS active drugs. Representative compounds from several drug classes were selected, including morphine- and fentanyl-like opioids, other opioids, benzodiazepines, and drugs commonly used to benchmark brain permeability.
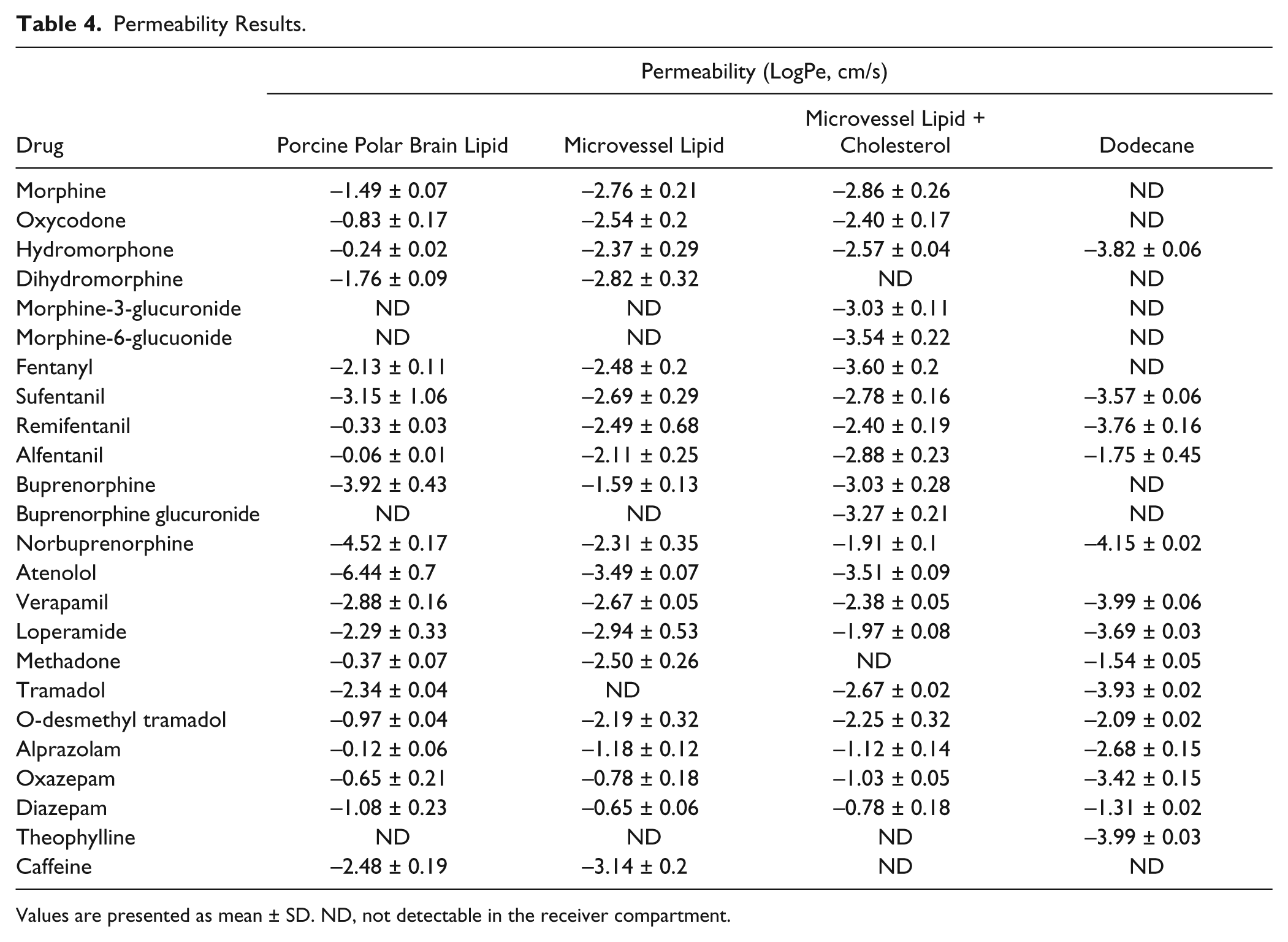
Control experiments were performed to determine if permeability was due to interactions with the phospholipids/sphingomyelin/cholesterol or with the nonpolar solvent dodecane. Thus, the PAMPA assay was performed using the lipid solvent dodecane alone, without lipid, between the donor and receiver chambers ( Table 4 ). Comparison of results to permeability with the lipid models reflects the role of the lipid membrane per se. With dodecane alone, morphine, oxycodone, dihydromorphine, fentanyl, buprenorphine, and the glucuronide conjugates of both morphine and buprenorphine were unable to cross the barrier, indicating the obligate necessity of lipid interaction for membrane permeability. The effective permeability in the dodecane-only model for hydromorphone, norbuprenorphine, tramadol, alprazolam, oxazepam, and diazepam was generally greater in the three lipid-containing models than with dodecane alone. For the remaining compounds, alfentanil, sufentanil, and O-desmethyl tramadol, the logPe values were equivalent in the presence or absence of lipid. The differences between these three groups of compounds are likely due to two separate properties. Compounds that were unable to cross the dodecane barrier, such as the glucuronide conjugates of buprenorphine and morphine, are likely too charged. These compounds have 5 H-donor and 10 (or 11) H-acceptor sites. In contrast, for compounds such as fentanyl and buprenorphine, which are highly lipophilic (as measured by their logP values), the rate-limiting step may be rapid uptake into the lipid membrane but slow desorption off the membrane surface into the receiving buffer.
Permeability Results.
Values are presented as mean ± SD. ND, not detectable in the receiver compartment.
Subsequent evaluations compared the influence of membrane lipid composition in the permeability of the representative compounds ( Table 4 ). Oxazepam, diazepam, and alprazolam had high permeability in all three lipid models. All three compounds are uncharged (>98%) at pH 7.4, and they may cross the membrane without the need for hydrogen bonding with the lipids. Another possibility is that these compounds cross the artificial membrane through paramembrane water channels that are thought to exist. 19 However, the lipid compositions in the three MVL and MVLC models did influence the movement of these benzodiazepines across the lipid membrane, with differences between the human and porcine lipids, as well as between the human lipid with and without cholesterol. This may be affected by tighter packing of the phospholipid/sphingomyelin molecules in the MVL and MVLC models. In contrast, compounds such as alfentanil, remifentanil, hydromorphone, and norbuprenorphine were highly permeable in the PBL model but one to two orders of magnitude less permeable in either the MVLC or the MVL models; in addition, the absence or presence of cholesterol in the MVL model had little influence on their permeability. These results are not readily explained by molecular charge, polar surface area, or logP.
For most compounds, the PBL model appears to be less restrictive than either of the models using species-relevant lipid compositions. The only exceptions were sufentanil, diazepam, and the glucuronide conjugates, which had greater permeability in both the human MVLC and MVL models than the PBL model. One possible explanation for the generally greater permeability of the porcine polar brain lipid monolayer, compared with human brain lipid, may be the lower content of sphingomyelin. Since, however, the lipid content of the PBL model is not fully characterized, with approximately 30% of the lipid undefined, additional explanations may exist.
For the two cholesterol-free models, porcine PBL and human MVL, the greatest difference in permeability between models appears to be for those compounds (i.e., remifentanil, alfentanil) that have extensive hydrogen bond acceptor potential but are primarily (>80%) uncharged at pH 7.4. In contrast, opioids that are structurally similar to remifentanil and alfentanil (i.e., fentanyl and sufentanil) but have a limited capacity to form hydrogen bonds and are charged at pH 7.4 have logPe values that are much more similar in the porcine and human lipid models. These differences may be reflective of lower sphingomyelin content in the PBL compared with MVL models. Sphingomyelins, because of their amide bond at position 2 and hydroxyl at position 3 of the sphingoid base, are capable of acting as both a hydrogen donor and acceptor. In contrast, phosphatidylcholine can act only as a hydrogen acceptor due to the two ester carbonyl groups. 20
The presence of cholesterol in the artificial human membrane appears to have the greatest influence on bulky amphipathic compounds such as the glucuronide conjugates of buprenorphine and morphine. These compounds have extensive hydrogen bonding potential and primarily exist as a zwitterion (±1) at pH 7.4. However, the presence of cholesterol also influences the permeability of highly lipophilic compounds such as fentanyl. Cholesterol has been shown to interact with sphingomylin, forming “rafts” that influence phospholipid packing, membrane fluidity, and the pharmacological effect of some drugs.21–24 The lower permeability of fentanyl in the cholesterol- containing MVL model is surprising and may represent a high level of lipid retention.
The unique barrier properties of brain microvessel endothelial cells function to maintain a defined environment for the CNS by allowing specific nutrients to enter, eliminating waste products, and excluding substances potentially toxic to neuronal tissue. Among those substances with variable entrée are drugs for treating neurological disorders and pain. The relative roles of carrier-mediated uptake and passive diffusion in brain access and bioavailability of drugs remain under active discussion.25–27 The standard model for assessing passive permeability is the PAMPA assay. PAMPA studies designed to model the blood-brain permeability of therapeutic drugs and metabolites, as well as predict the permeability of compounds in development, are increasingly evaluating larger families of compounds, yet continue to use porcine whole-brain polar lipid.15,19,28 Nevertheless, two considerations prevail—specifically, the relevance of whole-brain lipid and species differences.
The lipid composition of brain microvessel endothelial cells, which are the primary determinant of the BBB, is unique, such that the amount of sphingomyelin is approximately 3-fold higher, and phosphatidylcholine content is decreased by approximately one-third, compared with other endothelial cells.29,30 The relative abundance of these two structurally similar but functionally dissimilar phospholipids has the potential to alter phospholipid packing within the membranes and phospholipid-cholesterol interactions. These, in turn, can influence the ability of compounds to interact with the lipids of the BBB, as well as their ability to form hydrogen bonds, thereby affecting the permeability of (potential) therapeutic compounds.
The lipid composition of porcine whole-brain extracts differs from human brain endothelial cell lipid. The permeability of drugs in rodent in situ brain perfusion studies correlates well with PAMPA studies based on porcine brain lipid extract.15,19,28 However the ability of porcine whole-brain PAMPA assays to predict human brain drug access in vivo remains unknown.
These considerations are validated by the results and conclusions of the present investigation, which demonstrate that in a small series of drugs, there are significant differences in the passive permeability of various compounds, between porcine and human lipid, as well as between the porcine whole-brain lipid model and those reflecting human endothelial cell lipid composition. This highlights the need to reevaluate the current BBB-PAMPA assay using porcine polar brain lipid, to better model human brain biodistribution, and in order to identify compounds having the greatest likelihood of reaching their brain targets.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: National Institutes of Health grants R01-GM63674, R01-DA14211, R01-DA025931 and K24-DA00417 (to EDK).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.