
Abstract
Recent advances in stem cell technology have enabled large-scale production of human cells such as cardiomyocytes, hepatocytes, and neurons for evaluation of pharmacologic effect and toxicity of drug candidates. The assessment of compound efficacy and toxicity using human cells should lower the high clinical attrition rates of drug candidates by reducing the impact of species differences on drug efficacy and toxicity from animal studies. Methyl-β-cyclodextrin (MBCD) has been shown to reduce lysosomal cholesterol accumulation in skin fibroblasts derived from patients with Niemann Pick type C disease and in the NPC1-/- mouse model. However, the compound has never been tested in human differentiated neurons. We have determined the cholesterol reduction effect of MBCD in neurons differentiated from human neural stem cells (NSCs) and commercially available astrocytes. The use of NSCs for producing differentiated neurons in large quantities can significantly reduce the production time and enhance the reproducibility of screening results. The EC50 values of MBCD on cholesterol reduction in human neurons and astrocytes were 66.9 and 110.7 µM, respectively. The results indicate that human neurons differentiated from the NSCs and human astrocytes are useful tools for evaluating pharmacologic activity and toxicity of drug candidates to predict their clinical efficacy.
Keywords
Cell-based assays are commonly used in early drug discovery for high-throughput screening (HTS) to identify lead compounds. They are also extensively employed in secondary and tertiary screens to confirm the activities of compounds that are initially identified from HTS using biochemical assays.1, 2 Currently, cells primarily used for compound screens are either immortalized cell lines or primary cells freshly isolated from tissues. Both primary and immortalized cells can be derived from various species, including rodents (hamster, mouse, and rat) and humans, as well as from distinct tissues such as brain, heart, liver, muscle, and skin. Although the compound activity data obtained from animal cells are useful for guidance of drug dosing in animal model studies, compound potencies determined from human cells are expected to better predict the drug efficacy and toxicity in human clinical trials. Immortalized human cell lines are commonly cancerous and/or have accumulated significant genetic mutations. Hence, these do not provide a valid model of the biological and pharmacological effects of a given compound. 3 Human blood cells and skin fibroblasts are generally accessible; however, most primary cell types are not available in large quantities needed for drug screens or are prohibitively expensive. The recent availability of differentiated neurons derived from human-induced pluripotent stem cells (iPSCs) provides a new paradigm for the use of human cells to determine the efficacy and potential cytotoxicity of drug candidates.4,5
Niemann Pick type C (NPC) disease is caused by mutations in either one of the two genes encoding the lysosomal cholesterol binding proteins NPC1 or NPC2.6,7 NPC disease is characterized by lysosomal accumulation of unesterified cholesterol and other lipids in many cell types. Lysosomal accumulation results from the impaired transport of unesterified cholesterols from the late endosomes and/or lysosomes to the plasma membrane, trans-Golgi network, and endoplasmic reticulum. The clinical manifestation of NPC disease includes progressive neurodegeneration and is often accompanied by hepatosplenomegaly. The onset of symptoms generally occurs in early childhood, leading to death within a decade. Currently, there is no cure or effective treatment for NPC disease.
Cholesterol accumulation in late endosomes and lysosomes is a direct disease manifestation that can be used as a phenotypic screen to identify cholesterol-reducing compounds. 8 Skin fibroblasts derived from NPC patients exhibit significant lysosomal cholesterol accumulation and thus can serve as a cell-based disease model for compound screens. A recent HTS using the NPC patient fibroblasts led to identification of several compounds that significantly reduced cholesterol accumulation in late endosomes and lysosomes. 8 However, although neuronal degeneration is a major cause of death in NPC patients, cholesterol accumulation in skin fibroblasts does not directly contribute to the disease pathogenesis. Human neurons would thus be more desirable as a cell-based model for compound screens to evaluate activity and neuronal toxicity of drug candidates. Here, we report the application of human neurons differentiated from iPSC-derived neural stem cells (NSCs) as a model system to evaluate the pharmacologic activity and cytotoxicity of methyl-β-cyclodextrin (MBCD). Using a biochemical cholesterol assay, the potency of MBCD has been measured in these cells and compared with skin fibroblasts, astrocytes, and a neuroblastoma cell line. We propose that the human differentiated neurons can be employed as a new screen paradigm for evaluation of compound activity and toxicity.
Materials and Methods
Amplex-red cholesterol assay kit (catalog No. A12216) was purchased from Invitrogen (Carlsbad, CA). ATPlite cell viability assay kit (catalog No. 6016941) was obtained from PerkinElmer (Waltham, MA). DRAQ5 nuclear dye was obtained from Cell Signaling (Danvers, MA). Primary antibodies were purchased from GeneTex (Irvine, CA) and Cell Signaling Technology. MBCD was obtained from MP Biomedicals (Solon, OH). DyLight conjugated fluorescent rabbit and mouse antibodies were purchased from Epitomics (Burlingame, CA). The specific antibodies for Oct4, Sox1, Nanog, and Neurofilament (NF) were obtained from Cell Signaling Technology. The antibodies for Nestin, microtubule-associated protein 2 (MAP2), and β-tubulin class III (TUJ-1) were purchased from Genetex (Irvine, CA). The 96-well black clear-bottom plates (catalog No. 655090) were purchased from Greiner Bio-One (Monroe, NC).
Cells and Cell Culture
Human skin fibroblasts
The wild-type (GM05659) and NPC1 (GM03123) human skin fibroblast cell lines were purchased from the Coriell Cell Repository (Camden, NJ). The human primary skin fibroblasts are derived directly from skin biopsies and have not been transformed, although they can be passaged multiple times. The fibroblasts were cultured in DMEM medium (Invitrogen, catalog No. 11995-040) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin in a humidified incubator with 5% CO2 at 37 °C. For imaging experiments, cells were seeded at 1500 cells/well in 100 µL medium in the black/clear-bottom, tissue culture–treated 96-well plates and cultured overnight. Cells were seeded at 3000 cells/well in 100 µL medium in 96-well plates and cultured overnight before the LysoTracker fluorescence intensity experiment.
Neurons differentiated from human NSCs
Prior to cell plating, the dishes were coated with CELLstart from Invitrogen at 1:100 dilution in Dulbecco’s phosphate-buffered saline (DPBS) containing calcium and magnesium. Plates were rinsed with DPBS before cell seeding. The following method was used to coat plates for differentiation: (1) the surface of plates were coated with poly-L-ornithine working solution at 20 µg/mL in distilled water (14 mL for T-75, 7 mL for T-25, 3.5 mL for 60 mm dish, 2 mL for 35 mm dish) and incubated overnight at room temperature, (2) the poly-L-ornithine-coated plates were washed three times with distilled water and then coated with laminin working solution at 10 μg/mL in D-PBS without calcium or magnesium (14 mL for T-75, 7 mL for T-25, 3.5 mL for 60 mm dish, 2 mL for 35 mm dish), (3) the laminin-coated plates were incubated for 3 h at 37 °C before use or stored wrapped in parafilm for a week at 4 °C.
NSCs, originally derived from normal human iPSCs, were obtained from xCell Science (Novato, CA) and expanded in CellStart-coated dishes. NSCs were cultured in Neurobasal media supplemented with B27, glutamine, nonessential amino acids, and 20 ng/mL fibroblast growth factor (bFGF). To initiate differentiation of NSCs, cells were plated on poly-L-ornithine and laminin-coated 96-well plates in medium supplemented with 10 ng/mL brain-derived neurotrophic factor (BDNF) and 10 ng/mL glial-derived neurotrophic factor (GDNF). Cells were grown in differentiation medium for 10 to 12 days before proceeding to compound treatment and immunocytochemistry.
Astrocytes
Human astrocytes (catalog No. K1884, Invitrogen) are derived from normal human brain tissue and were cultured as per instructions provided by the manufacturer. Briefly, a single vial of astrocytes was immediately suspended in complete astrocyte medium (DMEM, 1X N-2, 10% FBS) and seeded into a single well of a six-well tissue culture–treated plate (catalog No. 353046, BD Biosciences) that had been coated with Geltrex (catalog No. 12760, Invitrogen). Cells were allowed to adhere for at least 24 h at 37 °C with 95% humidity and 5% CO2. The medium was exchanged every other day.
SH-SY5Y neuroblastoma cells
SH-SY5Y cell line (catalog No. CRL-2266) was obtained from ATCC. The base medium for this cell line is a 1:1 mixture of ATCC-formulated Eagle’s Minimum Essential Medium (catalog No. 30-2003) and F12 Medium. To make the complete growth medium, FBS was added to a final concentration of 10% to the base medium.
Neuronal and Stem Cell Markers
Immunofluorescent staining of differentiated neurons in 96-well plates
Cells were fixed in 100 µL 4% paraformaldehyde for 10 to 20 min at room temperature and then rinsed briefly with PBS before permeabilization with 0.3% Triton X-100 for 15 min. Nonspecific protein-protein interactions were blocked with 5% donkey serum and 1% bovine serum albumin. Differentiated neurons were incubated with primary antibody overnight at 4 °C, unbound antibody was removed by phosphate-buffered saline (PBS) washing, and the bound antibody was detected using a goat anti-rabbit secondary antibody conjugated with DyLight 594 (catalog No. 3066-1, 1:200 dilution; Epitomics, Burlingame, CA). After incubation, cells were rinsed twice with PBS and nuclei were stained with Draq5 for 20 min and immediately imaged using an InCell2000 Analyzer (GE Healthcare, Waukesha, WA) with 20× objective lens and filter sets for Cy5 and Texas Red. All secondary antibodies were tested for cross-reactivity and nonspecific immunoreactivity.
Immunofluorescence staining of astrocytes in 96-well plates
Human astrocytes (Invitrogen) were seeded into wells of a BD PureCoat black, clear-bottom amine-coated plate (BD Biosciences, No. 354717) at a density of 6400 cells/well in 100 µL of Complete Astrocyte Media (6.4 × 104 cells/mL). After incubation for 6 days at 37 °C with 95% humidity and 5% CO2, immunofluorescence staining was performed as described above, with minor changes. Astrocytes were incubated in 100 µL of primary antibody solution (1:400 dilution of rabbit anti-glial fibrillary acidic protein [GFAP], Sigma No. G9269) and allowed to incubate overnight at 4 °C, and bound antibody was detected with a goat anti-rabbit secondary antibody conjugated with Dylight 594 (Epitomics). Nuclei were stained with Hoechst 33342. Cells were imaged using the InCell2000 Analyzer with 20× objective lens and the filter sets for DAPI and Texas Red.
Amplex-Red Cholesterol Assay
Amplex-Red Cholesterol Assay Kit (Invitrogen) was used to measure the total cholesterol in patient cells. The unesterified cholesterol was determined using the same kit but without acid lipase. Esterified cholesterol was determined as the difference between the total and unesterified cholesterol values. The cells were seeded in black, tissue culture–treated 96-well plates at a density of 2500 cells/well in 100 µL medium and cultured for 24 h. The medium in the assay plates was removed and replaced with 100 µL/well of compound dissolved in medium. After incubation for 1 or 3 days, the cells were washed twice with HBSS, and 100 µL/well of reagent mixture from the cholesterol assay kit was added. The resulting fluorescence intensity after 1 hour incubation at 37 °C was measured with an excitation wavelength of 560 (±10) and emission wavelength of 590 (±10) in a ViewLux plate reader (PerkinElmer).
ATP Content Assay
The ATP content assay kit (ATPLite, PerkinElmer) was used to monitor compound cytotoxicity. Cells were seeded in white solid-bottom 96-well plates and treated with compounds as described above for the cholesterol assay. After incubation with compound for 1 or 3 days, 100 µL/well of reagent mixture (prepared according to the manufacturer’s instructions) was added to the assay plates followed by incubation at room temperature for 2 min. The luminescence signal was determined in the luminescence mode of the ViewLux plate reader.
Filipin Staining
At the concentration used in the assay, Filipin dye (50 ng/mL) stains unesterified cholesterol in cells. 9 Cells were washed twice with PBS and fixed with 100 µL/well of a 3.2% formaldehyde solution including Hoechst 33342 stain at room temperature for 30 min. After washing twice with PBS, the cells were stained with 100 µL/well of 50 ng/mL Filipin solution (freshly dissolved in DMSO at 10 mg/mL and then diluted in PBS) at room temperature for 1 h. The cells were washed twice with PBS to remove Filipin solution, and the plates were stored at 4 °C for further imaging analysis. For imaging, cells were stained with 100 µL/well 2 µM of CellMask-Red (Invitrogen) in PBS at room temperature for 1 h. The plates were imaged using the Incell2000 Analyzer with a 20× or 40× objective lens and the filter sets for DAPI and TRITC to visualize Filipin and CellMask staining, respectively.
Data Analysis
Concentration-response curves were analyzed and EC50 values calculated using Prism software (GraphPad, San Diego, CA). The bottom value in the four-parameter fit was fixed at 50% because the concentration-response curves of MBCD showed an incomplete inhibition at the bottom plateau due to the limited high concentrations used. Results in the figures are expressed as the mean of triplicates ± SD unless they are specified.
Results
Mature cells differentiated from iPSCs such as neurons, cardiomyocytes, and hepatocytes are desirable for use in compound screens. However, the process of generating differentiated cells from iPSCs usually takes 6 to 12 weeks before the cells are ready for compound screens. Currently, the prolonged time and complicated procedure limit broad application of stem cell–derived human cells for drug discovery. We have explored the use of multipotent NSCs as the starting point to obtain differentiated neurons for compound screens, which significantly reduces the preparation time to obtain differentiated neurons. This approach begins with frozen iPSC-derived NSCs that can be plated into 96-well plates for continuous differentiation into neurons in the proper cell culture conditions. After 2 to 3 weeks of culture in differentiation medium, the cells are ready for characterization and compound screens. To validate this approach, we have examined the activity of MBCD on the reduction of cellular cholesterol levels in the differentiated neurons in comparison with the results obtained from primary skin fibroblasts, astrocytes and other cell lines.
Characterization of Neurons Differentiated from iPS Cells
In contrast to immortalized cell lines and primary cells, it is critical to characterize the differentiated cells to confirm the cell lineage before their use in cell-based and cytotoxicity assays. Molecular markers that are commonly used for characterizing neural stem and progenitor cells are Nestin, an intermediate filament protein, and Sox2 and Sox1 (transcription factors).10,11 We first performed immunofluorescence staining of neural stem cells using anti-Oct4 and anti-Nanog antibodies to test for markers of pluripotency. The lack of Oct4 and Nanog staining is consistent with a more restricted lineage status (multipotency instead of pluripotency) of neural stem cells compared with iPSCs ( Fig. 1A ). The NSCs, maintained in media supplemented with bFGF (see the Materials and Methods section), were positive for Nestin and Sox1, two markers of early neuroepithelial cells, prior to directed differentiation ( Fig. 1A ).
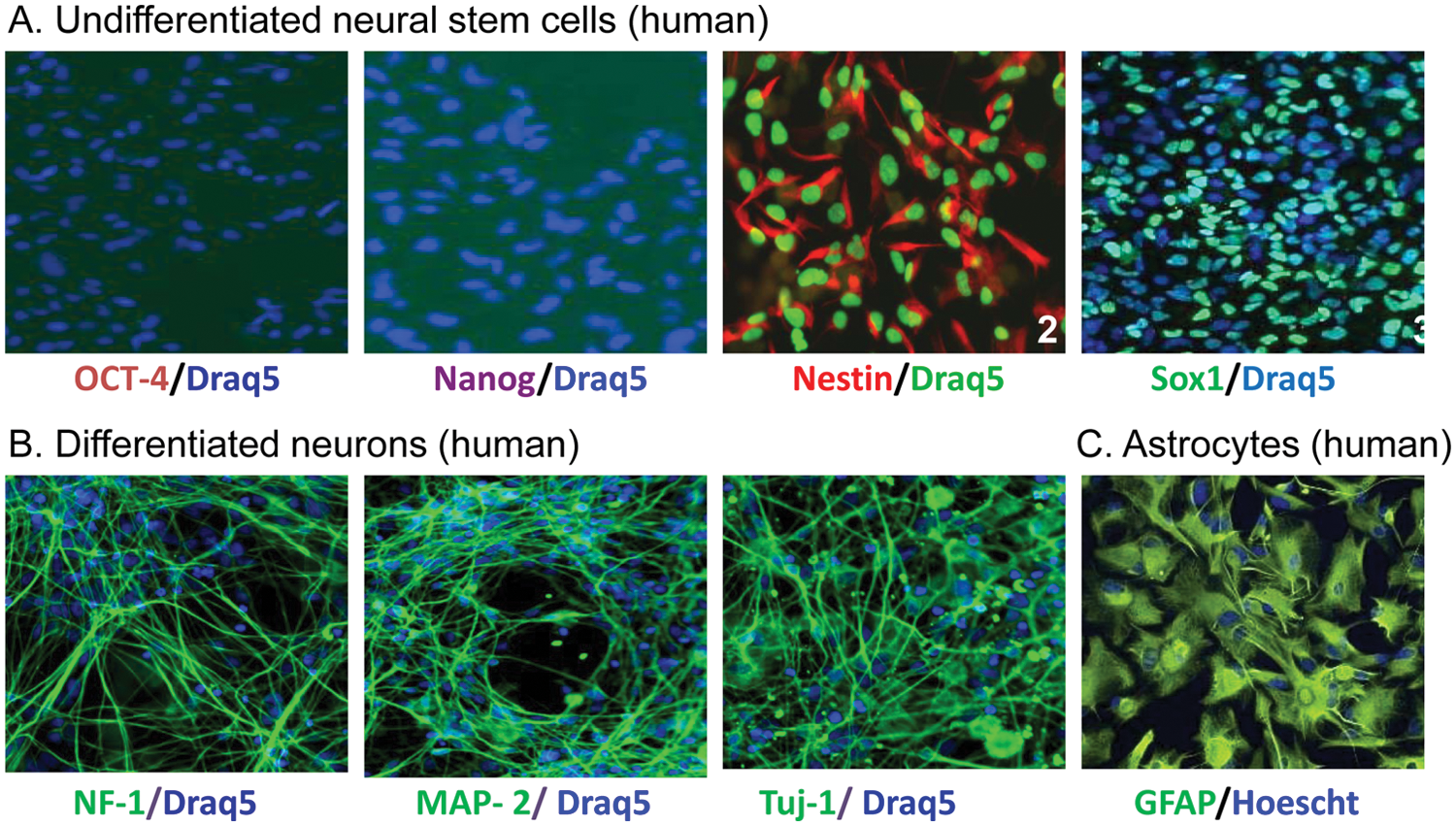
Fluorescence images of specific markers for stem cells and differentiated neurons. (
Switching of NSC media to media containing 10 ng/mL BDNF and 10 ng/mL GDNF (instead of bFGF) resulted in the differentiation of the majority of cells to a typical neuronal phenotype with a dense network of neurites. The neural lineages of the differentiated cells were analyzed by immunofluorescence staining using cell-type–specific markers. Phenotypically, these neurons were positive for β-tubulin III, microtubule associated protein 2 (MAP2), and neurofilament-1 ( Fig. 1B ). No immunoreactivity was detected in the neuronal cultures for GFAP, an astrocyte cell marker (data not shown). Immunofluorescence staining data revealed an abundance (90%) of neurons in our culture and correlated well with the high expression of β-tubulin III by real-time PCR (data not shown). Commercially available astrocytes were validated by immunofluorescence staining for GFAP ( Fig. 1C ).
MBCD Reduces Cellular Cholesterol Levels in Both Control and NPC1 Fibroblasts
β-cyclodextrins have been reported to reduce lysosomal cholesterol accumulation in skin fibroblasts derived from NPC patients as well as in the brain of an Npc1-/- mouse model, both detected by the Filipin staining assay.12–14 The Filipin dye stains unesterified free cholesterol in cells and is used for diagnosis of NPC disease. 9 NPC1 fibroblasts exhibited an excessive increase in Filipin fluorescence staining compared with the control cells ( Fig. 2A ), consistent with the published observation that the late endosomes and lysosomes are significantly enlarged in NPC1 cells due to the accumulation of free cholesterol and other lipids. 15 Treatment of NPC1 fibroblasts with 300 µM MBCD significantly reduced the fluorescence staining of Filipin ( Fig. 2A ). We then used a LysoTracker dye that specifically stains the acidic compartments in cells, which includes late endosomes and lysosomes, to confirm MBCD’s effect. 15 As in the Filipin staining experiment, the LysoTracker dye staining was significantly reduced after the treatment with 300 µM MBCD ( Fig. 2B ). These results confirmed the effect of MBCD on the reduction of cholesterol accumulation and lysosome size in NPC1 skin fibroblasts.
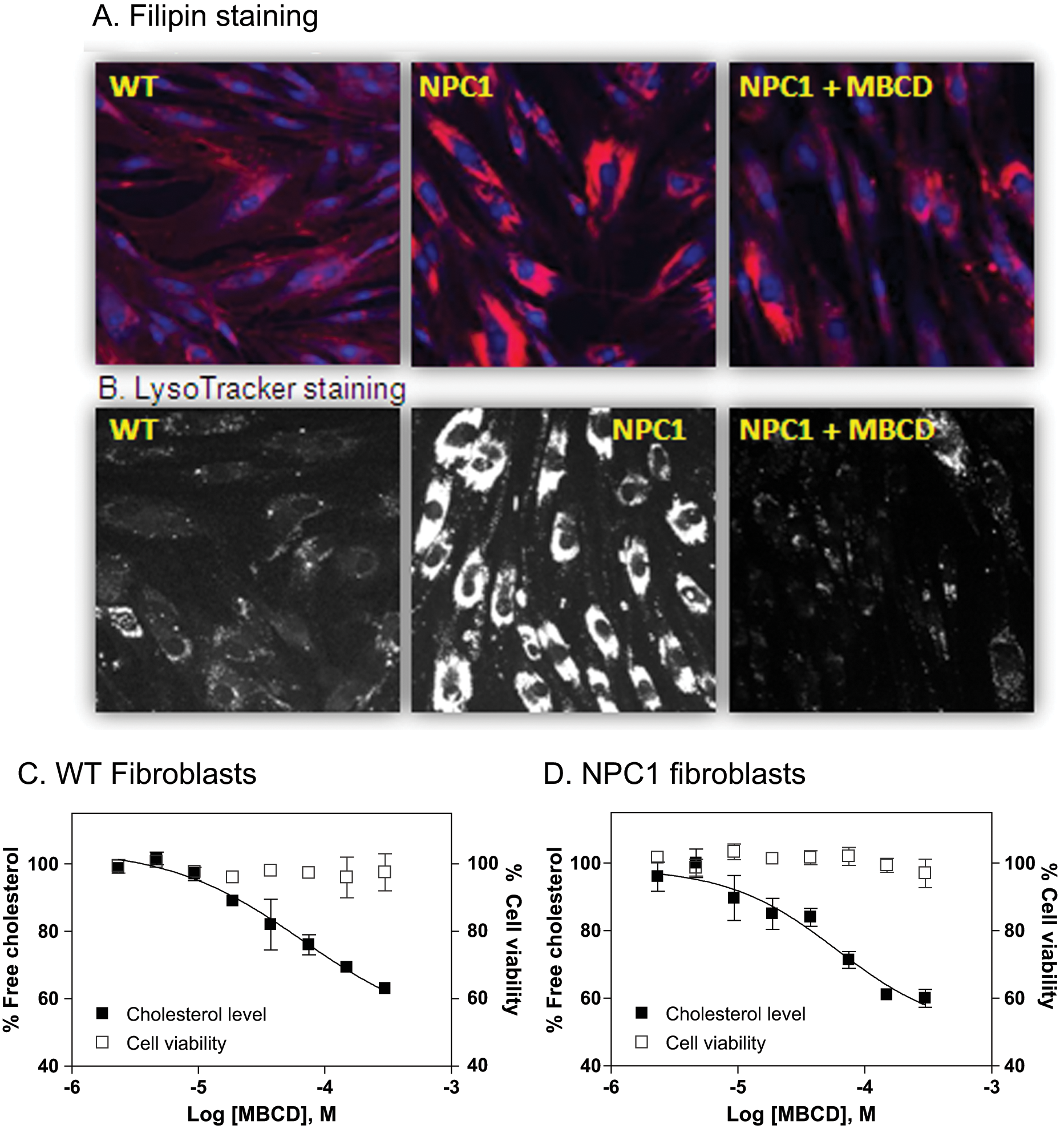
Effect of methyl-β-cyclodextrin (MBCD) on skin fibroblasts derived from NPC1 patients. (
We then employed an additional assay to confirm our findings—specifically, a biochemical assay to determine the cholesterol reduction effect of MBCD in both NPC1 and control fibroblasts. The cholesterol assay uses an enzyme-coupled reporting system that consists of cholesterol oxidase, horseradish peroxidase, and Amplex-red dye that involves a cell wash step to remove cell culture medium before the assay. As predicted, MBCD reduced the cellular unesterified cholesterol levels in both control and NPC1 fibroblasts in a concentration-dependent manner, although a complete inhibition curve at the lower plateau could not be achieved because of the cytotoxicity of MBCD at concentrations higher than 300 µM. The EC50 value of MBCD for reduction of cellular cholesterol level was 66.5 µM ( Fig. 2C ) in the control fibroblasts, which is similar to the EC50 of 60.5 µM determined in the NPC1 fibroblasts ( Fig. 2D ). Thus, the results indicate that normal cells could also be used to evaluate the cholesterol reduction effect of β-cyclodextrins or other compounds.
MBCD Reduces Cellular Cholesterol Levels in Differentiated Neurons and Additional Cell Types
Because NPC disease primarily affects the brain with neuronal degeneration as the cause of lethality, we wanted to examine the cholesterol reduction effect of MBCD in human neurons and, more desirably, in NPC disease neurons. In human neurons, the effect of β-cyclodextrins on cholesterol accumulation in late endosomes and lysosomes has not been studied because these cells are not generally available. An additional layer of technical difficulty applies to differentiated neurons, which are more fragile and may not be able to tolerate experiments that require multiple steps of cell wash, fixation, and dye staining included in the procedure of the Filipin staining assay. At this stage, iPSCs from NPC patients are not available, although attempts are being made to produce such cells (unpublished data). As an alternative approach, we determined the effect of MBCD in normal human neurons differentiated from iPSCs. Consistent with the data in fibroblasts, MBCD reduced the cellular cholesterol levels in differentiated neurons derived from normal human iPSCs with an EC50 value of 66.9 µM ( Fig. 3A ; Table 1 ). The EC50 values reported here are similar to that determined in patient-derived fibroblasts in this study ( Fig. 2 ) or reported elsewhere. 16 In human astrocytes, MBCD also reduced cellular cholesterol levels with an EC50 value of 110.7 µM ( Fig. 3B ). We then examined the effect of MBCD in a human neuroblastoma line, SH-SY5Y. The EC50 value of MBCD in the neuroblastoma cells was 81.9 µM ( Fig. 3C ), similar to that found for the normal skin fibroblasts. Since MBCD is cytotoxic at concentrations higher than 300 μM, its activity in the cholesterol assay was not shown for these concentrations, thus resulting in an incomplete concentration-response curve that is missing the lower plateau. We also monitored the cytotoxicity of MBCD using an ATP content assay in parallel to the biochemical cholesterol assay to ensure the compound concentration used did not cause cytotoxicity.
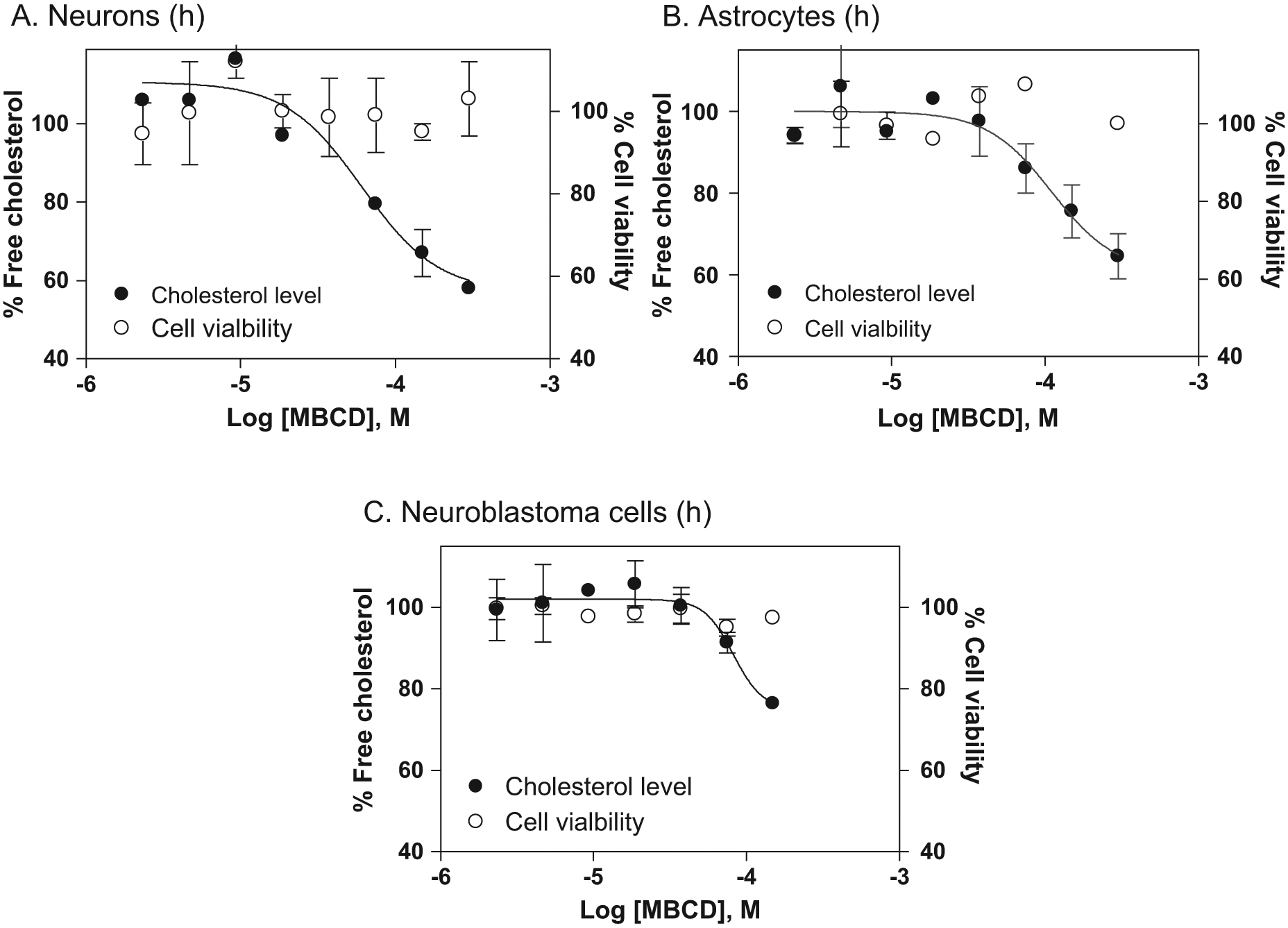
Concentration-response curves of methyl-β-cyclodextrin (MBCD) on reduction of cellular cholesterol levels in human neurons (
Summary of Methyl-β-cyclodextrin Activities on Reduction of Cellular Cholesterol
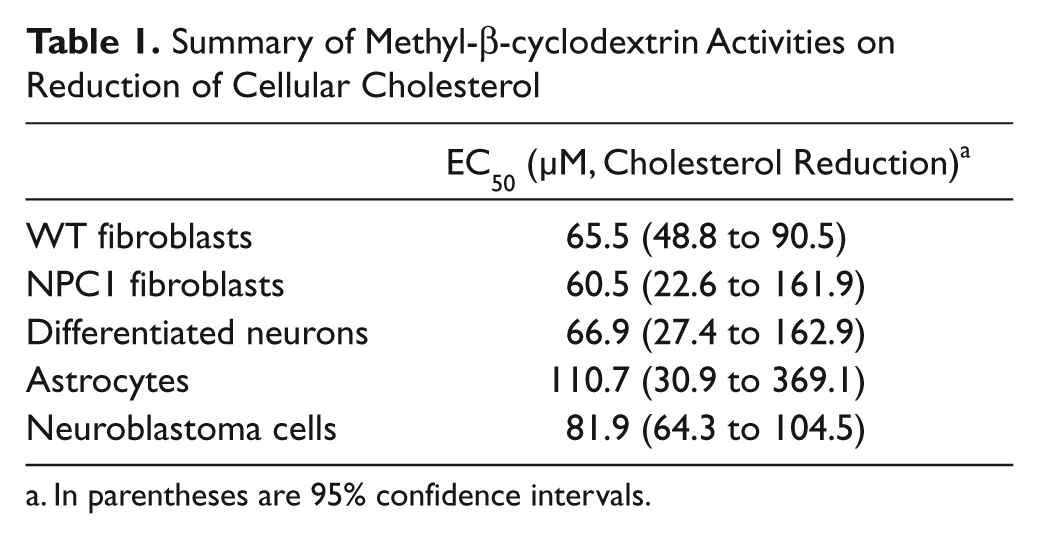
In parentheses are 95% confidence intervals.
Together, our results confirmed the activity of MBCD on the reduction of cellular cholesterol levels in human differentiated neurons and astrocytes. The data also demonstrated that human differentiated neurons (derived from NSCs) are a useful tool for confirmation of drug efficacy in human cells as well as for measuring compound cytotoxicity.
Discussion
In past two decades, HTS of molecular targets against large compound collections has become a major approach of drug discovery. Hits identified from the primary screen are confirmed in the secondary screens for target activity and selectivity. One or a few structurally appealing lead compounds subsequently undergo chemical optimization to improve potency and selectivity. The optimized compounds are then validated in the tertiary screens, which are more disease relevant and often use primary cells and cell-based disease models. Finally, a few top lead compounds are tested in the animal models before entering preclinical development for drug safety and ADME properties. A recent review of failed drugs in clinical studies revealed that overall, 31% of phase 2 and 61% of phase 3 trials failed because of lack of human efficacy (data from 2007–2011).17,18 These once promising drug candidates that have good drug efficacy in animal models may have a different effect in human patients, probably due to the species difference. 19 Thus, the current animal models may not always be good at predicting human efficacy, especially for certain diseases such as cancer and neuronal disorders. Therefore, the use of human cells and patient-derived disease cells for drug screens and validation of lead compounds has emerged as an alternative method to animal models that may improve the success rate of drug discovery and development.
Dependent on the origins of iPSCs generated, there are two major types of differentiated human cells: normal cells and disease-specific cells. The normal human iPSCs are readily available from several commercial sources such as the National Institutes of Health–sponsored Coriell Cell Repository (http://www.coriell.org/stem-cells) and American Type Culture Collection (ATCC). Normal iPSC-differentiated human cells such as cardiomyocytes, hepatocytes, and neurons have recently become available. Although several disease-related iPSCs have been generated and have become available, there have been many difficult cases for generation of iPSCs from patient cells. For example, the iPSCs from NPC disease cells have not been reported. Even though the disease-related iPSCs have been made, many of them do not exhibit the disease phenotype in the differentiated cells. In addition, large-scale production of differentiated cells from disease-related iPSCs may not be feasible in many cases. Therefore, the use of differentiated cells from normal human iPSCs, as used in this study, is a more practical approach for drug screens until the issues associated with disease-related iPSCs can be addressed. Specifically, using frozen cells that are already at an intermediate stage of differentiation, such as NSCs, for continued differentiation in microtiter plates can reduce the technical difficulties associated with a 6 week or longer differentiation process in microtiter plates, in addition to potentially reducing batch-to-batch variability in the differentiation process.
The generation of large quantities of differentiated cells with reproducible properties poses considerable challenges. In addition, it is not easy to produce disease models using cells differentiated from iPSCs. Several approaches may be employed for the assay development of cell-based disease models. First, disease models can be generated by addition of toxins or chemicals that damage cells or block a specific pathway in cells. In neuronal cells, the H2O2 challenge, 20 serum deprivation, 21 and toxins such as MPP+ 22 are commonly used to generate a model for examining neuronal protective activity of compounds. Second, a gene expression or pathway reporter assay can be set up in differentiated cells for the measurement of compound activity. Third, the cellular level of a protein, lipid, or metabolite in differentiated cells can be used for the determination of a specific therapeutic effect of compounds. In this study, we used a biochemical cholesterol assay to measure the effect of compounds on reduction of cellular cholesterol levels in human neurons differentiated from normal iPSCs.
Neuronal degeneration and massive loss of Purkinje cells in the brain are closely related to the pathogenesis of NPC disease. 7 Although the enlarged liver and spleen are often observed in NPC patients peripheral symptoms are usually rare. Thus, relevant drug therapies for the treatment of NPC disease have to target the central nervous system as the pathophysiology of NPC disease mainly affects the brain. In addition, these drug candidates should be able to penetrate the blood-brain barrier to exert its pharmacologic action in the central nervous system. Although skin fibroblasts derived from NPC patients have been used for compound screens, neurons differentiated from human iPSCs may be more useful as the secondary assays for evaluating drug efficacy and neuronal toxicity, in addition to confirming the pharmacologic action of drug candidates.
MBCD, a β-cyclodextrin, was previously reported to reduce lysosomal cholesterol accumulation in NPC1 fibroblasts and in the brain tissue of the NPC1-/- mouse model. The life span of these mice was also doubled after the treatment with β-cyclodextrins.12,14 The mechanism of action of β-cyclodextrin on the reduction of lysosomal accumulation of cholesterol and other lipids is linked to a calcium-dependent exocytosis of storage materials in the affected cells. 23 It has also been reported that β-cyclodextrin enters the cells through endocytosis. 16 Because the structure of β-cyclodextrin consists of seven sugars that are highly water soluble and impermeable to the blood-brain barrier, direct brain injection may be required for the therapeutic use of this compound in NPC patients. Our data demonstrate that 100 µM MBCD is nontoxic and reduces cellular cholesterol accumulation in human neurons differentiated from iPSCs.
In conclusion, we have developed an approach to use human neurons differentiated from normal iPSCs in a 96-well plate format to determine the efficacy of cellular cholesterol reduction and cytotoxicity of β-cyclodextrin compound. The application of starting neuronal differentiation from NSCs instead of iPSCs can significantly reduce the time required for cell differentiation and increase the reproducibility of cells used in a screen. As we demonstrate here for MBCD, the efficacy and potential cytotoxicity of compounds can be measured in parallel in human neurons differentiated from normal iPSCs, thus providing proof of concept that normal iPSC-derived cells can provide valuable information for the assessment of human efficacy and cytotoxicity of drug candidates and guide future clinical development.
Abbreviations
BDNF, brain-derived neurotrophic factor; bFGF, fibroblast growth factor; DPBS, Dulbecco’s phosphate-buffered saline; FBS, fetal bovine serum; GDNF, glial-derived neurotrophic factor; HTS, high-throughput screening; iPSCs, induced pluripotent stem cells; MAP2, microtubule-associated protein 2; MBCD, methyl-β-cyclodextrin; NF-L, neurofilament-L; NPC1, Niemann-Pick type C disease; NSCs, neural stem cells.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Intramural Research Program of the Therapeutics for Rare and Neglected Diseases, National Center for Advancing Translational Sciences, National Institutes of Health. We would also like to thank Dr. Anand Swaroop for comments on the manuscript.