
Abstract
Small-molecule compounds (SMCs) can provide an inexpensive and selective approach to modifying biological responses. High-content analysis (HCA) of SMC libraries can help identify candidate molecules that inhibit or activate cellular responses. In particular, regulation of cell death has important implications for many pathological conditions. Dependence receptors are a new classification of proapoptotic membrane receptors that, unlike classic death receptors, initiate apoptotic signals in the absence of their ligands. EphA4 has recently been identified as a dependence receptor that may have important functions in conditions as disparate as cancer biology and CNS injury and disease. To screen potential candidate SMCs that inhibit or activate EphA4-induced cell death, HCA of an SMC library was performed using stable EphA4-expressing NIH 3T3 cells. Our results describe a high-content method for screening dependence receptor-signaling pathways and demonstrate that several candidate SMCs can inhibit EphA4-mediated cell death.
Introduction
An improved understanding of cellular responses is critical for developing successful therapeutic strategies for human clinical trials. Several therapeutic strategies target cell death, including neuroprotective antiapoptotic approaches following traumatic CNS injury 1 and proapoptotic mechanisms in neoplasia. 2 Apoptotic cell death has been shown to result from a variety of stimuli, such as absence of survival factors (deprivation response)3,4 or ligand receptor–induced apoptosis (instructive response). 5 Members of the tumor necrosis factor receptor (TNFR) gene superfamily, such as TNFR1, Fas, and DR4/5, are well described in their ability to initiate caspase-dependent cell death following ligand activation. 6 Recently, a novel mechanism of apoptotic cell death was described for a family of receptors, termed dependence receptors, in which caspase-mediated cell death is initiated in the absence of ligand activation. 7 Alternatively, when dependence receptors are bound to their ligand, they transduce “positive” signals that elicit biological processes such as survival, differentiation, and/or migratory responses. The netrin receptors DCC and Unc5H1-3,8,9 p75NTR,10 the androgen receptor, 11 Patched, 12 αVβ3 integrin, APP, 7 MET, 13 TrkC, 14 and Neogenin/RGM, 15 have all been shown to have dependence receptor functions. Furthermore, we recently showed that the EphA4 receptor, which functions to regulate neuronal progenitor numbers during adult neurogenesis, is a new member of the dependence receptor family. In the absence of its ligand ephrinB3, EphA4 undergoes intracellular cleavage and activates caspase-dependent cell death, which can be blocked by restoring the ligand. 16
In cancer biology, dependence receptors are recognized as potential tumor suppressors, 17 and Eph receptors have been shown to regulate cancer cell death. 18 In these cells, activation of dependence receptor signals would be a potential target for reducing growth. Conversely, following disease or injury, cell loss is deleterious to recovery, and blocking dependence receptor signals might promote recovery. 19 This is most evident in CNS injury, after which neurons and glia are highly susceptible to cell death that is both acute and progressive. Unfortunately, few agents have been identified to adequately block cell death following CNS injury; therefore, we sought to identify new compounds that might inhibit dependence receptor-mediated cell death. Targeted therapies typically employ one of three strategies: the use of function blocking monoclonal antibodies (mAbs), antisense oligonucleotides (AONs), or small-molecule compounds (SMCs). We chose to test SMCs because they have therapeutic advantages over mAbs and AONs, such as increased stability, potential for oral delivery, moderate cost, and available for many defined targets. In particular, many SMCs are efficient at crossing the blood-brain barrier and plasma membranes and have been previously developed to target multiple cellular processes such as growth factor/receptor responses, signal transduction pathways, cell cycle control, and cell death. Here we describe a simplified cellular modeling system that uses high-content analysis (HCA) of NIH 3T3 cells expressing EphA4 to assess the ability of SMCs to modulate EphA4-mediated cell death signals. In addition, we have developed a reliable EphA4 ligand (i.e., the ectodomain of ephrinB3 protein) as a way to block EphA4-mediated cell death, which was used as a control to compare with SMC candidates.
Materials and Methods
Establishment of EphA4-V5 and V5 NIH 3T3 Stable Cell Lines
Stable or transient transfected NIH 3T3 cell lines expressing EphA4-V5 (EphA4-3T3) or V5 alone (vector-3T3) were generated by subcloning the murine EphA4 into the pcDNA3.1-V5 vector (Life Technologies, Carlsbad, CA). This construct, which encodes the full-length mouse EphA4 receptor with a C-terminal V5 epitope, was described elsewhere. 16 The construct was transfected into NIH 3T3 cells using Lipofectamine 2000 (Life Technologies) according to the manufacturer’s instructions. Clonal cell lines were derived from a nonclonal cell population using single-cell dilution cloning and cultured in Dulbecco’s modified Eagle’s medium (DMEM)/F12 medium (Life Technologies), supplemented with 10% fetal bovine serum (FBS; Hyclone, Logan, UT), 100 U/mL penicillin, 100 µg/mL streptomycin (Penicillin-Streptomycin; Life Technologies), and 800 µg/mL G418 (Geneticin; Life Technologies) for selection. Immunostaining against the V5 epitope was employed to demonstrate effectiveness of the transfection, and cells were routinely cultured in 100-mm culture plates at 37 °C and 5% CO2 at subconfluency. Cells were harvested every 2 to 3 days following 0.05% w/v Trypsin (Life Technologies) treatment and reseeded at a density of 1 X 106 cells/dish (100-mm plate) in DMEM/F12 medium containing G418.
Protein Preparation and Immunoblotting
Crude membrane/cytosolic preparations were extracted from stable cell lines. At 80% confluency, vector-3T3 or EphA4-3T3 cells were stimulated with either ephrinB1 or ephrinB3 aggregates at 1 µg/mL concentration for 1 h. The cells were then rinsed with phosphate-buffered saline (PBS) and were scraped off in the presence of 1× TENN buffer (50 mm Tris [pH 7.4], 5 mM EDTA, 150 mM NaCl) containing 1% Triton X-100 and a cocktail of protease (Complete; Roche, Basel, Switzerland) and phosphatase (1 mM sodium orthovanadate, 2 mM sodium pyrophosphate) inhibitors. The cells were homogenized 50 times with a glass Dounce homogenizer (Wheaton, Millville, NJ) and then sonicated (5 Watts [RMS] for 5 s on a Misonix Microson ultrasonic cell disruptor; Misonix, Farmingdale, NY) to disrupt cell membrane integrity. The homogenate was centrifuged at 12 000 rpm for 15 min at 4 °C to pellet cytoskeletal components. The supernatant was transferred to a new tube, and the protein concentration was quantified using the Lowry protein assay (Bio-Rad, Hercules, CA) against a bovine serum albumin (BSA) standard curve. Then, 10 µg of proteins were incubated with 0.1 µg of antibody (anti-V5 [R960-25] from Life Technologies or anti-EphA4 [sc-921] from Santa Cruz Biotechnology, Santa Cruz, CA) in a 500-µL volume of TENN buffer, and tubes were rotated overnight at 4 °C. Then, 25 µL of Protein G Plus-Agarose beads (Calbiochem, San Diego, CA) were added to each tube for at least 3 h at 4 °C. The beads were spun down at 12 000 rpm for 1 min, the supernatant was removed, and the pellet was washed twice in 500 µL PBS followed by two washes in 500 µL PBST (PBS with 1% Triton X-100). The protein-antibody-bead complexes were dissociated by boiling for 5 min in 25 µL sample loading buffer, and the proteins were then loaded onto a 7.5% polyacrylamide gel. After electrophoresis, the proteins were transferred to a nitrocellulose membrane, which was blocked in 5% nonfat milk/TBST or BSA solution and probed with the primary antibodies (anti-V5, anti-EphA4, and anti-phosphotyrosine from BD Biosciences, San Jose, CA [61000]). After several TBST washes, the membranes were probed with the appropriate horseradish peroxidase (HRP)–conjugated secondary antibodies (anti-rabbit [18-8816-33], anti-goat [18-8814-33], anti-mouse [18-8817-33]; eBioscience, San Diego, CA) and washed several times in TBST, and the proteins were visualized on X-ray film (MidSci blue autoradiography film; MidSci, St. Louis, MO) using enhanced chemiluminescence detection (SuperSignal West Pico solution; Thermo Scientific, Waltham, MA).
Immunostaining
EphA4-3T3 cells were plated at 5 × 103 cells/well on 13-mm round coverslips in 24-well plates (Falcon, BD Biosciences, Bedford, MA) on round coverslips, and they were incubated at 37 °C and 5% CO2 for 24 h in DMEM/F12 supplemented with 10% FBS. After this period, they were further incubated for another 24 h in DMEM/F12 and 10% FBS in the presence or absence of 1 µg/mL preclustered ephrinB3-Fc (395-EB-200; R&D Systems, Minneapolis, MN) or ectodomain (ed)–ephrinB3. Cells were then fixed in 4% paraformaldehyde (PFA) for 10 min at room temperature (RT), washed twice with PBS, and then permeabilized with 0.4% Triton-X100 for 10 min. The wells were blocked with 5% BSA for 30 min, after which they were treated with 1 µg/mL mouse anti-V5 antibody (Life Technologies) diluted in PBS, followed by incubation with Alexa Fluor 488–conjugated goat anti-mouse antibodies (A-11001; Life Technologies) for 30 min, and 10 µg/mL Hoechst 33258 solution (Life Technologies) was then added for 10 min at RT. After one wash with PBS, the coverslips were mounted on Snowcoat X-tra microslides (Leica, Buffalo Grove, IL), and images were acquired using a Zeiss Axiovert 200M microscope (Carl Zeiss, Oberkochen, Germany).
Generation of Soluble EphrinB1 and EphrinB3 Ectodomain Protein Fragments
The gene fragment coding for amino acids 26 to 236 of mouse ephrinB1 extracellular domain was amplified by PCR and cloned between the NcoI and EcoRI restriction sites of the pET32a vector (Novagen, Madison, WI) using the forward primer 5′CATCCCATGGATACGCCGTT GGCCAAGAAC and the reverse primer 5′GGCGAATT CTCACTTGGAGTTGAAGAAGCTGTC. The gene fragment coding for amino acids 28 to 227 of mouse ephrinB3 extracellular domain was amplified by PCR and cloned between the NcoI and EcoRI restriction sites of the pET32a vector (Novagen) using the forward primer 5′CATGCCATGG AACTCAGCCTGGAGCCTG and the reverse primer 5′GGCGAATTCTCACACTGCGGGCATGCTG. The proteins were expressed in Escherichia coli AD494 (DE3) pLysS strain (Novagen) and purified via sequential column chromatography. Briefly, cultures were grown at 37 °C with shaking at 225 rpm up to an OD600 = 0.6, and protein expression was induced with 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) for 16 h at 20 °C. Cells were harvested and lysed, and the nucleic acids from the crude extract were precipitated with 0.3% polyethylenimine (Sigma, St. Louis, MO). Proteins in the supernatant were precipitated with 0.8 M ammonium sulfate (AMS). The AMS pellet was resuspended in buffer A (20 mM Tris-HCl [pH 8.0], 150 mM NaCl, 10 mM imidazole, 10% v/v glycerol) and was run on a Ni2+ column (PorosMC; GE Healthcare, Piscataway, NJ) over a 10- to 500-mM imidazole gradient. The fractions from the Ni column containing the protein were pooled and run on an anion exchange column (MonoQ HR 10/10; GE Healthcare). The eluate was further run on a size exclusion column (Superdex 200 or 75; GE Healthcare) in a buffer containing 150 mM NaCl, 20 mM Tris-HCl (pH 7.5), 10% glycerol, 1 mM EDTA, and 1 mM dithiothreitol (DTT). The protein eluted in the void volume as soluble aggregates, and purity was confirmed by Coomassie staining after polyacrylamide gel electrophoresis.
Dynamic Light Scattering
Dynamic light-scattering (DLS) measurements of ectodomain ephrinB3 and B1 (ed-ephrins) were performed using a DynaPro-99-E-50 instrument (Wyatt Technology Corp., Santa Barbara, CA) in the batch mode. The data were collected at RT, with the laser power at 80% at a wavelength of 828.7 nm. A protein sample with a concentration of 0.4 mg/mL in a buffer containing 150 mM NaCl, 20 mM Tris-HCl (pH 7.5), 10% glycerol, and 1 mM EDTA was filtered through a 0.2-µm pore membrane; centrifuged for 15 min at 14 000 g; and then loaded into a 12-µL quartz cuvette. At least 10 measurements of scattered light intensity with a sum of squares (SOS) value less than 100 were recorded. Scattering data were analyzed using DYNAMICS V6 software version 6.3.40 (Wyatt Technology Corp.).
EphA4-Mediated Cell Death Assay and High-Content Analysis of SMC Libraries
To use HCA potential inhibitors/activators of EphA4-mediated cell death, we employed the following procedure. (1) EphA4-3T3 or vector-3T3 cells were plated at 103, 104, or 105 cells/well on 96-well plates (Falcon) and were incubated in DMEM/F12 (Life Technologies) supplemented with 10% FBS at 37 °C and 5% CO2 for 24 h. (2) The following day, the medium was changed to serum-free DMEM/F12 supplemented with 0.5, 5, or 50 µM SMCs from the ICCB Known Bioactives Library (Enzo Life Sciences, Farmingdale, NY) in the presence or absence of 1 µg/mL ed-ephrinB3. In the control wells, the medium was changed to DMEM/F12 only or DMEM/F12 supplemented with one of the following: 5% FBS, 1 µg/mL ed-ephrinB3 or ed-ephrinB1, or 1 µg/mL preclustered ephrinB3-Fc or ephrinB1-Fc (395-EB-200 and 473-EB-200, respectively; R&D Systems, Inc.). For the Fc reagents, clustering was performed with goat anti-human Fc antibodies (109-005-098; Jackson ImmunoResearch, West Grove, PA) (1:10 anti-Fc:ligand) for 1 h at 37 °C. The cells were further incubated for 24, 48, and 72 h at 37 °C and 5% CO2, after which they were treated with 10 µg/mL Hoechst 33258 solution and 5 µM Sytox Red (Life Technologies). (3) The live/dead cells were automatically counted using the Cellomics ArrayScan VTI Live HCS Reader (Thermo Scientific). The images were taken at a 5× magnification, and each well was divided into 49 fields, of which 3 were scanned and counted. The cells were stained for 10 min before imaging, and the chamber was set at 37 °C and 5% CO2.We found that a seeding concentration of 104 cells/well in 150 µL medium and an incubation time of 48 h after removing the FBS were the optimal conditions to produce at least 50% to 60% cell death.
Statistics
GraphPad’s InStat software was used for the Student two-tailed t test to compare two experimental groups (GraphPad Software, La Jolla, CA). One-way analysis of variance (ANOVA) on ranks was done for multiple comparisons. Differences were considered statistically significant if p-values were less than 0.05. Mean values were reported and graphed together with the standard error of mean. We used the formula Z′ = 1 – {(3 × positive control SD) – (3 × negative control SD)}/(positive control mean – negative control mean), where the positive control was EphA4-3T3 cells stimulated with ephrinB3 and the negative control was unstimulated EphA4-3T3 cells.
Results
Development of the Dependence Receptor Cell Death Model in Clonal Cell Lines
EphA4 receptors are newly defined dependence receptors that undergo intracellular cleavage that leads to caspase-mediated cell death (see Fig. 1A schematic). 16 To evaluate EphA4 dependence receptor functions, NIH 3T3 cell lines expressing V5-tagged EphA4 16 (EphA4-3T3) or control vector alone (vector-3T3) were generated and examined for expression and activation ( Fig. 1B - D ). A murine cell line model was chosen because it offers the possibility of further testing “hit” SMCs in primary cultures and in vivo wild-type or genetically modified murine models, as well as complex animal models of disease. In addition, human and mouse EphA4 have a high degree of homology (98.58% amino acid sequence identity). Immunostaining for the V5 tag showed strong expression of EphA4 in NIH 3T3 cells, and application of soluble ephrinB3-Fc ligands resulted in punctate EphA4 expression that likely reflects receptor clustering ( Fig. 1B ; see inset). To confirm cell surface expression of EphA4 in the stable cell line, we examined whether application of soluble ephrinB3 could induce EphA4 phosphorylation. Immunoprecipitation and Western blot analysis showed robust EphA4 protein expression in the EphA4-3T3 cell line and a significant enhancement in EphA4 phosphorylation upon ephrinB3-Fc stimulation ( Fig. 1C , D ). Following 48 h of serum withdrawal, we also observed a 21-kDa cleavage fragment in EphA4-3T3 cells that was attenuated following ephrinB3 stimulation or in the presence of the broad-caspase inhibitor z-VAD-fmk ( Fig. 1E ), similar to previous findings using transiently transfected HEK293T and immortalized neuronal 13.S.24 cells. 16
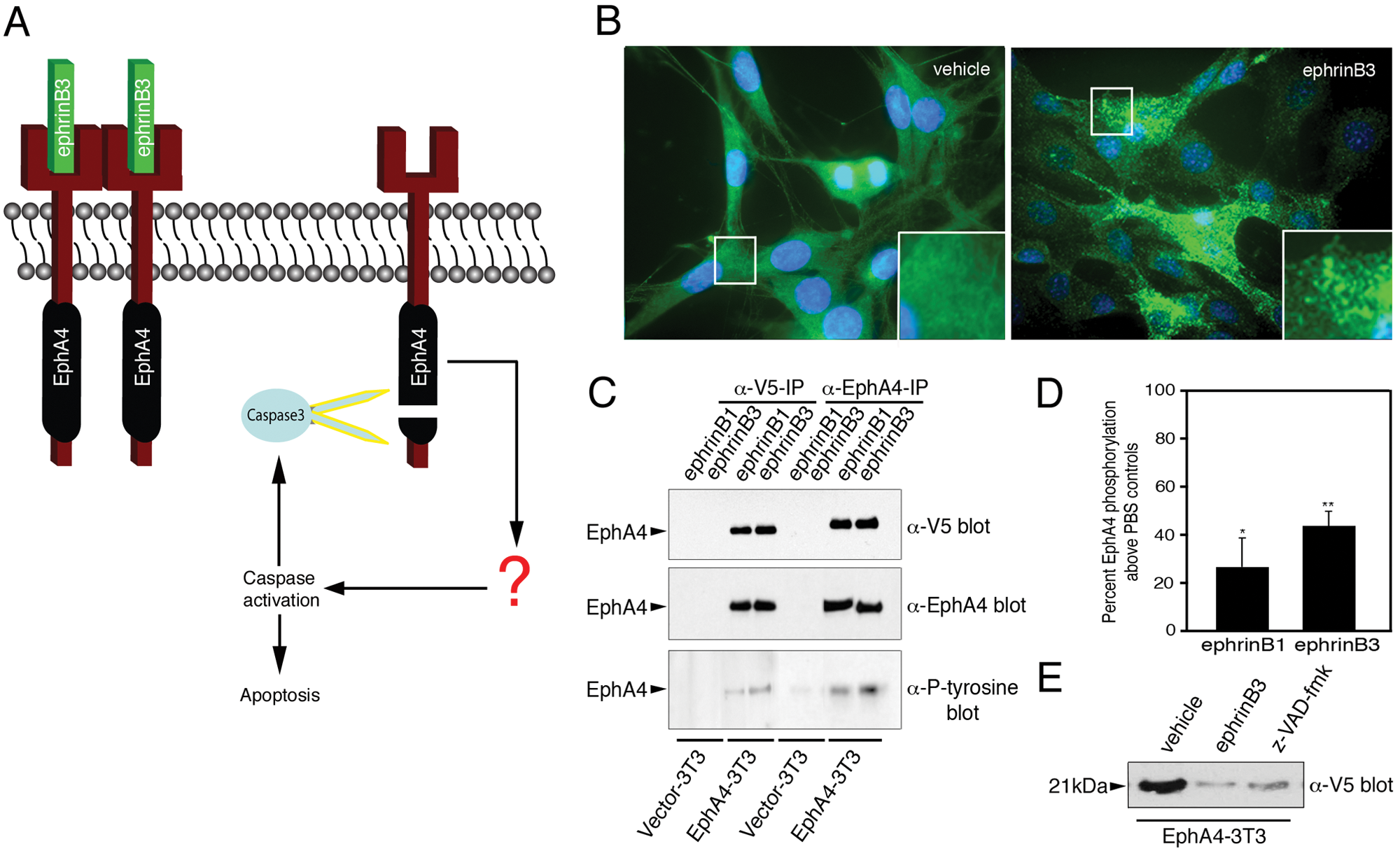
Modeling EphA4-mediated cell death. (
These studies support the use of stable EphA4-expressing cell lines for modeling EphA4-mediated cell death in the presence and absence of ephrinB3 stimulation.
Analysis of EphA4-Mediated Cell Death in a 96-Well Format
To screen the SMC library, vector-3T3 or EphA4-3T3 cells were seeded in 96-well plates in serum-containing medium. Outer wells of the 96-well plate were not used because of variability in medium evaporation that affected the cell death response. This phenomenon occurs commonly in 96-well plates and is known as the edge effect. 20 There are plates designed to overcome the edge effect (e.g., Nunc Edge 96-Well Plate; Thermo Scientific), but more consistent results were obtained using only the 60 inner wells of a 96-well plate.
After 24 h, the serum-containing medium was replaced with serum-free medium for 48 h to induce stress in the presence (positive control) or absence (negative control) of ephrinB3-Fc or SMCs ( Fig. 2A ). Cell survival was examined using a Sytox cell dead stain and compared with total Hoechst (blue) labeling ( Fig. 2B ) on the Cellomics ArrayScan VTI Live HCS Reader.
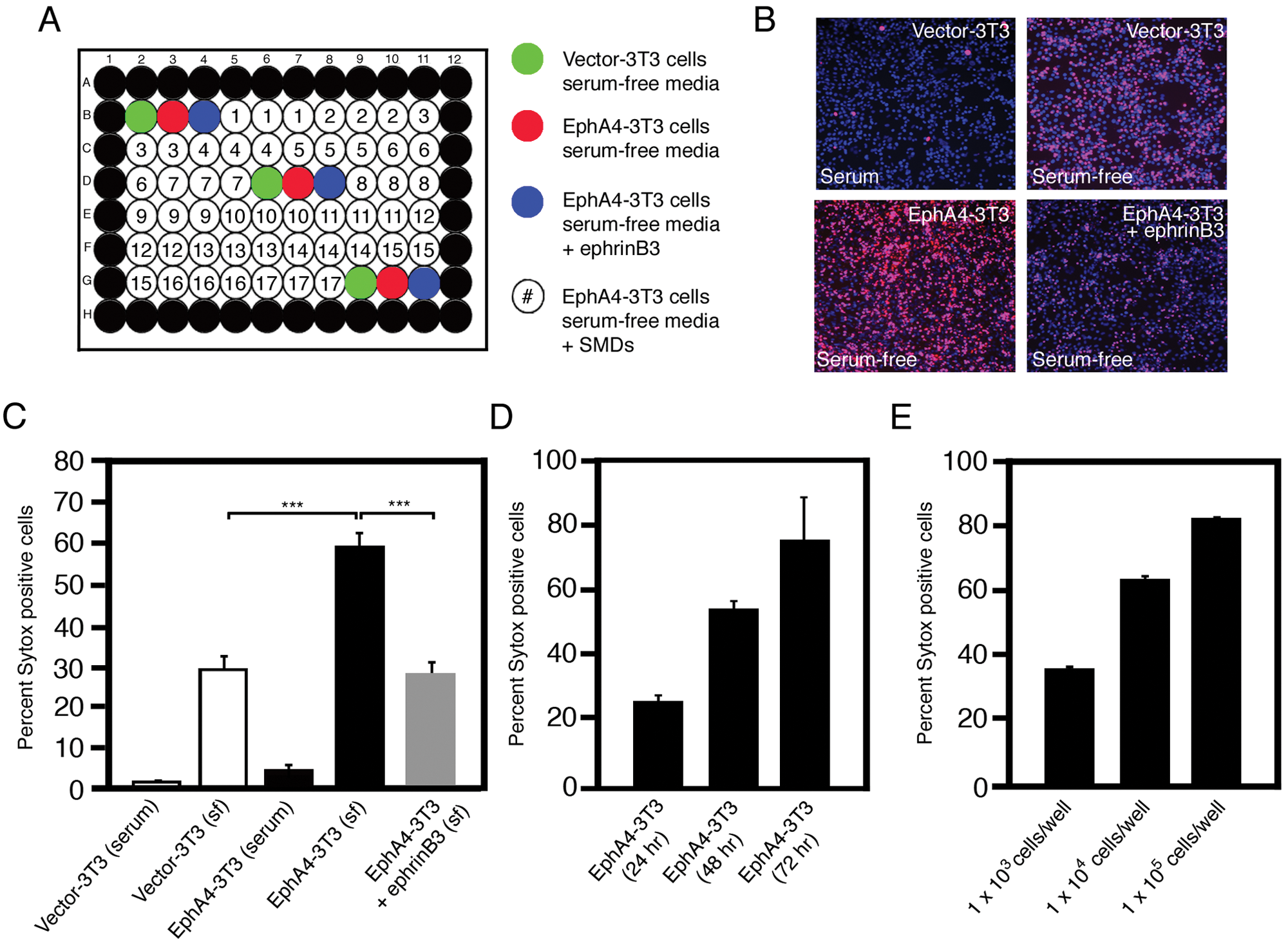
High-content analysis (HCA) in 96-well plates can be used to screen EphA4-mediated cell death, and ephrinB3 reduces cell death following serum deprivation. (
Vector-3T3 and EphA4-3T3 cells grown in serum showed little to no cell death; however, when these cells were grown in serum-free medium for 48 h, a significant increase in cell death was observed ( Fig. 2B , C ). Furthermore, the overexpression of EphA4 augmented cell death in the absence of serum, which was blocked by addition of ephrinB3-Fc ( Fig. 2B , C ). Cell death was consistent with our previous results obtained by using other methods for cell death detection such as trypan blue exclusion and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining. 16 The ability of ephrinB3 to block EphA4-mediated cell death further supports the dependence receptor functions for EphA4 and ephrinB3 in our assay. 16
In time course serum withdrawal experiments, we observed a proportional increase in the percentage of cell death with time and cell concentration, with the optimal incubation period for stress-induced cell death reaching 50% to 60% being 48 h, with an optimal seeding concentration of 1 × 104 cells/well ( Fig. 2D , E ).
We also evaluated whether clustered ephrinB3-Fc molecules block Eph-mediated cell death as compared with clustered ephrinB1-Fc, unclustered ephrinB3-Fc, and control Fc molecules. Eph receptor activation is known to require receptor aggregation to fully activate Eph signaling and subsequently inhibit dependence receptor-mediated cell death. 16 We found that application of preclustered ephrinB3-Fc molecules significantly reduced EphA4-mediated cell death by 60% to 75% at doses ranging from 0.5 to 2.0 µg/mL, respectively, as compared with clustered ephrinB1-Fc, unclustered ephrinB1 and ephrinB3, and Fc-only controls ( Fig. 3A ). No significant differences were observed among negative controls. The test for tolerance of cells to the SMC solvent showed that DMSO, which is typically used to dissolve many SMCs, altered cell survival in concentrations above 1%. However, in our experiments, the final DMSO concentration in the medium was between 0.3% and 0.5%, and we did not observe any toxic effects on cell survival (not shown). Finally, we showed that our assay was robust, with a calculated Z′ factor 21 of 0.58. Together, these findings suggest that EphA4 stable cell lines provide a good assay system for screening SMCs to identify potential inhibitors of cell death.
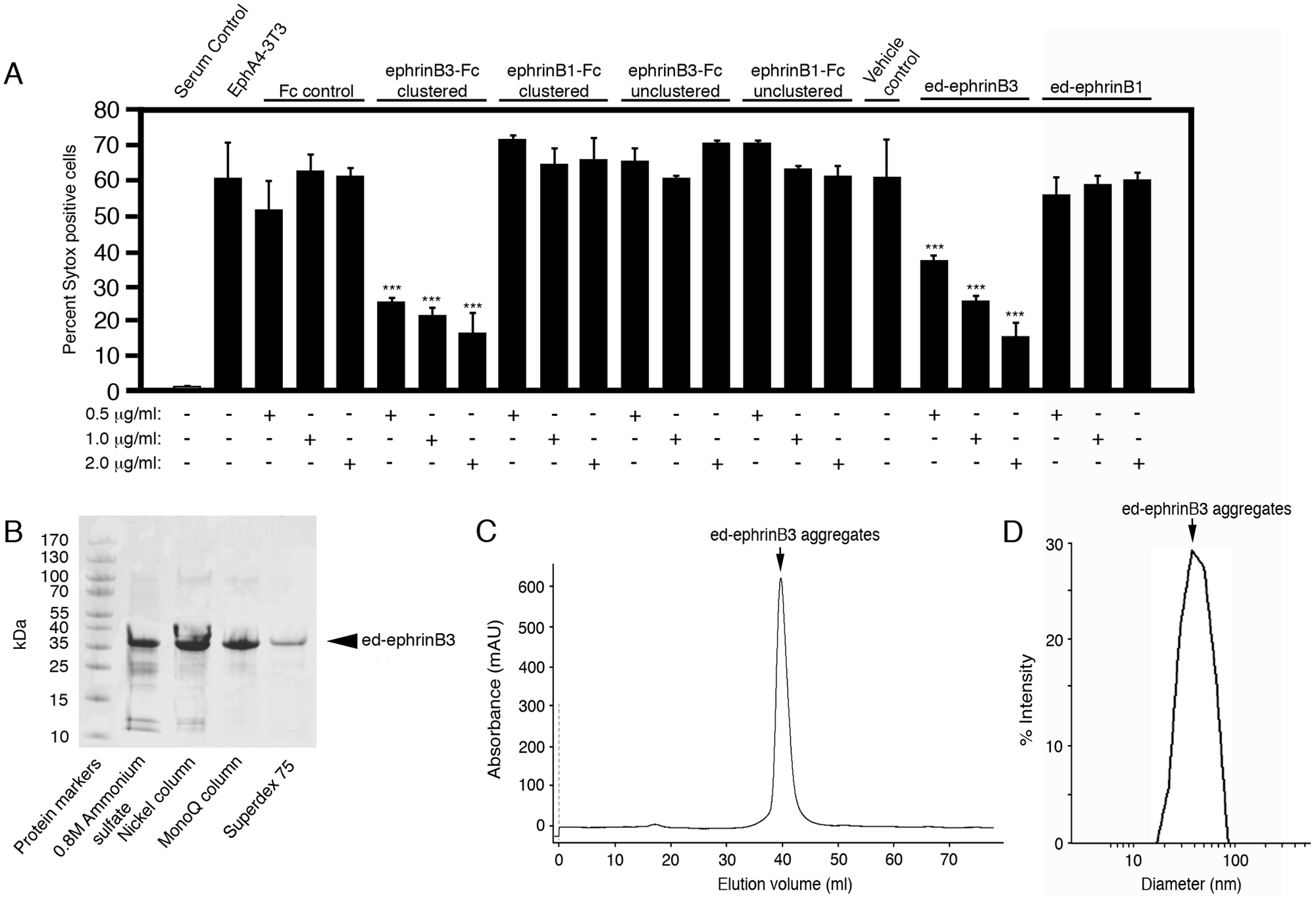
Development of ectodomain (ed)–ephrinB3 protein that self-aggregates and blocks EphA4-mediated cell death. (
Production of EphrinB1 and EphrinB3 Soluble Aggregates
The use of ephrin-Fc molecules to block Eph-mediated cell death has several disadvantages that include the expense of purchasing large quantities of ephrin-Fc, Fc controls and anti-human antibodies to cluster Fc-containing molecules, the variability resulting from the need to adequately cluster ephrin-Fc to generate biologically active molecules, and the requirement to maintain clustered molecules following delivery in animal models. To circumvent these problems, we generated soluble ephrinB1 and ephrinB3 ectodomain proteins (ed-ephrinB1 and ed-ephrinB3, respectively) that formed natural aggregates following purification by fast protein liquid chromatography (FPLC) ( Fig. 3B , C ). We successfully used these proteins to replace the ephrin-Fc molecules in our HCA assays. The purified proteins were of predicted size as revealed by denaturing gel electrophoresis analysis ( Fig. 3B for ed-ephrinB3; ed-ephrinB1 not shown). Both proteins eluted in the void volume fractions during size exclusion chromatography, with little to no monomers detected, suggesting that these proteins self-aggregate ( Fig. 3C ). The exclusion limit of the column that we used (Superdex 200; GE Healthcare) suggests that the size of these aggregates is at least 1300 kDa. To more precisely assess the molecular weight of the ed-ephrin protein aggregates, we conducted DLS experiments on these proteins. Consistent with gel-filtration data, purified ed-ephrinB3 protein shows a large average apparent hydrodynamic radius of 21 nm ( Fig. 3D ). This suggests a molecular weight of 4165 kDa, which would correspond to an average of 104 monomers per particle of protein aggregate, assuming a globular-shaped protein. Analysis of the ed-ephrinB3 aggregate reagents as compared with ephrinB3-Fc molecules showed a similar ability to inhibit EphA4-mediated cell death as compared with ephrinB3-Fc molecules ( Fig. 3A ). These findings demonstrate that ed-ephrinB3 may function as a simple, reliable, and cost-effective compound to block EphA4-mediated cell death; however, soluble ed-ephrinB3 would only be effective for Eph-mediated cell death and presumably would not inhibit other dependence receptors. For this reason, we used an HCA approach to identify intracellular signaling molecules that may function as common inhibitors of cell death, including but not necessarily restricted to Eph-mediated cell death.
Small-Molecule Screen Using the HCA Platform
Using an HCA approach, we analyzed an SMC bioactive library for compounds that might improve survival in EphA4-3T3 cells following serum deprivation. The ICCB Known Bioactives Library (Enzo Life Sciences) contains 480 biologically active compounds that include bioactive lipids, nuclear receptor ligands, ion channel ligands, protease inhibitors, kinase inhibitors and activators, guanine nucleotide binding protein-coupled receptor ligands, and signal transduction modulators. As a control for the EphA4 specificity, the SMCs were also examined on the control vector-3T3 cell line (not shown). Only compounds that significantly increased or decreased cell death of EphA4-3T3 but not on vector-3T3 cells were considered to have a specific effect on EphA4-mediated cell death. In evaluating the effects of SMCs on EphA4-3T3 survival, SMCs could potentially fall into one of four groups: (1) little to no effect; (2) significant change in survival in the absence of ed-ephrinB3, with no additive ed-ephrinB3 effect; (3) significant change in survival with an additive or enhanced ed-ephrinB3 effect; or (4) toxic. In Figure 4 , we show representative SMCs for classification groups 2, 3, and 4 as compared with control conditions in which EphA4-3T3 cells are not serum deprived (control), serum-deprived (vehicle) controls, and serum deprived plus 1 µg/mL ed-ephrinB3. Numerous SMCs showed little or no effect (not shown).
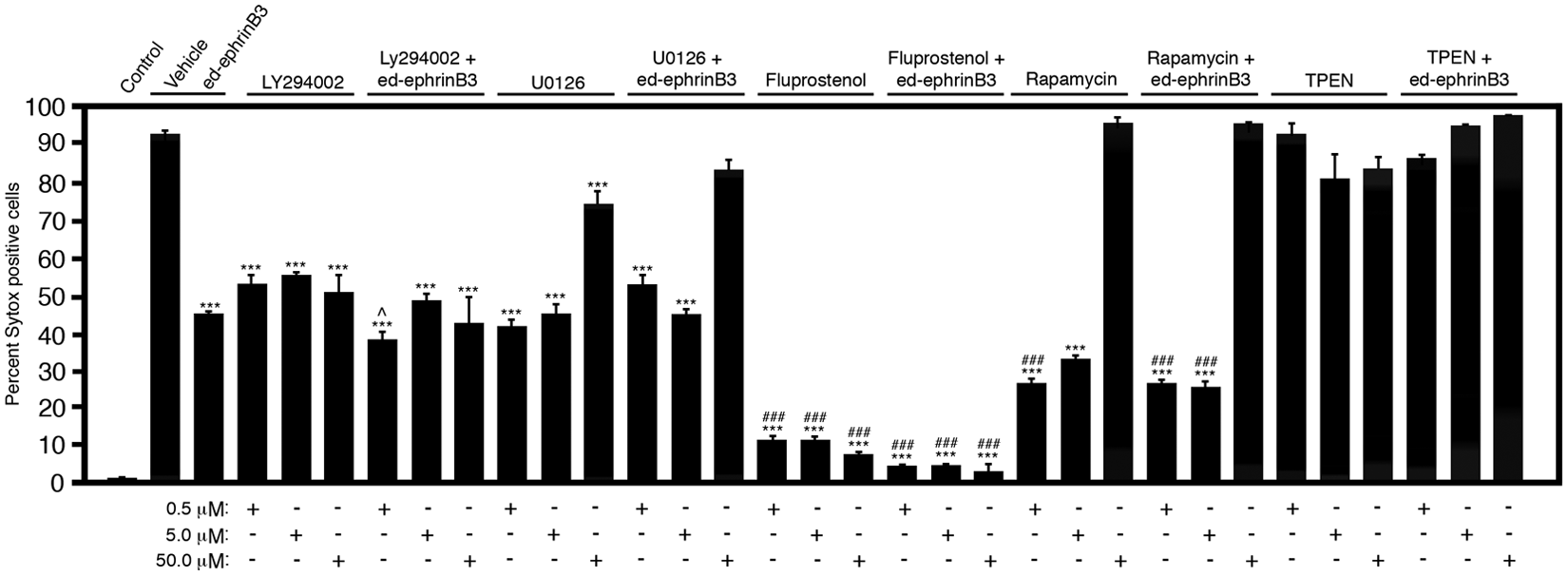
High-content analysis (HCA) of small-molecule compounds (SMCs) to block EphA4-mediated cell death in EphA4-3T3 cells. Representative SMCs can enhance cell survival in EphA4-3T3. The SMCs concentrations are shown at the bottom of the table. Significance is shown as the difference in survival between serum-free (sf) EphA4-3T3 cells (vehicle) and EphA4-3T3 cells treated with SMCs in the presence or absence of ed-ephrinB3 (***p < 0.001) or sf EphA4-3T3 cells (ed-ephrinB3) and EphA4-3T3 (SMCs) in the presence or absence of ed-ephrinB3 (###p < 0.001).
Figure 4 shows that application of ed-ephrinB3 reduced the number of Sytox-positive EphA4-3T3 cells by 50% following serum deprivation. Conversely, cell death of vector-3T3 cells following serum withdrawal remained virtually unchanged in the presence of ed-ephrinB3 (not shown). To identify potential cell death inhibitors, SMCs were added in the presence and absence of ed-ephrinB3. Most of the SMCs tested fell into group 1 (little to no effect), but a number of SMCs significantly reduced cell death comparable to the effect of ed-ephrinB3 ( Fig. 4 ). In particular, LY294002, a phosphatidylinositol 3-kinase (PI3K) inhibitor, improved EphA4-3T3 cell survival similar to ed-ephrinB3 alone. In addition, the presence of both LY294002 and ed-ephrinB3 showed an additive effect at a 0.5-µM concentration, suggesting that PI3K and Eph “dependence receptor” signaling may be independent of each other. Trends toward improved survival in the presence of ed-ephrinB3 were observed at higher concentrations but were not significantly different. U1026, a MEK (MAP/ERK kinase) inhibitor, showed similar improvements in cell survival with no additive effects at concentrations of 0.5 and 5.0 µM, but it became toxic at 50 µM. These results support the possibility that MAP/ERK kinase plays a role in Eph-mediated cell death. Fluprostenol, a prostaglandin F receptor agonist, showed significant improvements in cell survival at all concentrations tested in the presence or absence of ed-ephrinB3. Rapamycin, an mTOR (mammalian target of rapamycin) inhibitor, although toxic at the 50-µM concentration, improved cell survival at 0.5 and 5.0 µM more than ed-ephrinB3 alone, and no additive effects were observed in the presence of both rapamycin and ed-ephrinB3. Finally, TPEN (N,N,N′,N′-tetrakis-2-pyridylmethyl-ethylenediamine), an intracellular Zn2+ chelator, was toxic at all concentrations tested in both EphA4-3T3 and vector-3T3 cell lines, and addition of ed-ephrinB3 did not improve cell survival. Together, these studies support an HCA approach to screening inhibitors of EphA4-mediated cell death, as well as the potential involvement of MEK/ERK and mTOR in dependence receptor signaling cascades, but additional studies are needed.
Discussion
Dependence receptors have been implicated in various pathological conditions such as cancer, CNS injury, and disease, 22 but therapeutic targeting of dependence receptor mechanisms has presented numerous challenges for investigators. This is primarily because dependence receptors do not induce cell death following ligand-receptor interactions but, in fact, are cleaved and activated in the absence of their respective ligands. As a result, little is known about the signal transduction pathways that ultimately activate downstream caspases to induce cellular apoptosis. To study these responses, we created an HCA approach to screen large numbers of molecules that might inhibit or activate dependence receptor signaling.
For the present study, we chose to examine EphA4-mediated cell death, which is involved in numerous physiological and pathological processes, such as CNS development 23 ; cancer pathogenesis, including glioma 24 and melanoma 25 ; and adult neurogenesis. 16 This makes the EphA4 receptor a good target for drug therapies. Previous studies have identified small peptides 26 or small-molecule drugs27,28 that bind to the extracellular domain of EphA4 receptors and inhibit ephrin binding. Other compounds bind directly to the intracellular kinase domain and inhibit the tyrosine kinase receptor function of EphA4.29,30 However, we have recently shown that EphA4-mediated cell death activity is enhanced in the absence of ligand binding and is independent of its kinase activity. 16 In fact, EphA4-mediated cell death is a direct result of receptor cleavage of the intracellular EphA4 domain by caspase-3 or caspase-like molecules of the intracellular EphA4 domain, which leads to a unique signaling cascade involving caspase activation. Although common to many dependence receptors, this pathway remains poorly defined. Therefore, this study sought to develop an HCA screen to allow identification of downstream molecules that are involved in EphA4 signaling following cleavage by identifying SMCs that might interfere with or activate EphA4-mediated cell death. We chose to use a cell count assay based on Sytox Red to examine cell death, although alternative assays that measure total fluorescence/luminosity of live/dead cells (MTT Assay, Sigma; Cell Titer Glo Assay, Promega, Madison, WI) or caspase-activity (Fluorescent Caspase-3/7 Assay, Sigma) may also be possible. One advantage to employing fluorescence/luminosity assays is the simplicity and ability to use more common fluorescent plate readers as compared with the Cellomics ArrayScan VTI Live HCS Reader. Disadvantages of using fluorescence/luminosity live/dead assays include a relative analysis of cell death that requires a standard curve, low background, high signal-to-noise ratio, and highly consistent seeding. In contrast, the Cellomics ArrayScan VTI Live HCS Reader quantifies the ratio of live and dead cells to provide percent cell viability. Finally, employing caspase activity assays has the advantage of demonstrating selectivity for apoptosis but is more expensive for large HCA experiments.
Ligand replacement strategies are also a reasonable approach to blocking dependence receptor-mediated cell death; however, this strategy is largely confined to receptor-specific inhibition. For EphA4, ephrinB3 is one of several potential ligands, including A-class ephrins, which may bind and inhibit EphA4-mediated cell death. EphrinB3 also binds to EphB3, another potential dependence receptor in the Eph receptor family. 31 To inhibit EphA4- and EphB3-mediated cell death, we have developed and purified an ectodomain ephrinB3 (ed-ephrinB3) protein with binding/activating ability similar to commercially available ephrinB3-Fc molecules. The self-aggregating properties of this protein led to receptor activation and to reduced receptor cleavage and reduced cell death, properties that can be achieved only through antibody-mediated “preclustering” when using ephrinB3-Fc molecules. Ed-ephrinB3 provides a cheaper, simpler, and more reliable method for blocking EphA4-mediated cell death.
Another strategy to block or enhance Eph-mediated cell death is through the inhibition or activation of downstream signaling molecules, which may have an additional benefit of affecting multiple dependence receptor types. HCA provides a reasonable approach to identifying downstream mediators of dependence receptor-induced cell death. Our study represents a starting point by screening a small library of SMCs with known bioactivities on EphA4-mediated cell death to identify unique candidates that could be screened using other dependence receptors or libraries. Two protein inhibitors were identified that reduced cell death and showed no additive effects when applied with soluble ed-ephrinB3. In particular, the mTOR inhibitor rapamycin and the MAP/ERK (MEK) inhibitor, U1029, may reveal potential components of the EphA4-dependence receptor-signaling pathway. Rapamycin has already been shown to be an antiapoptotic agent by means of reducing the levels of activated caspase-9 and caspase-3, 32 two known potential effectors of the EphA4 dependence receptor pathway. 16 Similarly, MAP/ERK kinases are known to be activated by Ephs33,34 and do play roles in cell survival. These candidates will need to be confirmed and evaluated as downstream mediators of other dependence receptors. Identifying potential activators of EphA4-mediated cell death is more difficult since toxicity can complicate initial analysis. To overcome this limitation, parallel analysis of vector-3T3 control cell lines provides a tool to evaluate toxicity effects independent of EphA4-mediated cell death, where enhanced cell death in EphA4-3T3 but not vector-3T3 cells would support a pro-dependence receptor function and not toxicity. Together, these approaches may help function to identify novel inhibitors or activators of dependence receptor-induced cell death and provide a therapeutic strategy to block or enhance these effects in the setting of injury or cancer, respectively.
Footnotes
Acknowledgements
We thank Dr. Arun Malhotra for his help in purification of ed-ephrinBs. We also thank Dr. Pavan Vaidyanathan for critical reading of the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for research and/or authorship of this article: This work was supported by the Miami Project to Cure Paralysis, DOD W81XWH-05-1-0061 (DJL, VPL, JLB), NIH NS049545 (DJL), NS059866 (JLB), HD057632 (VPL), The Walter G. Ross Foundation (VPL), North Dade Medical Foundation (CMN), Christopher and Dana Reeve Foundation Fellowship Award (ER), and NINDS/NIH T32 NS007459 (ER).