
Abstract
Aberrant activation of the Wnt/β-catenin signaling pathway is associated with a wide range of human cancers. The interaction of β-catenin with T cell factor (Tcf) is a key step in activation of proliferative genes in this pathway. Interruption of this interaction would be a valuable strategy as a tumor therapy. In this study, we developed a novel fluorescein isothiocyanate (FITC)–labeled Tcf4-derived probe for identification of inhibitors of the β-catenin/Tcf4 interaction using a fluorescence polarization assay. This assay shows high potential for use in high-throughput screening for the discovery of inhibitors of the β-catenin/Tcf4 interaction.
Keywords
Introduction
The Wnt/β-catenin signaling pathway plays a critical role in embryonic development as well as in the growth of various cancers such as colorectal cancer.1,2 Normally, β-catenin is located predominantly at cell-cell junctions and at very low levels in both the cytoplasm and nucleus. Excess β-catenin usually undergoes GSK-3β/APC/axin (glycogen synthase kinase–3β/adenomatous polyposis coli tumor suppressor protein/axin) complex-mediated phosphorylation and subsequent proteasome-mediated degradation. 1 However, the binding of Wnt ligands to the surface receptor, Frizzled (Fzd), which activates Disheveled (Dsh), or the presence of certain mutations in APC, axin, or the β-catenin gene causes a cytoplasmic pool of β-catenin to accumulate. 1 This increase in cytoplasmic β-catenin results in its translocation to the nucleus, where it binds to members of the T cell factor/lymphoid enhancer factor (Tcf/Lef) family and stimulates transcription of Wnt target genes, including c-Myc and cyclin D1, which are critical for cell growth, proliferation, and differentiation.3,4 Consequently, interaction of β-catenin with Tcf4 is a key step in activating target genes for this pathway.4,5 Interruption of this interaction would be valuable as a tumor therapy. Current studies have already demonstrated that blockage of this interaction effectively inhibits in vitro colon cancer cell growth. 4 Seeking out novel agents that disrupt the β-catenin/Tcf interaction is clearly worthwhile. However, methods for screening large libraries of compounds library as potential small-molecule antagonists of the β-catenin/Tcf interaction first need to be developed. Several methods have been devised to detect the β-catenin/Tcf interaction or its mediated transcriptional activity. For instance, an enzyme-linked immunosorbent assay (ELISA)–based assay was developed to probe the binding between β-catenin and the N-terminal domain of Tcf4, 6 and a cell-based TOPflash/FOPflash assay was applied to screen compounds in libraries or to evaluate the inhibitory effects of candidate inhibitors on β-catenin/Tcf-mediated transcriptional activity. 1 However, these assays are not suitable for high-throughput screening (HTS) because they are too time-consuming and expensive. One possible technique is fluorescence polarization (FP), a powerful tool for studying receptor-ligand interaction that is widely used in HTS of small-molecule inhibitors. 7 In this article, we report the development of a new FP-based assay for testing the binding between β-catenin and Tcf4-derived peptide. This assay is simple, quick, and cost-effective and can be used for high-throughput drug screening.
Materials and Methods
Peptide Synthesis
All peptides were synthesized by solid-phase synthesis using a TentaGel (Sigma, St. Louis, MO) amide resin as the solid support 8 and a 9-fluorenylmethoxycarbonyl chemistry. A fivefold excess of Nα-Fmoc-amino acid, diisopropylcarbodiimide (DIC), and hydroxybenzotriazole (HOBt) was used in every coupling reaction step. Removal of the N-terminal Fmoc group was accomplished by 20% piperidine in dimethylformamide (DMF). The cleavage of a peptide from the resin was carried out by gentle stirring for 2 h at room temperature in a cleavage cocktail composed of water (5% [vol/vol]), thiophenol (5% [vol/vol]), and trifluoroacetic acid (TFA) (90% [vol/vol]). After precipitating with ice-cold diethyl ether and washing with ice-cold diethyl ether, the crude peptides were dissolved in 20% acetonitrile in water and then lyophilized. They were further dissolved in water and purified using semi-preparative reverse-phase high-performance liquid chromatography (HPLC). The purity of the final products was assessed by analytical reverse-phase HPLC and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. All peptides were at least 95% pure.
Peptides labeled with fluorescein isothiocyanate (FITC) (coupled to the NH2 group of the side chain of lysine) were prepared by linking the lysine side chain with 6-aminohexanoic acid at the C-terminal and protected with a (4,4-dimethyl-2,6-dioxocyclohex-1-ylidene)ethyl (Dde) group. The N-terminal amino acid was N-protected with tert-butyloxycarbonyl. At the end of the synthesis, the Dde group was removed from the peptide resin with 3% hydrazine in DMF (3 min at room temperature, three times). The resin was then washed with DMF and treated at room temperature with fluorescein isothiocyanate isomer I (5 eq) and diisopropylethylamine (DIEA) (10 eq) in DMF (minimum to cover the resin) overnight. When necessary, the coupling was repeated to obtain a negative Kaiser test. The resin was finally washed with DMF and dried with dichloromethane, cleaved, and purified as described above.
Protein Purification
Plasmid pET28a-FL-hβ-catenin (kindly provided byDr. Wenqing Xu, University of Washington), containing a full-length human β-catenin gene, was used to transform BL21(DE3)star (Invitrogen, Carlsbad, CA). After induction with 0.5 mM isopropyl-β-D-thiogalactopyranoside (IPTG), bacteria were harvested and lysed. The supernatant was loaded onto the affinity column containing Ni-NTA beads for His-tag fusion protein (QIAGEN, Valencia, CA). Purified protein was concentrated to 6 mg/mL in buffer containing 20 mM Tris-HCl, 100 mM NaCl, 2 mM dithiothreitol (DTT), and 20% glycerol and stored at –80 °C. The GST-Tcf4-1-54 fusion protein was prepared using a pGEX-6p-1-Tcf4-1-54 vector to transform the Escherichia coli strain BL21(DE3)star. The expression and purification procedure was similar to that used for β-catenin except that glutathione Sepharose 4B (GE Healthcare, Piscataway, NJ) was used for affinity chromatography, and the protein was concentrated to 1 mg/mL.
Fluorescence Polarization Assay
FP experiments were performed in black 96-well plates (Greiner Bio-One, Monroe, NC; Sigma, St. Louis, MO). The FP values were measured using a Synergy2 reader (BioTek, Winooski, VT) with excitation and emission wavelengths of 485/20 and 528/20 nm, respectively. For the saturation binding assay, 20 nM FITC-labeled Tcf4 8-30 peptide (tracer) was mixed with serial dilutions of β-catenin protein (1 nM to 10 µM) in 1× phosphate-buffered saline (PBS) buffer (containing 0.01% Triton X-100) at a final volume of 100 µL.
Competition experiments were carried out by mixing 20 nM tracer and 1.5 µM β-catenin protein (1.5–3 × Kd) 9 in the presence of different concentrations of competitors or vehicle DMSO. The plate was covered with adhesive aluminum foil and kept at room temperature for 3 h to reach equilibrium. 9 The data were expressed as relative binding compared with a nontreated control. Relative binding (% of control) = [(mPboud – mPfree) inhibitor/(mPboud – mPfree) control] × 100.
Pull-Down Assay
In total, 5 µg of GST-Tcf4-1-54 fusion protein was immobilized to glutathione Sepharose 4B resin, and 3 µg purified β-catenin was added in the absence or presence of 100 µM Tcf4 8-30 peptide. After overnight incubation at 4 °C, the bound protein was eluted with reduced glutathione and subjected to 7.5% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE).
Statistical Analysis
The statistical analysis was performed using the Student paired t test. Two-sided p values of <0.05 were considered statistically significant. Average values were expressed as mean ± SD.
Results and Discussion
Development of an FP Assay for β-Catenin/Tcf4
Fluorescence polarization assays have been widely used for studying protein-protein interactions. They have also been used in HTS for identification small-molecule agonists or antagonists of targets of interest. 10 However, to our best knowledge, the cell-free–based FP assay has not been developed for HTS of β-catenin/Tcf4 interaction inhibitors.
The β-catenin binding domain (CBD) of Tcf4 is located in the N-terminal 1-54 amino acids 5 and has a very high binding affinity for β-catenin. 11 On the basis of the crystal structures of β-catenin/Tcf4 complex and mutagenesis studies,5,12,13 we synthesized several FITC-labeled Tcf4 CBD-derived peptides and measured for their binding activity to β-catenin. One of these, corresponding to Tcf4 8-30 residues (designated as tracer in this article), retained the good capability to interact with β-catenin with a Kd value of 1 µM in the FP-based saturation binding assay. The binding reached equilibrium at 30 min and remained stable up to 24 h ( Fig. 1A ). Another two FITC-labeled peptides, TCf4 13-27 and 16-29, did not show high binding affinity with β-catenin (Kd >10 µM; data not shown). To confirm that the Tcf4 8-30 peptide specifically binds to the same pocket of β-catenin as its parental Tcf4 CBD does, we performed a pull-down assay to evaluate the binding activity of this peptide. As expected, the Tcf4 8-30 peptide was able to compete with GST-Tcf4 CBD for binding to β-catenin ( Fig. 1B ).
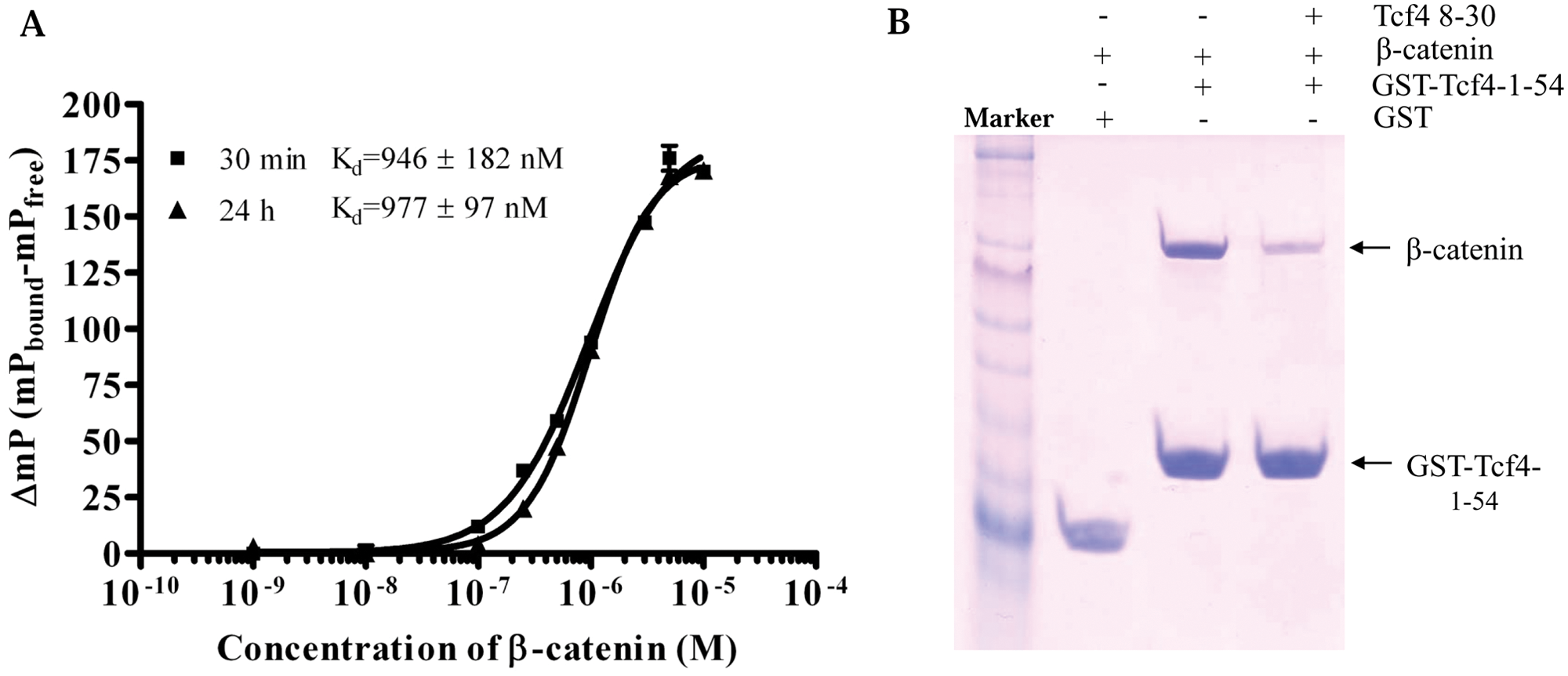
Tcf4 8-30 peptide specifically binds to β-catenin. (A) Saturation binding curve. Fluorescein isothiocyanate (FITC)–labeled Tcf4 8-30 peptide (tracer, 20 nM) was incubated with different amounts of β-catenin (1 nM to 10 µM). Fluorescence polarization values were recorded at 30 min and 24 h. Three separate experiments were performed and data were analyzed in Microsoft Excel (Microsoft, Redmond, WA) and plotted in Prism 4 (GraphPad Software, San Diego, CA). (B) Pull-down assay. Tcf4 8-30 peptide (100 µM) inhibits the β-catenin/Tcf4 interaction. The assay was performed three times, with one of representatives shown. M, marker.
The crystal structure of the β-catenin/Tcf4 complex revealed that Tcf4 residue Asp16 formed a hydrogen bond with β-catenin Lys435. The Glu17 of Tcf4, which was adjacent to Asp16, formed a salt bridge hydrogen bond with β-catenin Lys508. 5 Both residues were critical for the β-catenin/Tcf4 binding. Fasolini et al. 11 performed an alanine scanning of all Tcf4 residues on the binding interface and found that in addition to Asp16, which is the most important in binding, alanine replacement of Asp11 also resulted in a significant decrease in the binding constants. The Tcf4 8-30 peptide we used in the FP assay included these three important residues (Asp11, Asp16, and Glu17).
Competitive Binding Assay
We further performed an FP-based competitive binding assay to validate our established β-catenin-Tcf4 8-30 assay system for investigation of protein-ligand interaction and for potential HTS. As shown in Figure 2 , the binding between tracer (20 nM) and β-catenin (1.5 µM) was inhibited by either unlabeled Tcf4 8-30 peptide or GST-Tcf4-1-54 protein, with IC50 values of 5.7 µM and 1 µM, respectively. The binding activities were unaffected in the presence of 1% to 10% DMSO (p > 0.05).
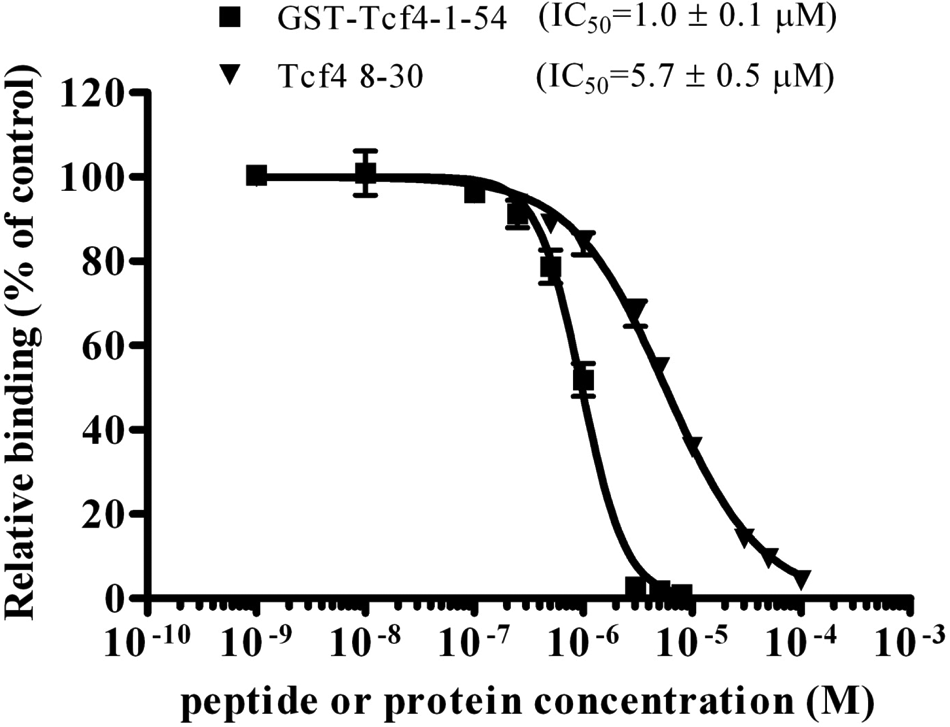
Competitive binding assay. A mixture of 20 nM tracer and 1.5 µM β-catenin was incubated in the presence of different concentrations of unlabeled 8-30 peptide (1 nM to 100 µM) or purified recombinant GST-Tcf4-1-54 protein (1 nM to 10 µM). Fluorescence polarization values were recorded after 3 h. The experiments were performed three times and data were analyzed in Microsoft Excel (Microsoft, Redmond, WA) and plotted in Prism 4 (GraphPad Software, San Diego, CA).
We also applied this competitive assay to map the minimum region of Tcf4 required for the interaction. As shown in Table 1 , the IC50 value for Tcf4 10-27 peptide is only two-fold higher than 8-30 (11 µM vs. 5.7 µM), suggesting that 10-27 might be the minimum part for the β-catenin/Tcf4 binding. Although our data showed that Tcf4 13-30 is not able to efficiently inhibit (<20% inhibition at 100 µM) the β-catenin/Tcf4 interaction (not consistent with one of the previous reports), 13 because it lacks Asp11 of Tcf4, this result does agree with another report that shows that Asp11 is also important for the interaction. 11
Determination of Minimal Region in Tcf4 for Binding to β-Catenin
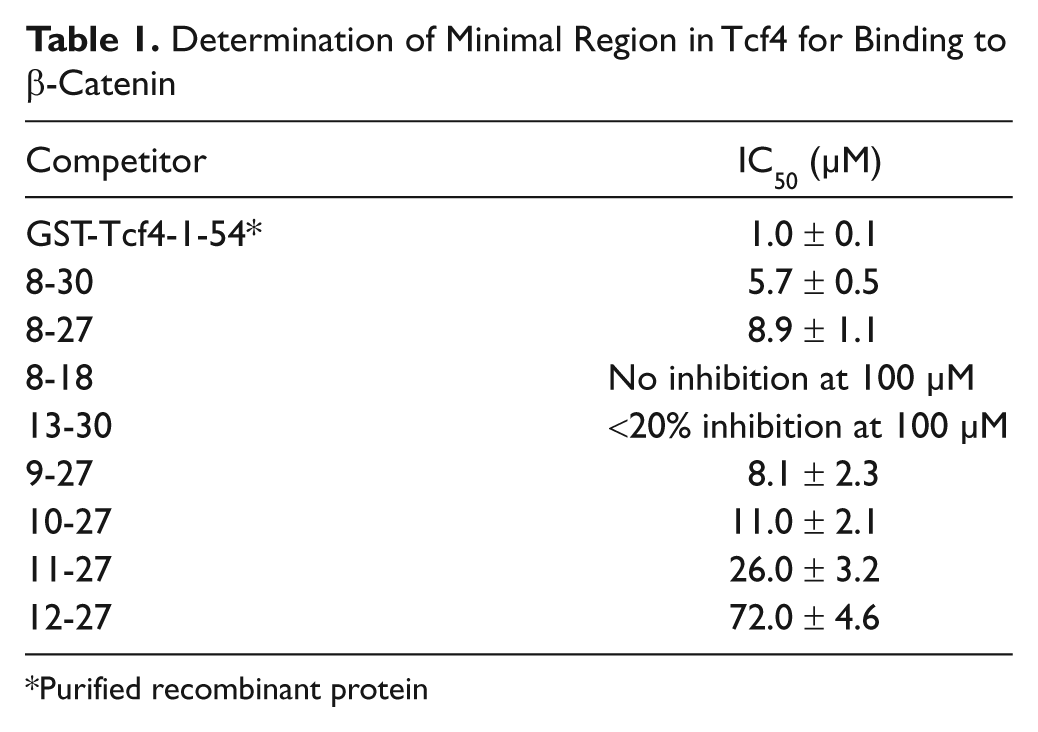
Purified recombinant protein
FP-Based Binding Assay Is Suitable for HTS
We further evaluated the quality and suitability of this assay for HTS by testing the well-to-well and plate-to-plate variations of free tracer (20 nM tracer) and bound tracer (polarization values recorded for 20 nM with 1.5 µM β-catenin). Thirty-six repetitions were tested for both free tracer and bound tracer in the same plate. The average values for free tracer and bound tracer were 118 ± 6.6 and 226.2 ± 2.3, respectively. The Z′ factor for well to well was 0.75 ( Fig. 3A ). The FP binding experiments were also conducted in twenty-six 96-well plates on different days. As shown in Figure 3B , the average values for free and bound tracers were 119.3 ± 4.8 and 232.6 ± 4.7, respectively. The Z′ value was also 0.75, which is excellent for HTS.
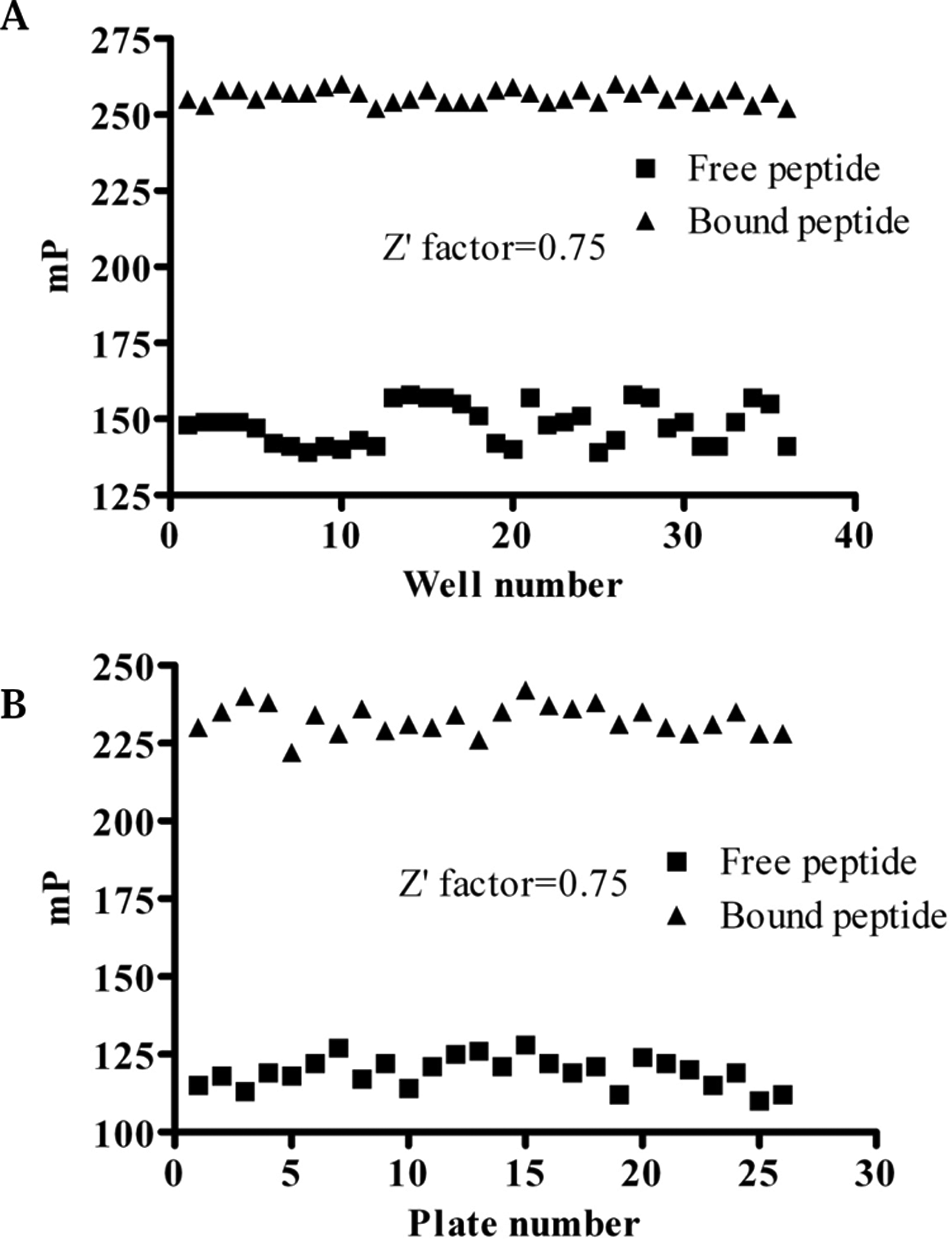
Evaluation of fluorescence polarization–based assay for high-throughput screening. (A) The well-to-well signal-to-noise ratios and Z′ factors. (B) The plate-to-plate signal-to-noise ratios and Z′ factors.
Although Tcf4 8-30 peptide retains its ability to interact with β-catenin, other regions in Tcf4 also contribute to the binding activity, such as hydrophobic residues (Leu41, Val44, and Leu48).11,13 It should be noted that some drug candidate hits might be missed when using this tracer in binding assays. Therefore, further development of the FP-based assay using the full-length Tcf4 CBD (N-terminal 1-54) as a probe may add more dimensions to FP-based, β-catenin/Tcf interaction-targeted drug discovery. In addition, β-catenin is a multiple functional protein. Besides the Tcf family, it also interacts with other proteins such as E-cadherin and APC. Crystal structures revealed that the Tcf binding groove overlaps with that for E-cadherin and APC.5,14,15 It is important to analyze and determine the selectivity of lead compounds when using this assay.
In summary, we have established a new FP-based assay that can be used to measure the β-catenin/Tcf4 peptide interaction. This assay is fast and robust and shows good reproducibility. At concentrations below 10%, DMSO had no effect on this assay. The assay can be used for HTS for identifying small-molecule inhibitors of the β-catenin/Tcf4 interaction.
Footnotes
Acknowledgements
The authors thank Dr. Wenqing Xu (University of Washington) for providing the β-catenin expression vector.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: The project described was supported in part by grants to JA from the Connolly Endowment/Hendricks Fund and the LUNGevity Foundation and the Carol M. Baldwin Breast Cancer Research Fund.