
Abstract
CEfrag is a new fragment screening technology based on affinity capillary electrophoresis (ACE). Here we report on the development of a mobility shift competition assay using full-length human heat shock protein 90α (Hsp90α), radicicol as the competitor probe ligand, and successful screening of the Selcia fragment library. The CEfrag assay was able to detect weaker affinity (IC50 >500 µM) fragments than were detected by a fluorescence polarization competition assay using FITC-labeled geldanamycin. The binding site of selected fragments was determined by co-crystallization with recombinant Hsp90α N-terminal domain and X-ray analysis. The results of this study confirm that CEfrag is a sensitive microscale technique enabling detection of fragments binding to the biological target in near-physiological solution.
Introduction
Fragment-based drug discovery (FBDD) is a well- established and successful drug discovery approach widely adopted by the pharmaceutical community. FBDD relies on the use of sensitive biophysical techniques, such as X-ray crystallography, 1 nuclear magnetic resonance, 2 and surface plasmon resonance (SPR), 3 to be able to detect the often weak affinity interactions (Kd >10 µM) of the low molecular weight (<300 Da) drug fragments with the target of interest. It is important to validate fragment hits using a number of techniques to help prioritize fragment follow-up. Capillary electrophoresis is a well-known analytical technique, which has recently been adapted for fragment screening and termed CEfrag. The main advantages that CEfrag have over other fragment screening techniques are lower protein consumption, higher throughput, and the fact that screening can be carried out using unmodified protein in solution. This last point means that feasibility studies and assay setup can be rapidly carried out as there is no need to tether the protein.
Capillary electrophoresis (CE) is a high-resolution, microscale electrophoretic technique that rapidly separates both charged and uncharged molecules. CE instrumentation consists of capillary, electrodes, a high-voltage power supply, a detector, and a data acquisition and handling device. The source vial, destination vial, and capillary are filled with an electrolyte buffer (the “running buffer”). The sample is introduced into the capillary by capillary action or pressure, and then voltage is applied, inducing migration of the analytes. Ionic species are separated by their charge and hydrodynamic radius, and uncharged species can be separated by introducing electro-osmotic flow (EOF). For a more detailed introduction to CE and its application, see Camilleri. 4
Affinity CE (ACE) is a specific application of CE used to detect biomolecular interactions under near-physiological conditions. With ACE, interacting molecules are detected by monitoring changes in the electrophoretic mobility of one or both components upon binding. ACE has been successfully used to detect many drug-protein and protein-protein interactions.5-7 Applications of ACE for drug screening were pioneered by Cetek Corporation5-8 and recently extended by Selcia Ltd to include fragment screening (CEfrag). 9 CEfrag relies on the use of a known competitive ligand with affinity to the binding site of interest on the target protein (the “probe ligand”) and detects a reduction in the interaction of the probe ligand with its target protein in the presence of test compounds (e.g., fragments) binding to the target. Binding occurs in free solution using physiological buffers, thus preventing artifacts that may result from target immobilization, which is a requirement for some techniques such as SPR. The use of a probe ligand and the fact that CE is a visual separation technique help to rule out false positives often caused by nonspecific binding. CEfrag is suited to screen for fragment inhibitors of drug-protein, protein-protein, protein-nucleic acid, or other interactions between biomolecules.
We present here the case of a small-molecule–protein interaction using Hsp90. Heat shock protein 90 (Hsp90) is a molecular chaperone that is upregulated in response to stress and is of current interest as a therapeutic target in oncology. 10 Fragment screening to identify inhibitors of Hsp90 has been performed successfully by others using alternative fragment screening techniques11–15; these approaches have resulted in several compounds currently in clinical development. 16 Hsp90 was therefore deemed to be a good test case to compare the performance of CEfrag with other fragment screening technologies. The study described here used human full-length Hsp90 in conjunction with the Selcia Fragment Library and the known Hsp90 ligand radicicol as a probe ligand. 17 Fragments identified as hits in the CEfrag assay were further evaluated using a fluorescence polarization assay and X-ray crystallography.
Materials and Methods
Materials
Human recombinant full-length Hsp90α was purchased from Assay Designs (now part of Enzo Life Sciences, Exeter, UK). Radicicol was purchased from Tocris Bioscience (Bristol, UK). Unless otherwise specified, all other reagents were obtained from Sigma-Aldrich (Poole, Dorset, UK).
The Selcia fragment library was designed based on rule-of-three (Ro3) criteria 18 and fragmentation of known biologically active compounds (C. Swain, unpublished data). Fragments were selected for aqueous solubility of ≥1 mM under assay conditions. Briefly, 30-mM DMSO stock solutions were diluted to 1 mM in 35 mM HEPES (pH 7.8), and a turbidimetric method19 was used to assess solubility. Structural integrity was confirmed and purity assessed for all compounds in the library by proton nuclear magnetic resonance ( 1 H-NMR) using a Bruker Avance 500-MHz instrument (Bruker, Billerica, MA). Fragments were >95% pure. For the Hsp90 screen, a subset (609 compounds) of the Selcia fragment library was used.
Expression of Recombinant Hsp90 N-terminal ATPase Domain (Hsp90N)
A synthetic DNA fragment (GenScript, Inc., Piscataway, NJ) with optimized Escherichia coli codons and encoding residues 131–358 (NCBI accession # NP_001017963) of human Hsp90α was cloned into plasmid pET28a (Novagen, Madison, WI) and the resulting plasmid transformed into E. coli BL21 (DE3). The expressed protein was purified by nickel affinity and gel filtration chromatography as described previously. 11
CEfrag Assay and Screening of the Selcia Fragment Library
The Hsp90 CEfrag assay was developed on a Beckman P/ACE MDQ capillary electrophoresis instrument (Beckman Coulter, Inc, Brea, CA); screening was performed on the Geminis, 4-capillary instruments developed by Cetek Corp (Marlborough, MA). 20 Uncoated capillaries, 50 µm i.d. × 375 µm o.d. × 30 cm (Polymicro Technologies, Phoenix, AZ), were prefilled with Tris-Tween running buffer (10 mM Tris-HCl [pH 7.5], 0.0005% Tween-20, 5 mM MgCl2) with or without inhibitor (i.e., fragment) or DMSO as a control, with or without 0.25 µM Hsp90α. Probe ligand, radicicol (150 µM) in Tris-Tween buffer, was injected at 0.5 psi for 5 s (approximately 20 nL injection plug volume), and a separation voltage of 15 kV in reverse polarity (i.e., cathode to anode) was applied; the mobility of radicicol was monitored by UV at 254 nm. Fragments were initially screened at 500 µM; the final concentration of DMSO in all runs was kept constant at 1.67%. Primary hits were confirmed in a repeat screen of the hits. The percent inhibition was determined from the following equation: 100(TH – TL)/(TH –TR), where TH = migration time of radicicol peak in the presence of Hsp90 in the running buffer, TL = migration time of radicicol peak in the presence of Hsp90 and test ligand in the running buffer, and TR= migration time of radicicol alone. The hits were scored as strong (>50% inhibition) or weak (20%–50% inhibition).
Fluorescence Polarization Assay
The assay was established using fluorescein isothiocyanate- labeled geldanamycin (FITC-GM, from StressMarq, Victoria, Canada) and a modified version of the method described by Llauger-Bufi et al. 21 Briefly, 50 nM Hsp90 was incubated in black 96-well HE plates (Molecular Devices, Wokingham, Berkshire, UK) in the presence of 25 nM FITC-GM, with or without inhibitor, in assay buffer consisting of 20 mM HEPES (pH 7.2), 50 mM KCl, 0.1 mg/mL bovine serum albumin (BSA), 5 mM MgCl2, 1 mM dithiothreitol (DTT), and 0.01% NP40. Controls in the absence of enzyme were performed. The final DMSO concentration in the presence of inhibitor was 0.05% DMSO. The plates were incubated at 4 °C for 18 h; fluorescence polarization was then measured using a SpectraMax M5 (Molecular Devices) with excitation at 485 nm and emission at 530 nm. Fluorescence anisotropy was calculated, and results were expressed as percentage inhibition compared with the no-inhibitor control.
X-ray Crystallography
The purified Hsp90N protein was crystallized by hanging drop vapor diffusion with and without bound ligand using conditions (0.2 M MgCl2, 0.1 M HEPES, 20% w/v PEG2000, pH 7.2, incubated at 4 °C) established by others. 11 Ligands were dissolved in DMSO to produce concentrated stock solutions and added to the protein solution to achieve the required protein-ligand molar ratio. Crystals were flash frozen in oil prior to data collection at 100K using a Bruker X8 prospector X-ray generator (Cu-Kα radiation) with APEXII CCD detector. Diffraction images were processed and integrated with PROTIUM2 and structures determined by molecular replacement with PHASER 22 using a ligand-free Hsp90α N terminal domain structure (pdb code 2XDL) as the search model. The structures were refined with REFAMAC5, 23 and model building, including ligand fitting, was completed with COOT. 24 Ligand dictionaries were generated from SMILE strings using the CCP4 Libcheck program. 25 Structures of ligand-free Hsp90N, Hsp90N-radicicol, Hsp90N–fragment 6 (N,N-diethyl-4-hydroxy-3-methoxy-benzamide), Hsp90N–fragment 7 ((4-hydroxy- phenyl)-morpholin-4-yl-methanone), and Hsp90N–fragment 9 (4-ethylsulfanyl-6-methyl-[1,3,5]triazin-2-ylamine) complexes were validated using MolProbity. 26
Results
Hsp90 CEfrag Assay Development and Screening of the Selcia Fragment Library
The Hsp90 CEfrag assay was established using radicicol as the injected probe ligand; its electrophoretic mobility was monitored by UV at 254 nm (top trace, Fig. 1 ). In the presence of full-length Hsp90 in the running buffer, the migration time of the radicicol peak was significantly increased due to interaction of radicicol with Hsp90 during the capillary runtime (second trace, Fig. 1 ). In the presence of an Hsp90 binding fragment (fragment 1: 4-chloro- benzene-1,3-diol; see Table 1 ) in the running buffer, the radicicol/Hsp90 interaction was dose dependently reduced (traces 3–8, Fig. 1 ). This manifested as a reduction in the Hsp90-dependent migration time shift depending on the affinity of the fragment for Hsp90. Fragment 1 ( Table 1 ) is a resorcinol fragment, whose structure is present in radicicol ( Fig. 1 ); such resorcinol fragments are well-known inhibitors of Hsp90. 27 Other resorcinol fragments were also detected by CEfrag (fragment 2: 5-methoxy-benzene-1,3-diol and fragment 3: 4-ethyl-benzene-1,3-diol); their activity in the fluorescence polarization (FP) assay suggests these fragments are relatively weak ( Table 1 ). A simpler resorcinol fragment (fragment 4: benzene-1,3-diol) was not detected either in the FP assay or by CEfrag.
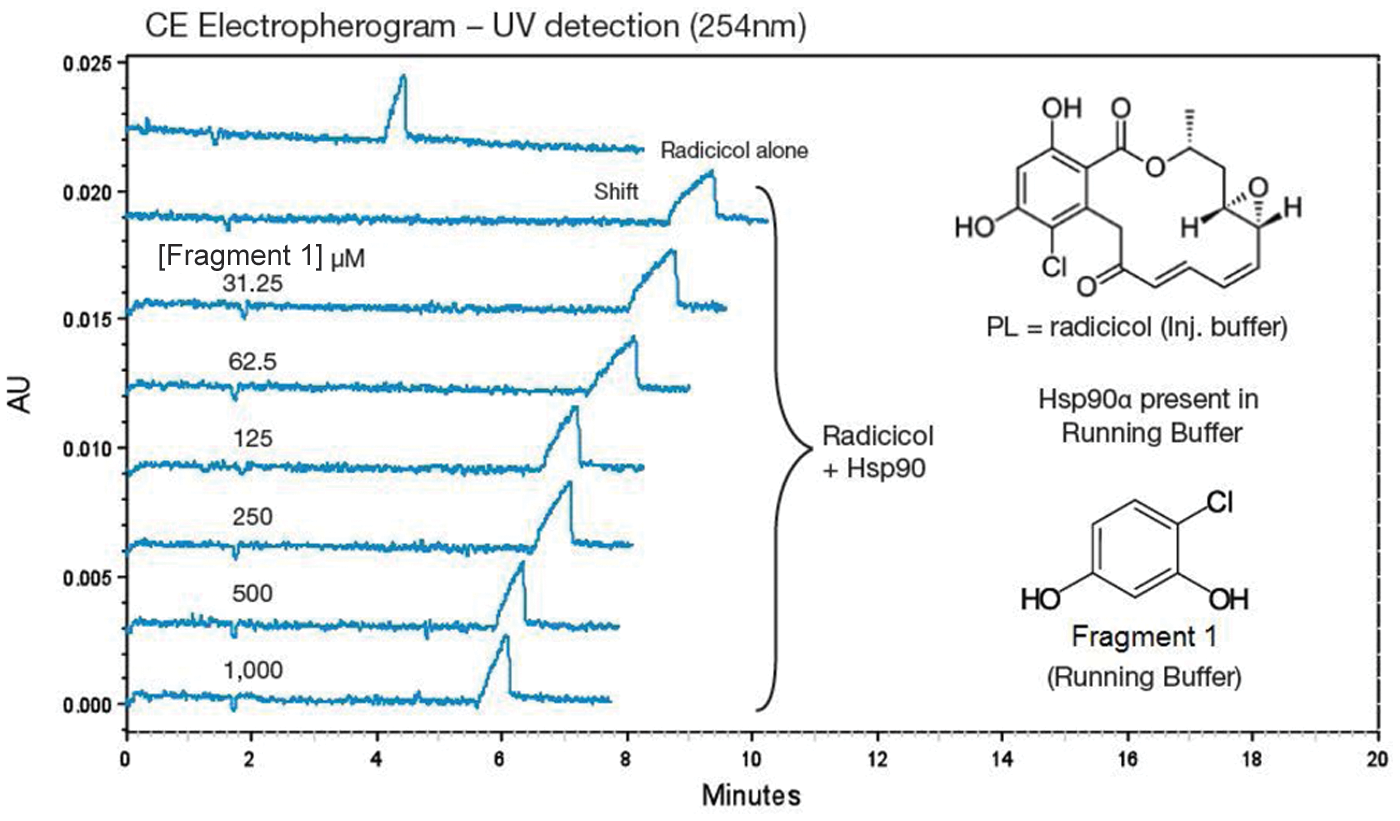
Exemplar capillary electrophoresis (CE) electropherograms from the Hsp90 CEfrag assay development. Mobility of radicicol in the absence and presence of Hsp90 (top two curves, respectively) and with increasing concentrations of fragment 1 in the running buffer. Traces were generated using a Beckman P/ACE MDQ CE instrument. AU, absorbance unit.
Structures, CEfrag, FP Assay IC50 Values, Ligand Efficiencies, and X-ray Summary of Selected Fragments
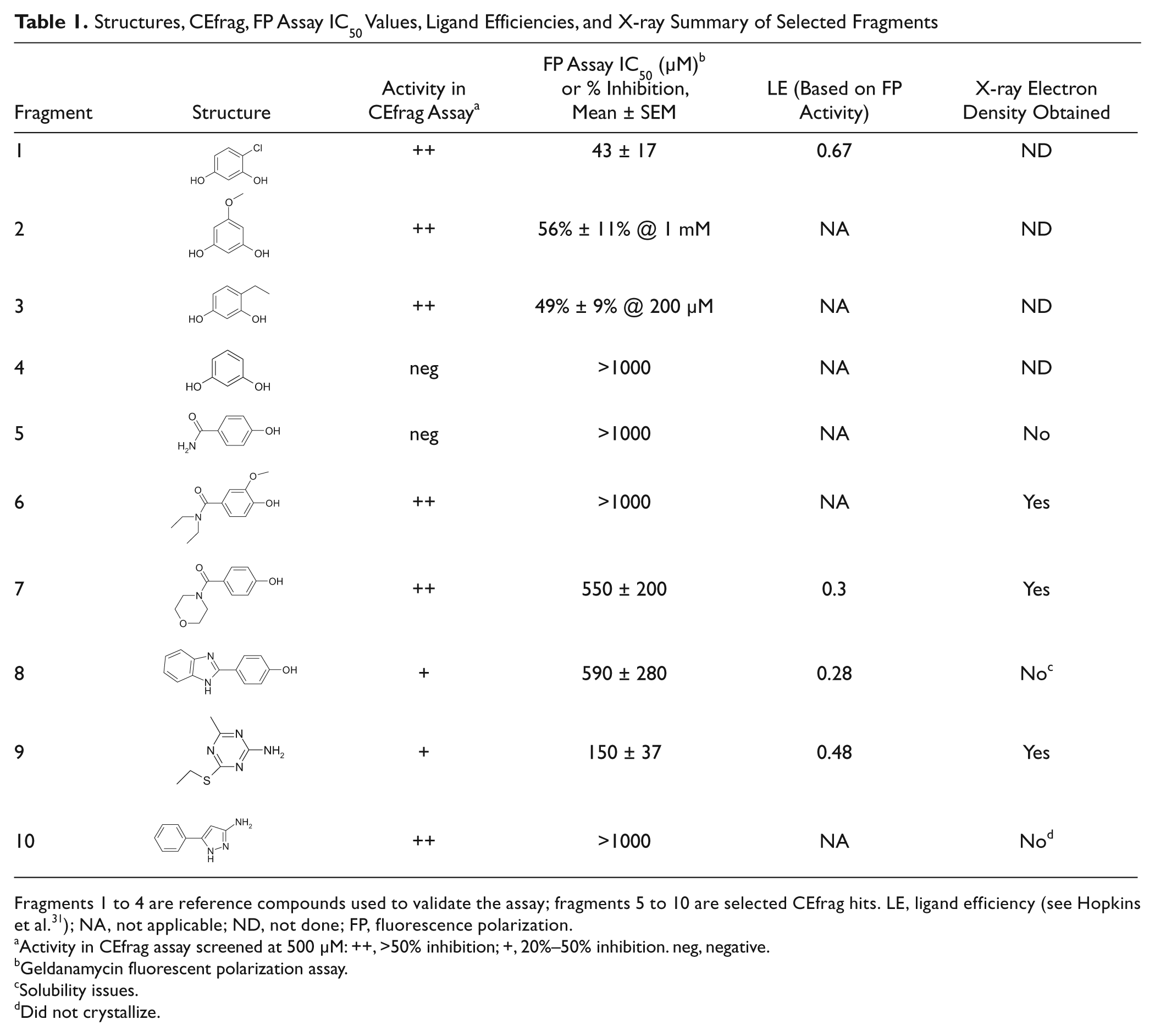
Fragments 1 to 4 are reference compounds used to validate the assay; fragments 5 to 10 are selected CEfrag hits. LE, ligand efficiency (see Hopkins et al. 31 ); NA, not applicable; ND, not done; FP, fluorescence polarization.
Activity in CEfrag assay screened at 500 µM: ++, >50% inhibition; +, 20%–50% inhibition. neg, negative.
Geldanamycin fluorescent polarization assay.
Solubility issues.
Did not crystallize.
Screening of a subset of the Selcia fragment library (609 compounds) at 500 µM using the established Hsp90 CEfrag assay identified 42 compounds that reproducibly inhibited (≥20% inhibition) the radicicol mobility shift in the presence of full-length Hsp90 (i.e., a confirmed hit rate of 6.9%). The Z factor for the assay was 0.6. The use of the four-capillary Gemini instruments 20 enabled a throughput of 100 samples/instrument/d.
Hit Confirmation by FP Assay and X-ray Crystallography
The FP assay detected 12 of the confirmed CEfrag hits with inhibition >15% at 500 µM, and of these, 5 had measurable IC50 values in the FP assay (3 of these are shown in Table 1 : fragment 7, fragment 8 [4-(1H-benzoimidazol-2-yl)-phenol], and fragment 9). To validate these hits further, fragments 7, 8, and 9, as well as two strong CE hits that were not confirmed as actives in the FP assay (fragment 6 and fragment 10 [5-phenyl-1H-pyrazol-3-ylamine]) and one compound that was negative in both the CE and FP assays (fragment 5), were selected for X-ray crystallography. Not surprisingly, fragment 5 (4-hydroxy-benzamide), which was negative in the CE and FP assays, did not show any evidence for binding by X-ray. Of the fragments active in both assays, fragments 7 and 9 were shown by X-ray to bind to the adenosine triphosphate (ATP) binding site of Hsp90 ( Figs. 2 and 3 ), but no ligand density was observed for fragment 8. This may be due to the compound solubility as precipitation was observed in the crystal drop. The real power and sensitivity of CEfrag are illustrated with fragment 6, which, despite being a strong CE hit, was too weak to be detected by the FP assay and yet was still confirmed by X-ray to bind to the ATP binding site of Hsp90.
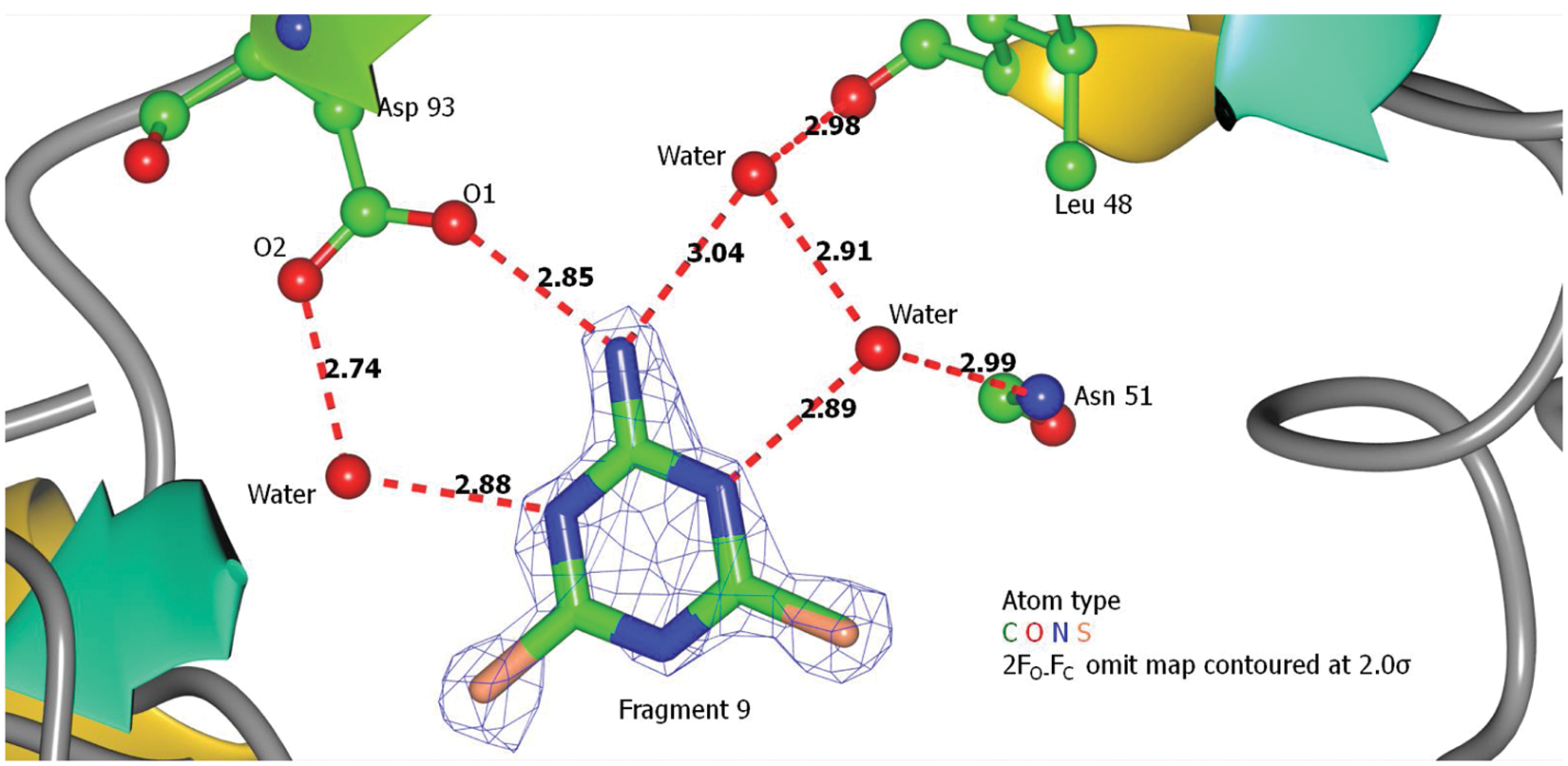
Binding of fragment 9 to Hsp90N. Ligand protein interactions for fragment 9 are predominantly hydrogen bonding, one direct and three water mediated. The interactions involved the fragment’s nitrogen atoms and residues Leu48, Asn51, and Asp93. The amino group of fragment 9 formed a direct hydrogen bond with O1 of Asp93. Three other water-mediated indirect hydrogen bonds were connected to O2 of Asp93, Nδ2 of the Asn51 side chain, and the carbonyl oxygen of Leu48. Electron density is not complete for fragment 9 with the ethyl tail missing. The sulfur atom and the ethyl tail could be on either side of the triazine ring. Numbers next to each red dashed line indicate bond length in Angstroms.
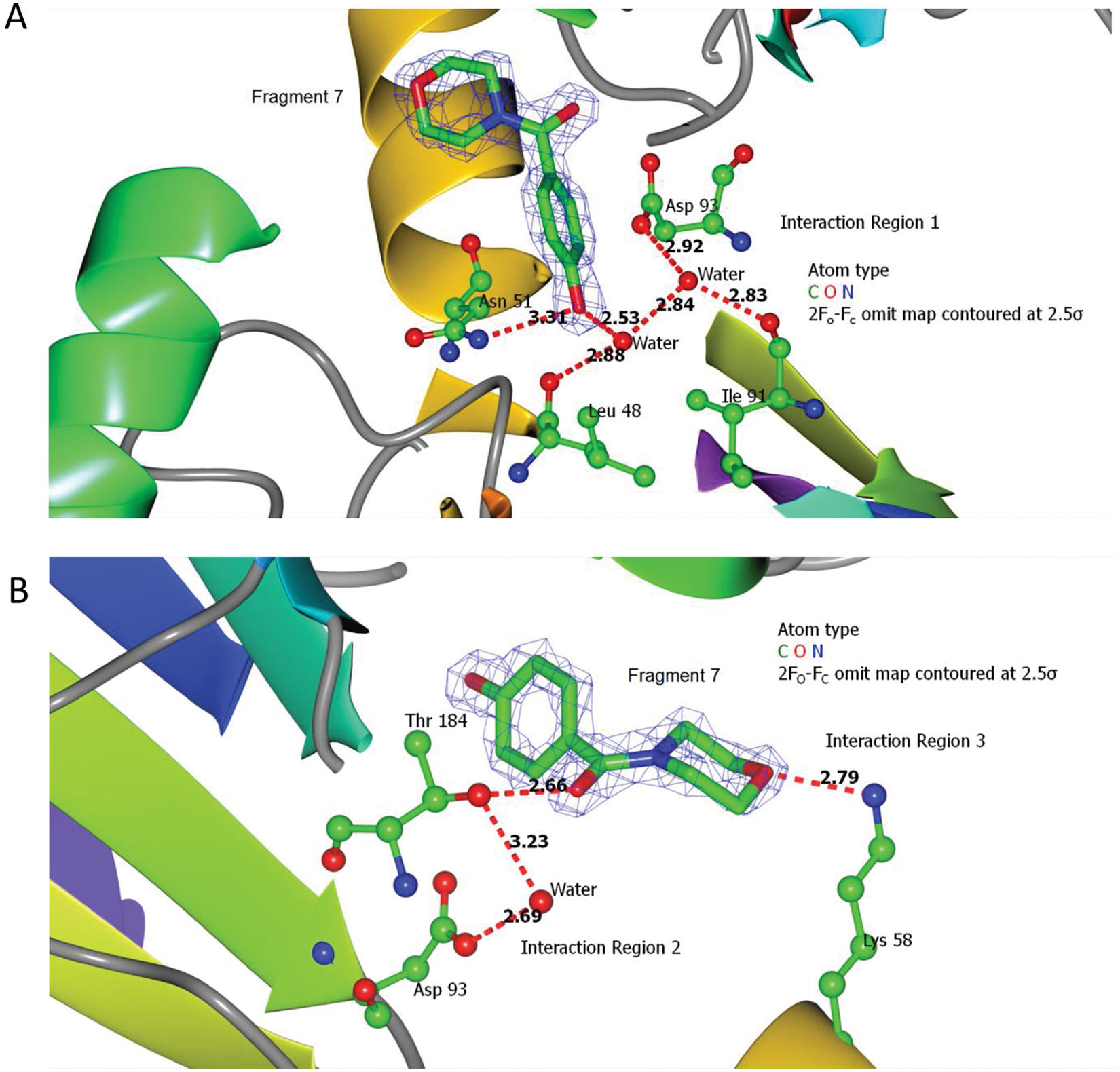
X-ray data: binding of fragment 7 to Hsp90N. (
Structure Analysis
The purified Hsp90N protein was crystallized with and without bound ligand; the crystals showed strong diffraction, enabling collection of full data sets to 1.7Å resolution in 24 h using the sealed tube source. Data collection and refinement statistics are included in Table 2 for apoprotein and complexes produced with radicicol (figures not shown) and three fragments whose solubility allowed co-crystallization ( Figs. 2 and 3 ).
Crystallographic Statistics of Hsp90N Structures
The protein domain structure was very similar to that described by previous investigations showing an extensive eight-stranded β-sheet with one parallel strand toward the C-terminus. Seven regions of extended α-helix reside on one side of this sheet, and the ATP binding pocket is located in a cleft between helices with the mainly hydrophobic sheet side chains as its floor. One side of the pocket is made up of a long helix (residues 41–61), whereas the other comprises interhelix regions. The pocket is filled with well-ordered water molecules hydrogen bonded to the protein and to each other. The adenine ring of ATP when bound is inclined within the pocket, interacting with Asp93 beneath the long helix, whereas the ribose and phosphate groups lead out to the domain surface. Ligands are partially enclosed in the pocket by Lys58.
Binding of radicicol with our recombinant human Hsp90α N-terminal ATPase domain solved at 1.69Å was consistent with that published for radicicol binding to yeast 28 and dog Hsp90 29 (see Table 2 for crystallographic statistics; figure not shown). Fragments 6, 7, and 9 were all shown to interact at the ATP binding site. The data are not shown for fragment 6 as this has previously been published by Murray et al. 11 ; our results were consistent with those published. Fragment 7, which comprises morpholine and phenolic rings coupled through a carbonyl carbon, interacts at three major ligand regions with the protein, involving direct residue-ligand and indirect hydrogen bonding mediated by water molecules ( Fig. 3 ). Interaction of the morpholine ring (interaction region 3; Fig. 3 ), which is the main chemical difference between fragments 6 and 7, is a single charge-assisted hydrogen bond between the oxygen atom of the morpholine ring and the amino group of the Lys58 side chain.
Fragment 9, a modified triazine compound, formed H bonds directly with Asp93 and indirectly through water molecules also with Asn51 and Leu48. Fragment 9 only produced crystals when present in a molar excess of 1.75:1 (see Fig. 2 ). Greater quantities produced no crystals, suggesting that a lower affinity site may be present on the protein surface that interferes with packing. Rearrangement of the polypeptide chain between residues 105 and 114 was observed with this ligand ( Fig. 4 ) compared with the structure obtained with fragments 6 and 7.
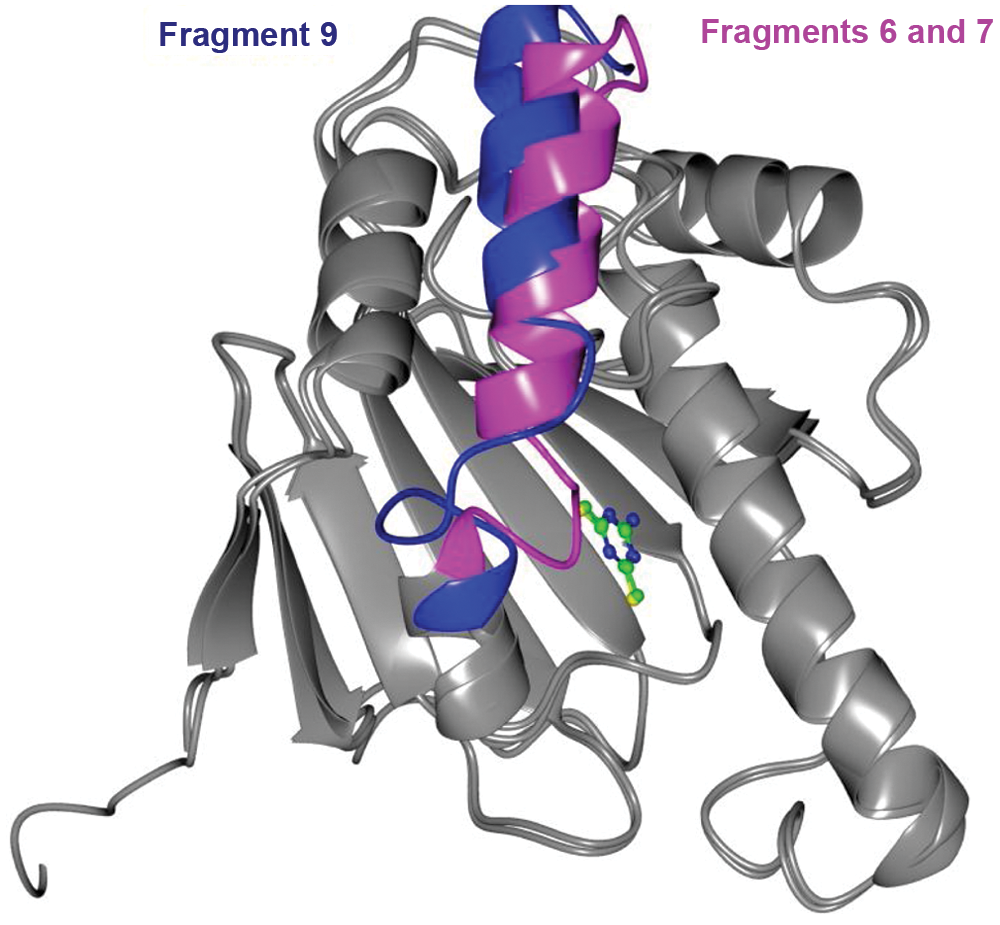
Structural change caused by fragment 9. Protein structure of Hsp90α ATPase domain bound to fragments 6 and 7 was superposed to its structure when bound to fragment 9. Both structures were obtained from crystals that were grown under identical conditions. The structural difference is highlighted as shown. A dramatic loop rearrangement spans from residues 105 to 114, and a minor translation of the helix is also observed.
Discussion
CEfrag has been developed as a novel fragment screening technique able to detect the weak affinity interactions often attributed to fragments binding to their targets. In this study, we selected Hsp90 as a well-known target that has previously been subjected to numerous fragment screening campaigns by others.11–15 The availability of these publications assists with validation of the hits obtained by CEfrag.
The assay was validated using a selection of resorcinol fragments that are well-known inhibitors of Hsp90 at the ATP binding site. 27 CEfrag readily detected inhibitors in the high µM to mM range; fragment 2 has previously been shown to have an IC50 of 1.2 mM. 27 The hit rate obtained using the Selcia fragment library and the Hsp90 CEfrag assay (6.9%) was consistent with that obtained by others (6.1%) using NMR and the Vernalis fragment library, 30 thus confirming that Hsp90 is a druggable target and highlighting that CEfrag is a technique capable of detecting weak affinities.
We have demonstrated here that CEfrag is a powerful technology for fragment-based drug discovery, especially in combination with an orthogonal assay and structural biology. The majority of the hit structures detected by CEfrag were amino heterocyclic compounds that are consistent with known Hsp90 inhibitors,12,13,27 and we have confirmed the binding mode of some of these fragments by X-ray crystallography. It is noteworthy that the competitor probe ligand itself does not need to be a charged species as illustrated by the fact that radicicol is a neutral molecule. The majority of the CEfrag hit compounds detected by the orthogonal FP assay showed very low activity in this assay (i.e., IC50 >500 µM), underlining the sensitivity of the CEfrag technique.
As the FP assay did not detect all the fragment hits identified by CE, it is hypothesized that the majority of the CEfrag hits were too weak to be detected by FP. The lack of detection of these CE hits in the FP assay is unlikely to be due to aggregation or BSA binding as the FP assay was repeated in the absence of detergent and BSA and their activity was still not detected (results not shown). When testing high concentrations of fragments, biochemical assays based on fluorescence polarization often suffer from interference by intrinsic fluorescence, thus rendering them less sensitive than biophysical assays. This is illustrated by the CEfrag hit, fragment 6, which is a known drug (ethamivan) and was previously demonstrated by Murray et al. 11 to be a weak Hsp90 binding fragment of 790 µM affinity as determined by isothermal calorimetry; our FP assay was not able to detect the fragment’s activity, but we demonstrated binding by X-ray crystallography consistent with that previously shown by the Astex group (PDB ID: 2XDL). Interestingly, 16 of the CEfrag hits, instead of showing inhibition in the FP assay, showed >15% apparent stimulation of FITC-geldanamycin binding to Hsp90; this may be due to interference in the FP assay, masking their real affinity for Hsp90. The IC50 of radicicol in our FP assay was 40 ± 15 nM, which is consistent with that obtained by other techniques (Kd = 19 nM 28 ).
The triazine compound (fragment 9) was the most potent compound identified from the screen with a good ligand efficiency (LE 0.48) 31 ; it is very similar to a compound identified as an Hsp90 binder by the Vernalis Plc group using NMR, 30 where they showed a corresponding S-methyl compound had a measured IC50 of 350 µM. After fragment growth and optimization, a triazine-originated orally active compound is now in clinical trials.
The type of assay described here has wide-ranging applications with certain advantages compared with the currently used approaches for fragment screening in terms of throughput, protein consumption, and speed of assay development. It is capable of detecting in solution the interactions between two or more biomolecules as well as disruptions of such interactions by small molecules. Even weak interactions (often the case with proteins) can be detected with great sensitivity. The technique is highly flexible, offering the possibility to monitor one or several interacting molecules (biological macromolecules as well as small-molecule ligands). Moreover, the assay involves a separation step and does not monitor the protein, eliminating the need for highly pure target protein.
In summary, we have demonstrated that CEfrag is a new method for fragment-based drug discovery and highlighted some of the strengths and advantages of capillary electrophoresis in biomolecular fragment screening.
Footnotes
Acknowledgements
The authors thank Chris Swain for help with designing the Selcia fragment library, the Selcia medicinal chemists for review of the fragment library compounds, and in particular Dave Scowen, Andy Keats, Jussi Kangasmetsa, Ian Marshall, and David O’Brien for help with the fragment library quality control. Thanks also go to Hans Fliri and Bob Carling for critical reading of the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
WenJie Chen is a recipient of an IMPACT PhD studentship grant code HN9A, which is sponsored by Selcia Ltd. The other authors received no financial support for the research, authorship, and/or publication of this article.