
Abstract
Connexins are transmembrane proteins involved in gap junction intercellular communication. They present cell- and tissue-specific expression, with own electric and metabolic coupling specificities. These proteins are involved in numerous physiological processes in the brain and among them neuronal synchronization and trafficking of glucose. Such proteins are also described as being misregulated in various pathologies in the central nervous system. Thus, connexin blockers have been proposed as pharmacological tools to dissect these implications. However, such approaches lack accurate characterization of known inhibitors toward gap junction isoform specificity. In addition, those compounds are limited to few chemical classes and exhibit other activities, for example, an anti-inflammatory effect. The aims of this study were to evaluate the selectivity of described inhibitors and to enrich this pharmacopeia by new chemical classes. In this study, we present the specificity of published inhibitors toward several connexin isoforms expressed in the brain. Furthermore, after a screening of compounds using cellular models, we identified seven new inhibitors, with high functional reversibility and different relative selectivity toward isoforms. They constitute new chemical classes of connexin modulators completing those previously described. These new inhibitors might also provide new insights in understanding numerous pathophysiological processes involving gap junctions.
Introduction
Gap junctions are protein channels coupling the cytosol of adjacent mammalian cells in numerous tissues. This gap-junctional intercellular communication (GJIC) allows the direct transfer of ions, metabolites, adenosine triphosphate (ATP), and other molecules up to 1 kDa. 1 Gap junctions are composed by assembly of two connexons (or hemichannels), which are hexamers of connexin proteins. Twenty isoforms have been discovered in the human genome. 2 Several of them exhibit a specific cellular and tissue distribution, and some are expressed at different stages during embryological development. 3 Moreover, cells may express multiple connexins, and these connexins may form homochannels as well as heterochannels. 2 Connexins are involved in numerous physiological processes: intercellular communication, 1 regulation of growth control, apoptosis, neuronal synchronization, and formation of electric synapses. 4
For example, Cx32 is expressed in matured neurons and oligodendrocytes 3 and may be involved in metastatic tumors. 5 Moreover, a role in enhancing neuron survival and resistance in forms of ischemia has been proposed. 6 Moreover, along with Cx43, Cx40 forms gap junctions that couple atrial myocytes, 7 and in the CNS, they are expressed in astrocytes and developing neurons. 3 In addition, Cx40 expression is also altered after blood vessel injuries, 8 and Cx43 may be involved in numerous neurodegenerative diseases. 9 Altogether, these results indicate the high level of implication of connexins in human pathologies, notably for cerebrovascular disease, Alzheimer and Parkinson diseases, epilepsy, 3 neuropathic pain, and depression. 10
To better characterize gap junction physiological and pathophysiological activity, knockout models have been developed, but precocious lethality is often observed, and compensatory changes sometimes occur during development. 11 In addition, silencing RNA and antagonist peptides have been proposed, but these approaches are limited by their ability to reach all connected cells in complex tissues such as the CNS. 11 Several pharmacological gap junction inhibitors have been discovered. 12 However, their therapeutic uses are limited by the lack of knowledge of their specificities toward connexin isoforms 12 and by the fact that they belong to a few chemical classes (quinones, 13 fenamates, 14 licorice/glycyrrhetinic acid derivatives 15 ) with other in vivo therapeutic effects, notably anti-inflammatory properties. Such effects limit data interpretation on in vivo connexin inhibition.
Hence, to study the accurate implication of connexins in diverse processes and develop innovative treatments, we aimed at evaluating the specificity of described gap junction inhibitors toward different connexin isoforms. In addition, we planned on enriching this range by new chemical classes, which do not target inflammation pathways and might constitute innovative connexin modulators in physiological and pathological pathways.
To fulfill these objectives, we focused on the RIN β-cell line, isolated from a rat insulinoma, since it is well described as deficient in GJIC. 16 We focused our research on three human connexins—Cx32, Cx40, and Cx43—for their implication in human pathologies. We present here the establishment of new cellular models expressing these connexins. They allowed the characterization of the connexin channel specificity of classical inhibitors. Furthermore, after screening 162 molecules prescribed for human medicine in other indications, we identified seven compounds as new potent GJIC inhibitors.
Materials and Methods
Cell Culture
The rat insulinoma RIN cell line, deficient in GJIC, 16 was grown in OptiMem medium (Invitrogen, Saint-Aubin, France), supplemented with 10% fetal calf serum (Invitrogen). Cells were passaged twice a week and diluted 1:5.
Connexin Coding Plasmids, Cell Transfection, and Analysis of Gene Expression
GJB1 (Cx32), GJA5 (Cx40), and GJA1 (Cx43) open reading frames were amplified from human cDNA isolated from laboratory samples, using specific primers and proofreading Pfx polymerase (see primers and PCR conditions in
Genomic DNA was isolated using the DNeasy Blood & Tissue Kit (QIAGEN). RNA was extracted with the RNeasy kit (QIAGEN). First-strand cDNA was prepared from total RNA (10 µg) using Superscript II RT (Invitrogen). Expression of GJB1, GJA5, and GJA1 was evaluated by PCR, on genomic DNA and complementary DNA, using gene- or plasmid-specific primers.
Chemical Compounds
Chemical compounds such as quinidine, quinine, mefloquine, meclofenamic acid, flufenamic acid, niflumic acid, carbenoxolone, and 18-α- and 18-β-glycyrrhetinic acids, along with other compounds constituting the library, were bought from Sigma-Aldrich (Lyon, France) or Sequoia (Pangbourne, UK).
Dye Transfer Experiment
Cells were seeded in 24-well plates and loaded for 1 h at 37 °C with two fluorochromes, calcein acetoxymethyl ester (Calcein-AM, 1 µM; Sigma-Aldrich), a gap junction permeable dye, and Vybrant Dil (1 µM; Invitrogen), a membrane lipophilic dye, in serum-free medium. Cells were washed and grown overnight. The next day, cells were dissociated and incubated for 3 h in the presence of previously seeded nonloaded cells and in the presence of chemical compounds (n = 3 trials). Cells were dissociated and rinsed in phosphate-buffered solution, and flow cytometry was conducted on a FACScan (BD Biosciences, Rungis, France). Region of interest was determined based on the detection of nonfluorescent cells. Compensation between the two fluorochromes was obtained after cell analysis based on cells loaded either with calcein or DiL trackers, using FlowJo software (FlowJo, Tree Star Inc, USA).
Inhibition was quantified as the relative number of receiver cells that gained fluorescence to the total number of receiver cells (non–gap-junctional–mediated dye transfer was then subtracted from this ratio based on connexin nonexpressing RIN cells, defined at a background dye transfer ratio). This ratio of cellular coupling was then normalized, after each treatment, on the vehicle one. Data presented in the figures and Table 1 are the mean of three independent experiments.
Efficiency (IC50) and Toxicity (CD50) of Some Gap-Junctional Intercellular Communication Inhibitors
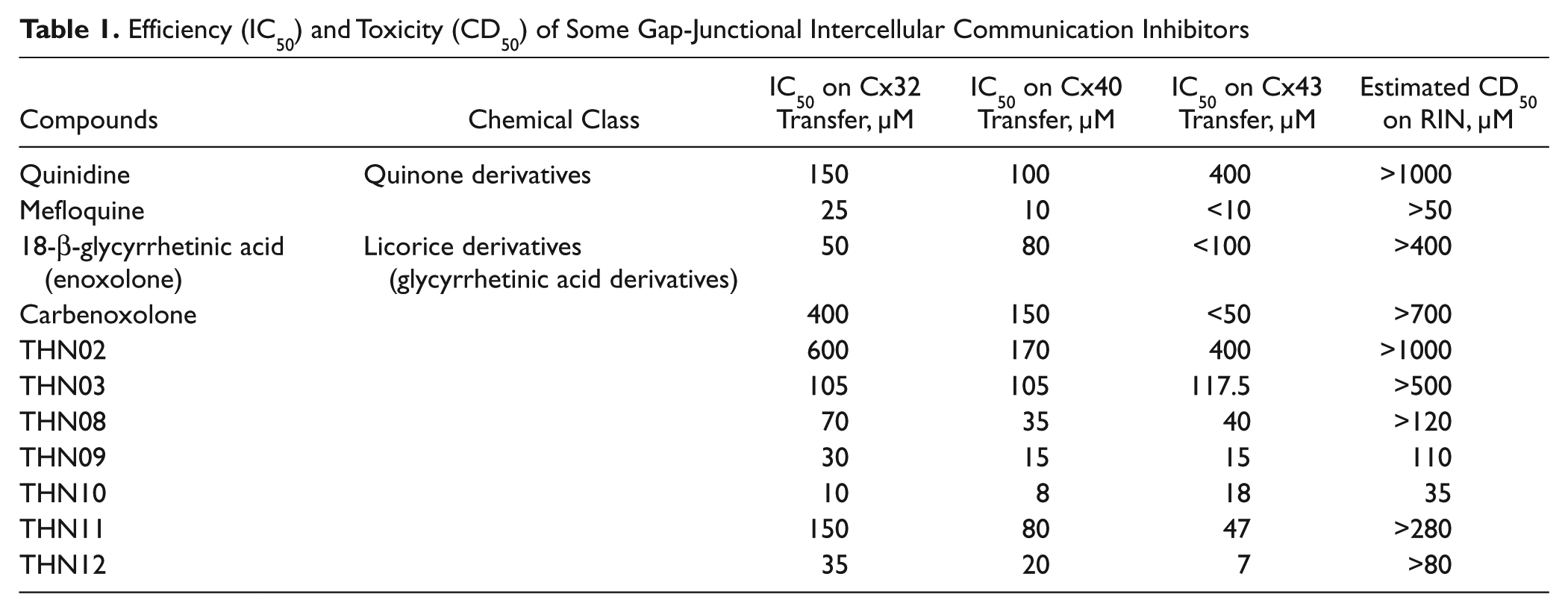
Inhibitory concentrations (IC50) were estimated graphically. Data (dye transfer or viability) are assessed to follow a sigmoid curve and are linearized using Y(data) = ln(1/data – 1). A linear regression (GraphPad Prism; GraphPad Software, La Jolla, CA) based on sigmoid assessment was realized to determine the equation of the sigmoid.
For the studies of gap junction inhibition reversibility, after seeding loaded cells on nonloaded cells, compounds were added for 2 h and cells were washed. Dye transfer rates were assessed the following hours.
Toxicity Analyses
In 96-well culture plates, 20 000 RIN cells were seeded in 100 µL fresh culture medium. They were treated with a range of dilutions of the compounds for 3 h, and viability was assessed after overnight incubation by a WST-1 test (Roche, Meylan, France). Cytotoxic dose (CD50) was then estimated graphically.
Statistical Analysis
Data are presented as mean ± standard deviation. Statistical analyses were obtained with GraphPad Prism, and significance of the results was calculated with t-tests or one-way analysis of variance (ANOVA) tests (*p < 0.05).
Results
Stable Connexin Expression in RIN, a Gap Junction–Deficient Cell Line
Plasmids coding for human Cx32, Cx40, and Cx43 were cloned and sequenced (Biofidal, Vaulx-en-Velin, France). RIN cells were transfected and selected by antibiotic resistance. The presence and expression of plasmids in these cells, further referred to as RIN32, RIN40, and RIN43, were assessed with plasmid- and gene-specific primers PCR (see Fig. 1A ). In addition, no differences in cell growth rate between RIN, RIN32, RIN40, and RIN43 cells were seen.
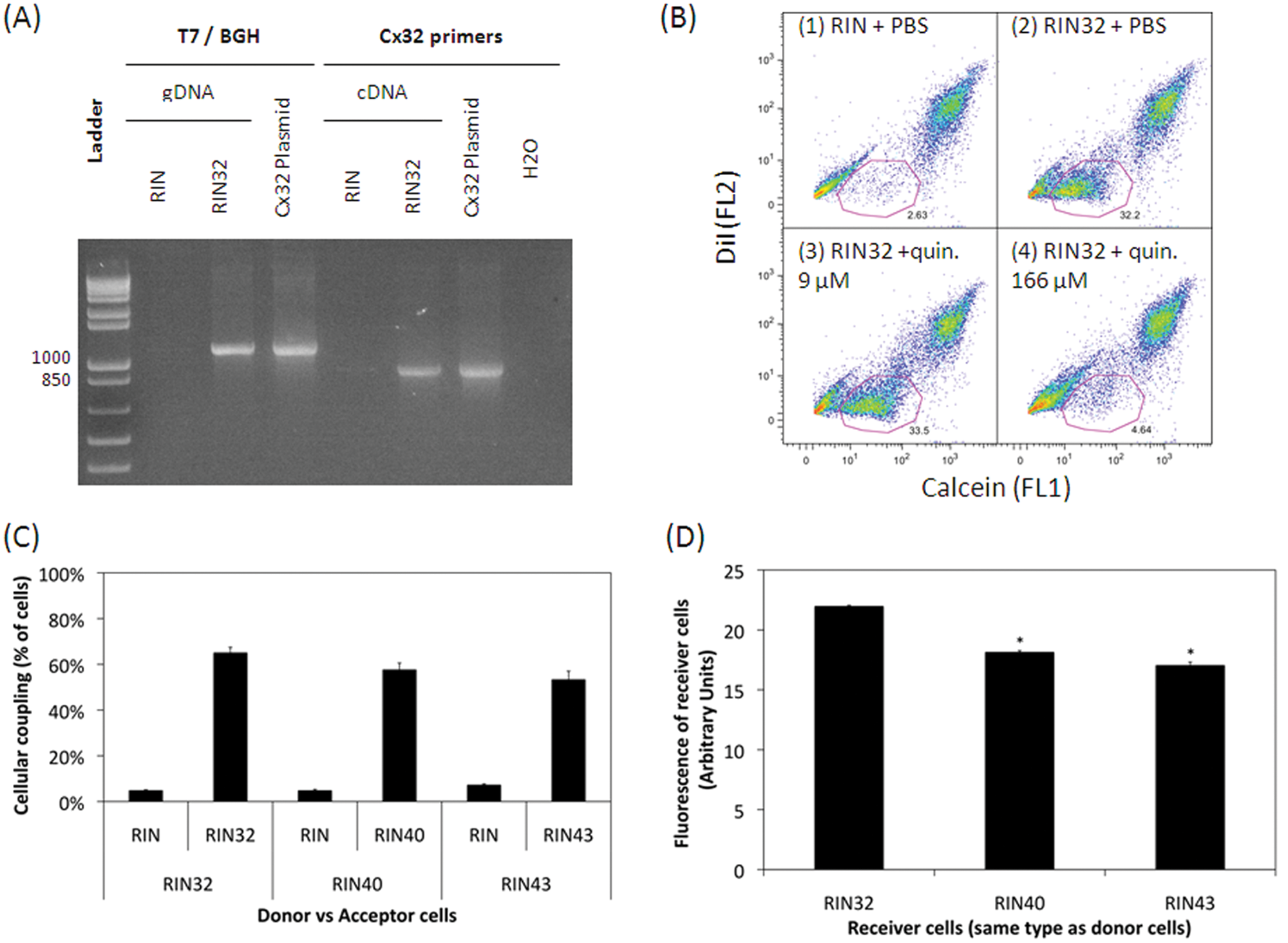
Evaluation of gap junction in Cx-expressing cells. (
Functionality of the Connexin-Expressing RIN Cell Line Compared with the RIN Cell Line
Functionality of the transfected connexin was estimated by flow cytometry analysis, with a modified parachute assay method. 17
RIN are deficient in GJIC ( Fig. 1B1 ). Transfection with a plasmid coding for Cx32 restores the functionality of gap junctions in RIN cells, as indicated by efficient dye transfer ( Fig. 1B2 ). Transfection with other coding plasmids also restored GJIC (data not shown).
Levels of cellular coupling were assessed by quantifying the number of nonfluorescent cells that gained fluorescence. Figure 1C indicates that gap junctions couple more than 50% of Cx-expressing cells. In addition, as shown in Figure 1D , we determined that dye transfer is more efficient in Cx32-expressing cells than in Cx40- or Cx43-expressing ones.
This gap junction activation can be inhibited by quinidine in a dose-dependent manner ( Figs. 1B3 , B4 and 2A ). Figure 2A shows the determination of an inhibitory concentration (IC50) of quinidine estimated at 100 µM. The inhibition profile of other known gap junction blockers has been established ( Fig. 3 and Table 1 ). Thus, quinone derivatives (quinidine) preferentially target Cx32 and Cx40, whereas some licorice derivatives (carbenoxolone) inhibit Cx43 and Cx40 more efficiently. Meanwhile, fenamate derivatives (meclofenamic acid, niflumic acid, flufenamic acid, mefenamic acid) showed low efficiency or inefficiency toward tested connexin isoforms at nontoxic doses (data not shown).
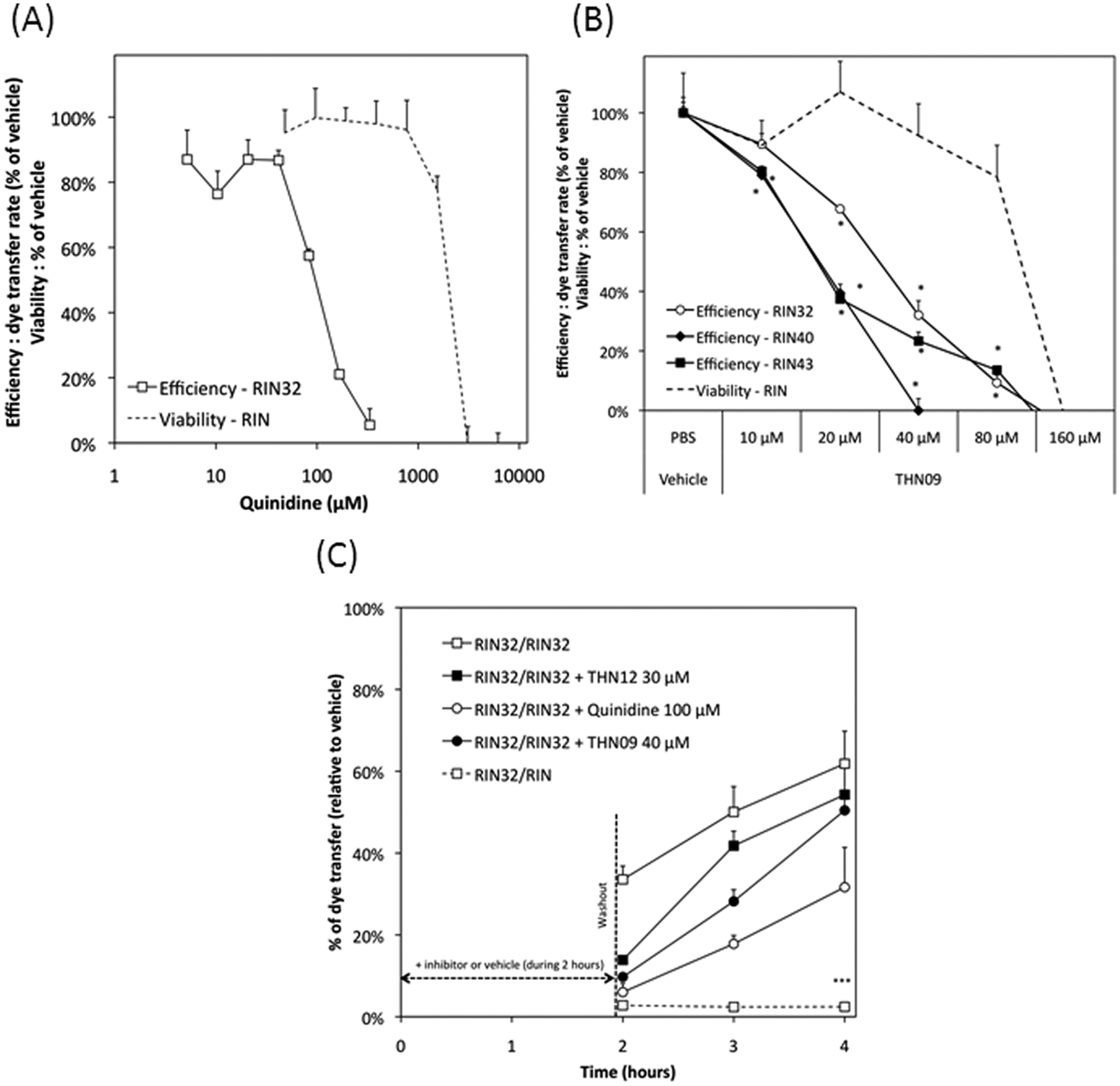
Connexin inhibition by diverse chemical compounds. (
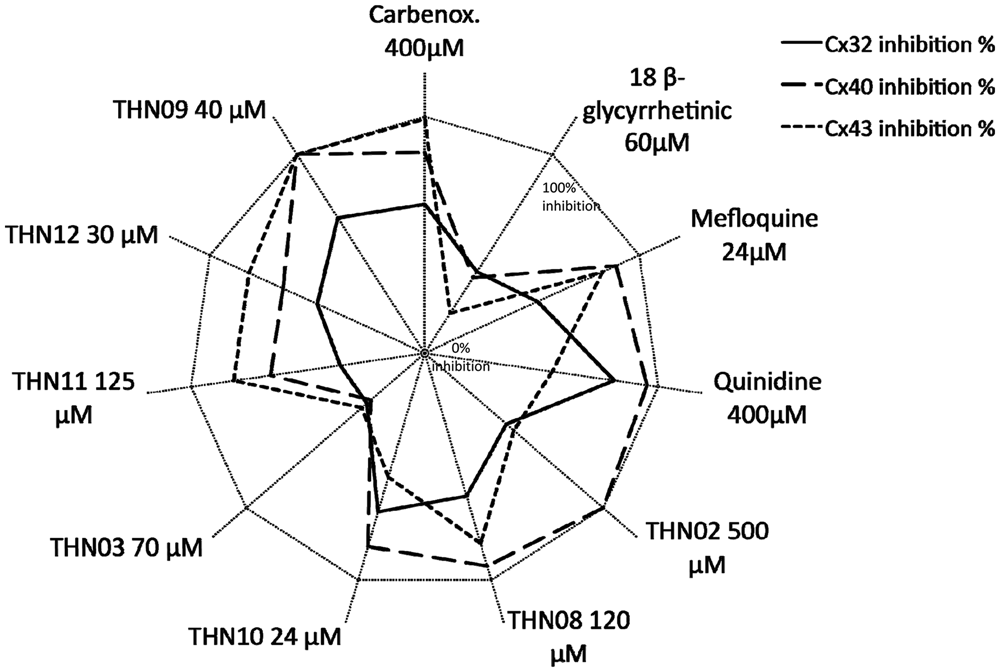
Inhibition spectra of several gap junction blockers. Cx32, Cx40, and Cx43 inhibition levels by different compounds were evaluated in cell culture. One hundred percent corresponds to full Cx inhibition and 0% to the absence of effect on gap-junctional functionality.
Screening of Compounds That Inhibit GJIC
One hundred sixty-two compounds, chosen among already genericized drugs and acting on other molecular targets, were tested at several concentrations (20–500 µM). Compounds were tested at soluble concentrations (for which no precipitate or suspension has been observed, with less than 0.1% DMSO in cell culture medium, and a DMSO concentration at which no effect on gap junction functionality was observed; data not shown). Among these compounds, seven presented reproducible gap junction inhibitions, in a dose-dependent manner, with no toxicity at inhibiting doses, as shown for one of these compounds in Figure 2B . As an example, THN09 showed specific inhibition of Cx40 and Cx43 and, to a lesser extent, Cx32 (see Table 1 ).
Reversibility of Gap Junction Inhibitors
Inhibition reversibility was evaluated by flow cytometry, and results are presented in Figure 2C . Quinidine, THN12, and THN09 showed reversible inhibition, but new compounds presented better reversibility than quinidine.
Discussion
Gap junction inhibitors are compounds that disrupt GJIC functionality. 18 Several methods have been successfully used in numerous laboratories to evaluate this functionality and their inhibition by gap junction blockers (for review, see Abbaci et al. 18 ). The method of preloading (or parachute assay) consists of preloading cells with fluorescent dyes and then letting them adhere to a confluent layer of unloaded cells. 19 Dye transfers are then evaluated by confocal microscopy or flow cytometry analysis. Such techniques have allowed the screening of large chemical libraries. 20 However, these techniques present some drawbacks: need to prior cell dissociation, allowing only endpoint analysis, and difficulty in precisely assessing time of cellular coupling. Nevertheless, flow cytometry parachute assay is one of the most adapted methods to study pharmacological response, for numerous reasons: (1) It allows the quantification of dye-coupled cells, (2) it maintains the cell integrity, (3) it allows a high number of analyzed cells for statistically reliable data, and (4) it gives reproducible results.
Establishment of Functional Connexin-Expressing Cells
Between 5% and 10% of nonloaded RIN cells are calcein positive after incubation with preloaded RIN or Cx-expressing cells. Probenicid, a pannexin inhibitor, 21 has no effect on this nonspecific dye transfer, and none of the tested compounds inhibited this nonspecific transfer (data not shown). This nonspecific transfer has been described in other studies 22 and could be explained by an alternative way of dye transfer.
Besides, significant dye transfer confirmed that the transfection with GJB1, GJA1, and GJA5 plasmids restored gap junction functionality and therefore allowed robust human Cx32, Cx40, and Cx43 expression. Dye couplings were not observed after medium transfer from preloaded to nonloaded cells, indicating the absence of hemichannel-mediated dye transfer. Moreover, such level of dye coupling has been evaluated elsewhere 19 and is significantly more pronounced in Cx32-expressing cells (Cx32 > Cx40 = Cx43). That could indicate (1) a higher level of Cx32 expression that should be further tested; (2) a better functionality of Cx32 in RIN32 cells; (3) a different metabolism of Cx32 in RIN cells; (4) a difference in initial levels of fluorescence that, however, did not appear in our analysis of preloaded cells; or (5) a more permissive specificity of Cx32 to calcein transfer due to its own structural characteristics compared with other connexins. 23
Furthermore, no significant differences in population doubling time were found, indicating that RIN might constitute a connexin-independent proliferating cell model, completing those described elsewhere. 24
Specificity of Known Gap Junction Inhibitors on Human Gap Junction Isoforms
Connexins have been involved in numerous pathologies and are, for example, overexpressed in neuropathic pain, which results in neuronal sensitization and excitotoxic neuronal cell death. 25 Furthermore, intrathecal administration of carbenoxolone reduces neuropathic pain after sciatic nerve inflammation. In parallel, upregulation of connexins is often described in neurodegenerative disorders 3 or depression, 10 and blocking of glial connexin has been proposed elsewhere to evaluate their contribution. 11 Since similar implications of several isoforms have been described, establishing that the inhibition profile of known blockers might be of interest to evaluate this implication.
First, we confirmed the fact that glycyrrhizic acid does not inhibit the tested gap junctions as described elsewhere.
15
Some glycyrrhetinic acid derivatives such as carbenoxolone and β-form, known as gap junctions blockers,
13
showed different GJIC inhibition profiles (see
We pointed the high efficiency of mefloquine on gap junction inhibition, not only on Cx26, Cx32, Cx36, Cx43, and Cx50, as previously described, 27 but also on Cx40 at low doses. This compound is therefore a useful agent to evaluate physiological roles of different connexin subtypes simultaneously and phenotypes in connexin-deficient mice.
Quinine and quinidine, its stereisomer, are two gap junction inhibitors, 28 and we report here this activity in the blocking of dye transfer in Cx32-expressing cells. Nevertheless, quinine has been described as not substantially affecting Cx32 gap junction–mediated electric current transmission (at concentrations below 500 µM 28 ). This apparent discrepancy might suggest that quinine could induce blockade of chemical transfer but not of electric current. It would therefore be of interest to test transfected cells using the whole-cell voltage clamp technique to dissect this particular mechanistic dichotomy. Moreover, identification of blockers that specifically inhibit metabolic or electric coupling might provide interesting tools to enrich data about metabolic trafficking through astrocytes. 29
We also evaluated fenamate derivatives. Surprisingly, we showed that fenamates were not efficient at connexin metabolic coupling, contrary to previously published data on gap junction electric inhibition by fenamates. 14 Therefore, similarly to quinidine, fenamates might allow a dichotomy between these two cellular couplings.
To summarize the inhibition profile of known gap junction inhibitors, high levels of inhibition of Cx40 and Cx32 are achieved by quinidine (quinidine > mefloquine = carbenoxolone for Cx40, quinidine > carbenoxolone > mefloquine for Cx32), and Cx43 is highly inhibited by carbenoxolone (carbenoxolone > mefloquine > quinidine).
Identification and Characterization of New Gap Junction Inhibitors
Classical gap junction inhibitors, such as carbenoxolone or quinone derivatives, belong to few chemical classes, 12 and most of them present anti-inflammatory activity or antiarrhythmic effects. Therefore, interpretation of in vivo pharmacological modulation of GJIC by these compounds is limited by their other therapeutic effects (notably on inflammation or cardiac rhythm). New modulators, however, with their own effects but different from classical inhibitors, might therefore circumvent this limitation by extending the range of gap junction inhibitors.
To try and identify new GJIC inhibitors for CNS use, we selected drugs according to some of these diverse criteria: (1) their ability to cross the blood-brain barrier, (2) their belonging to a chemical class different from classic gap junction inhibitors, and (3) their availability in human pharmacopeia for easier drug repositioning. One hundred sixty-two compounds were therefore selected and screened using RIN32, RIN40, and RIN43 cell cultures. Seven new compounds blocking gap junction–mediated dye transfer were therefore identified and presented inhibition selectivity to Cx32, Cx40, and Cx43. Consequently, and due to the criterion of the constitution of the library, they might constitute valuable pharmacological in vivo modulators of connexins.
Reversibility of gap junction inhibitors has been previously shown for quinone derivatives 28 ; we tested this reversibility using an approach different from a previously described method. 28 Due to their high efficiency in dye transfer, we confirmed here on RIN32 cells that quinidine, a quinone derivative, effectively presents a reversible effect on gap junction (either on established channels or gap junction recycling). Nevertheless, we showed interesting data on the higher reversibility rate of newly identified gap junction modulators, such as THN12 and THN09, indicating promising new pharmacological tools.
Moreover, we demonstrated that THN09 showed specific inhibition of Cx40 and Cx43 and, to a lesser extent, Cx32 (Cx40 > Cx43 > Cx32). Six other compounds showed higher inhibition efficiency on Cx32 (THN08 > quinidine), Cx40 (THN02 = THN09 > quinidine), or Cx43 (THN11 = THN12). More important, contrary to known GJIC blockers that present low specificity, we showed a high inhibition profile of THN02 on Cx40. These data complete those obtained elsewhere on other gap junctions such as 2-aminoethoxydiphenyl borate (2-APB; Cx40 > Cx43 > Cx32). 30
Consequently, we identified compounds with a unique spectrum of gap junction inhibition presenting high or low reversibility. Further structure-activity relationships should lead to the identification of precise chemical groups in such compounds involved in a specific property of gap junction inhibitors.
In conclusion, we successfully defined a new human Cx32-, Cx40-, and Cx43-expressing cellular system, which allowed the study of these connexins in cell culture. We characterized the inhibition profile of known gap junction inhibitors, which could help determine their specificity toward cell-type isoforms. We also identified seven new compounds for their blocking of gap junctions, with (1) a new and better inhibition profile, (2) a new reversible effect on GJIC inhibition, and (3) no described effects on inflammation or cardiac rhythm. These new gap junction inhibitors might constitute interesting candidates for pharmacological in vitro and in vivo blocking of specific connexins since (1) they are already prescribed for human health and numerous data are available concerning their pharmacology in human and animals, (2) they belong to other chemical classes than classically described gap junction inhibitors, (3) they can cross the blood-brain barrier after peripheral administration, and (4) they show specific dose-dependent human Cx32, Cx40, or Cx43 inhibition.
In addition and from our point of view, these results emphasize the possible role of connexins in the pharmacological effect of several drugs proposed initially in human health.
The determination of an inhibition profile of new compounds identified in this study and known GJIC blockers might be of interest not only to evaluate the involvement of connexin subtypes in human pathologies but also to constitute innovative therapies, which might inhibit specific connexins involved in these pathologies.
Footnotes
Acknowledgements
The RIN cell line was a generous gift from Pr. Paolo Meda (University of Geneva, School of Medicine). We warmly thank Dr. Laurent Cronier (Université Poitiers) and Dr. Valérie de Broglie (Fondation Alliance BioSecure) for providing scientific and technical assistance, as well as the Program “Technologies pour la Santé” (Drs. Jacques Grassi and Raymont Pommet) and iMETI (Pr. Philippe Leboulch, Dr. Jean-Philippe Deslys) for their support.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.