
Abstract
Inhibitors of human dimethylarginine dimethylaminohydrolase-1 (DDAH-1) are of therapeutic interest for controlling pathological nitric oxide production. Only a limited number of biologically useful inhibitors have been identified, so structurally diverse lead compounds are desired. In contrast with previous assays that do not possess adequate sensitivity for optimal screening, herein is reported a high-throughput assay that uses an alternative thiol-releasing substrate, S-methyl-L-thiocitrulline, and a thiol-reactive fluorophore, 7-diethylamino-3-(4′-maleimidylphenyl)-4-methylcoumarin, to enable continuous detection of product formation by DDAH-1. The assay is applied to query two commercial libraries totaling 4446 compounds, and two representative hits are described, including a known DDAH-1 inhibitor. This is the most sensitive DDAH-1 assay reported to date and enables screening of compound libraries using [S] = KM conditions while displaying Z′ factors from 0.6 to 0.8. Therefore, this strategy now makes possible high-throughput screening for human DDAH-1 inhibitors in pursuit of molecular probes and drugs to control excessive nitric oxide production.
Introduction
One potential advantage for indirect regulation of NO production through the inhibition of DDAH-1, rather than direct inhibition of NO synthase, has been proposed for the treatment of septic shock, which is characterized by an NO-mediated drop in blood pressure. 1 Inhibitors of NO synthase can block vascular NO production but would also inhibit NO production in macrophages, compromising their ability to fight off infection and worsening sepsis. However, because DDAH-1 is not significantly expressed in immune cells but does regulate vascular dimethylarginine levels and NO production, selective inhibitors of DDAH-1 could overcome this limitation. 1
To date, only a limited number of human DDAH-1 inhibitors have been identified, and many of these are substrate analogs.3–5 The dependence of these highly charged inhibitors on the y+ cationic transport system places limits on which modifications are tolerated. A few classes of substrate-dissimilar inhibitors have been reported, including indolylthiobarbituric acids, 6 pentafluorophenyl sulfonates, 7 4-halopyridines, 8 and ebselen, 9 but these have mostly been derived from studies using the Pseudomonas aeruginosa isoform of DDAH, which has only 25% sequence identity to human DDAH-1. Notably, indolylthiobarbituric acid inhibitors of P. aeruginosa DDAH are unable to inhibit human DDAH-1, 5 emphasizing the critical need to use the human DDAH-1 isoform for high-throughput screening (HTS).
However, no suitable HTS assay has been reported for human DDAH-1. A 96-well plate colorimetric assay has been developed to detect the product citrulline through derivatization but uses harsh conditions and heating steps that are not scalable. 10 We recently developed an alternative colorimetric HTS assay for P. aeruginosa DDAH, 9 but the lower kcat and KM values of human DDAH-1 preclude its effective use. Neither assay was found to have sufficient sensitivity to detect linear product production by human DDAH-1 under [S] = KM conditions, which allow for the balanced detection of competitive, noncompetitive, and uncompetitive inhibitors. Because all of these different inhibitor types are of interest, we sought to develop a more sensitive assay that could reliably detect nanomolar quantities of product and enable HTS of human DDAH-1.
Overcoming these limitations, a fluorescent HTS assay is developed in which human DDAH-1 catalyzes the hydrolysis of an alternative substrate, S-methyl-L-thiocitrulline (SMTC), to produce methanethiol, which subsequently reacts with the sensitive fluorogenic probe 7-diethylamino-3-(4′-maleimidylphenyl) -4-methylcoumarin (CPM) 11 to provide an increase in fluorescence (Exmax ≈ 385 nm, Emmax ≈ 465 nm) that can be followed in a continuous manner. The assay was optimized, miniaturized to 384-well plate format, and used to screen human DDAH-1 against two commercial libraries totaling 4446 compounds for validation. This is shown to be the most sensitive DDAH assay reported to date, capable of detecting approximately 70 nM of product, and it displays a mean Z′ factor of 0.71, which now enables the optimal HTS of human DDAH-1 for discovery of novel compounds to control NO production.
Materials And Methods
Materials
All reagents were purchased from Sigma-Aldrich (St. Louis, MO), unless otherwise specified. Black 384-well polypropylene plates (catalog no. 264576) were from Nalge Nunc (Rochester, NY). Microcentrifuge tubes of 1.5 mL were purchased from Fisher Scientific (Pittsburgh, PA).
CPM stock solution was prepared by dissolving CPM in dimethyl sulfoxide (Fisher Scientific, Pittsburgh, PA) to 2 mM and stored in 1 mL aliquots in opaque amber microcentrifuge tubes (Fisher Scientific) at −20 °C.
Expression and purification of human DDAH-1
Recombinant His-tagged human DDAH-1 was expressed in and purified from BL21 Escherichia coli cells as previously described. 4 Protein was then subjected to overnight dialysis at 4°C in 1 L of 2 mM 1,10-phenanthroline, 10 mM KH2PO4, and 100 mM KCl (pH 7.3), followed by three consecutive 4 h dialysis steps at 4°C into 1 L of 10 mM KH2PO4 and 100 mM KCl (pH 7.3) containing glycerol (10% v/v) and treated with Chelex-100 (Bio-Rad Laboratories, Hercules, CA). Protein concentration was determined by first denaturing a 30 µL aliquot of protein in 6 M guanidinium chloride, 20 mM KH2PO4 buffer (pH 6.5), and then measuring absorbance at 280 nm using the published extinction coefficient (7680 M−1cm−1). 4 The remaining protein was aliquoted and stored at −80°C. The recombinant protein has steady-state rate constants similar to DDAH-1 isoforms isolated from mammalian sources. 12
Typical assay
Typically, enzyme solution was prepared by adding recombinant human DDAH-1 (40–60 nM) to assay buffer (344 mM KH2PO4, 344 mM KCl, 0.02% Tween-20, 4 mM EDTA, pH 8.0). Substrate+CPM solution was prepared by adding SMTC (0.75 µM) and CPM (7.1 µM) to CPM buffer (5 mM KH2PO4, 5 mM KCl, 0.02% Tween-20, pH 2.5) and sealing in a polypropylene tube shielded from light at room temperature. Enzyme solution (45 µL) was dispensed into each well of a 384-well black polypropylene plate. Substrate+CPM solution (45 µL) was then added to enzyme solution to initiate the reaction, making the final concentrations 30 nM human DDAH-1, 0.4 µM SMTC, and 3.6 µM CPM, with a final reaction pH of 8.0. Plates were loaded into the Wallac 1420 plate reader with filters for excitation at 355 nm and emission at 460 nm to measure product formation at room temperature and to generate a fluorescence versus time plot, from which initial rates were determined. All reactions were run at least in triplicate.
Effect of varying enzyme concentration
DDAH-1 (0–420 nM) was added to reactions containing 7 µM SMTC under the typical assay conditions described above. Rates were determined as above and plotted against enzyme concentration, and all reactions were run in triplicate.
Steady-state kinetics
Enzyme solution was prepared as described above. Substrate+ CPM solutions were prepared with SMTC (0–23 µM) and CPM (7.1 µM) in CPM buffer. Substrate+CPM solution (45 µL) was dispensed into a black 384-well polypropylene plate, and reactions were initiated by adding enzyme solution (45 µL). Reactions were monitored as described above.
HTS assay
HTS enzyme solution (65 µL) consisting of 238 mM KH2PO4, 238 mM KCl, 0.02% Tween-20, and 4 mM EDTA (pH 8.0) with or without 41.5 nM DDAH-1 was dispensed into each well. For the Chembridge Fragment Library, library compounds (10 mM in DMSO) were diluted 10-fold prior to use. Library compounds (1–10 mM) or DMSO without library compound (0.9 µL) was then dispensed into each plate using a Janus Workstation (PerkinElmer, Waltham, MA) and mixed by pipetting up and down with the dispenser. Typically, columns 1 and 2 were used as positive controls (with enzyme and without library compound), and columns 23 and 24 were used as negative controls (without enzyme or library compound), for a total of 32 positive and 32 negative controls per plate. The remaining 320 wells each contained a library compound. Plates were centrifuged at 1000g for 1 min and incubated at room temperature for 10 min. Then, 25 µL of HTS substrate+CPM solution (12.6 µM CPM, 1.4 µM SMTC, 5 mM KH2PO4, 5 mM KCl, 0.02% Tween-20, pH 2.5) was added to each well. Final concentrations were 30 nM human DDAH-1, 0.4 µM SMTC, and 3.6 µM CPM, and the final reaction pH was 8.0. Plates were centrifuged for 1 min at 1000g to remove any bubbles and loaded into an Envision Microplate Reader (PerkinElmer) with filters for excitation at 355 nm and emission at 460 nm. Fluorescence of each well was read 10 times over approximately 10 min. Rates were determined by taking the slope of the fluorescence versus time data.
Orthogonal assay of hits
Samples with and without DDAH-1 (1 µM) were incubated in 70 mM KH2PO4, 100 mM KCl, 1 mM EDTA, 0.02% Tween-20, pH 7.3, with and without inhibitor (1 mM). Reactions were initiated with 0.5 mM Nω-methyl-L-arginine (final reaction volume = 60 µL) and incubated at room temperature for 30 min. Samples were quenched with 3 µL of 6N trifluoroacetic acid, and citrulline produced during the reaction was derivatized and detected as described. 10
Analysis and curve fitting
Qtiplot 0.9.8 (ProIndep Serv SRL, Craiova, Romania) was used for all curve fitting. Normalized percentage inhibition was calculated using the in-plate controls by the equation. Initial rates for compoundsshowing ≥50% inhibition that also showed background fluorescence less than threefold from the mean of the negative controls was further corrected for possible interference by, 13 the factor where Flcontrols and Flcompound are the background fluorescence values estimated by linear extrapolation to the time the reaction was initiated. Z′ factors were determined as previously described. 14 Signal-to-background ratio was calculated by . Intraplate coeffcient ofvariation (%CV) was calculated as , whereσ and µ are the standard deviation and mean, respectively, of the uninhibited controls.
Results And Discussion
In our hands, the two preexisting colorimetric DDAH assays9,10 did not have adequate sensitivity or discriminating power for HTS of human DDAH-1 (Z′ factors <0, data not shown). Therefore, we developed a fluorescent assay for human DDAH-1 using CPM-based detection of methanethiol liberated from an alternative substrate, SMTC (
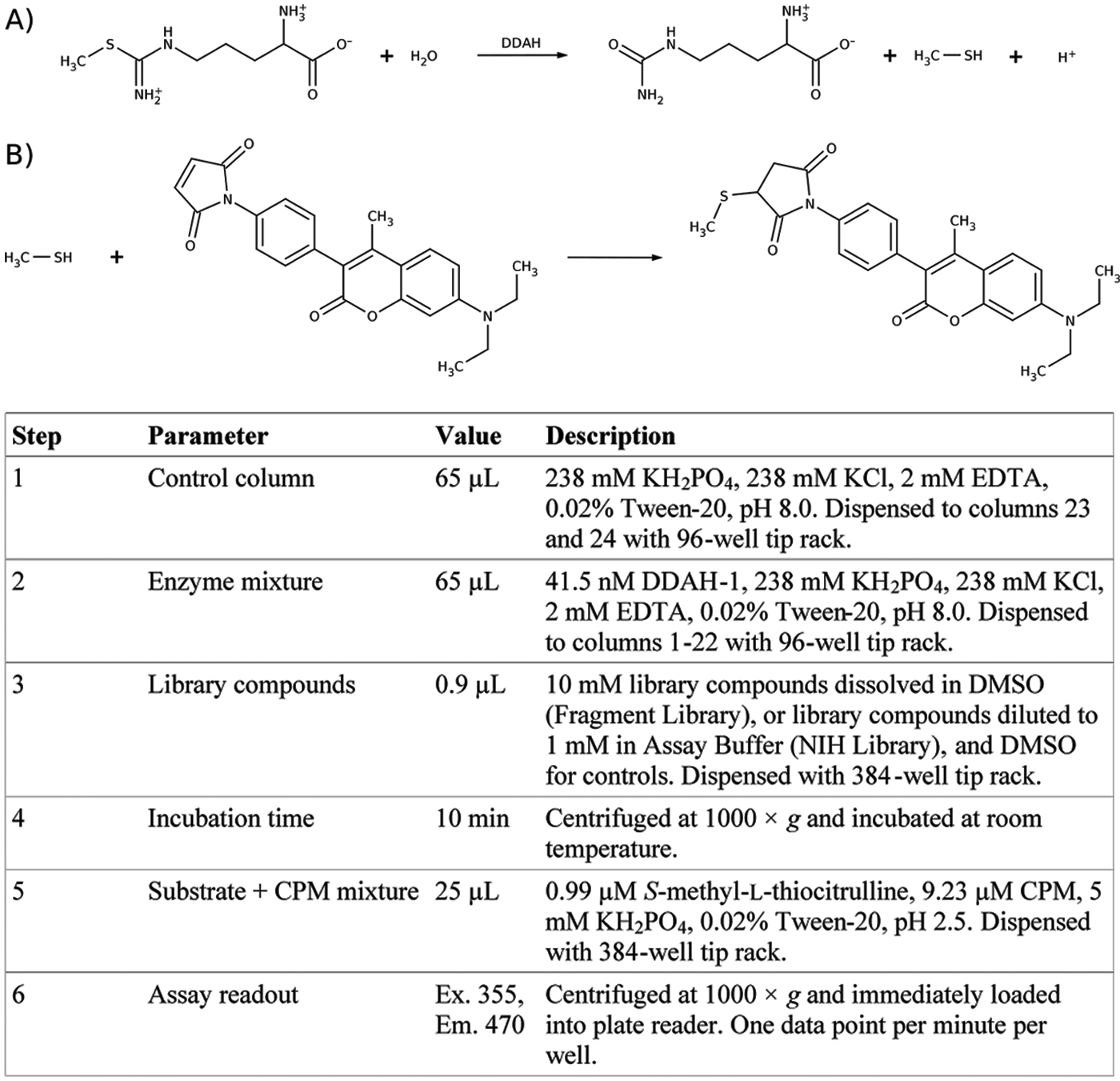
High-throughput screening assay protocol. (
Development of CPM-based DDAH assay
Because CPM is known to have low aqueous solubility and is susceptible to alkaline hydrolysis,
15
we optimized the stock solution and assay conditions to minimize precipitation and cosolvent addition while ensuring that the rate of enzymatic hydrolysis is rate limiting. To maximize stability and solubility, CPM (7–13 µM) was kept at pH 2.5 prior to the kinetic assay. Upon final dilution, the CPM (3.5 µM) remains soluble at the assay pH of 8.0. A slow background hydrolysis of CPM is observed under assay conditions, but the rate of hydrolysis was not significant compared with the rate of the enzyme-catalyzed reaction (
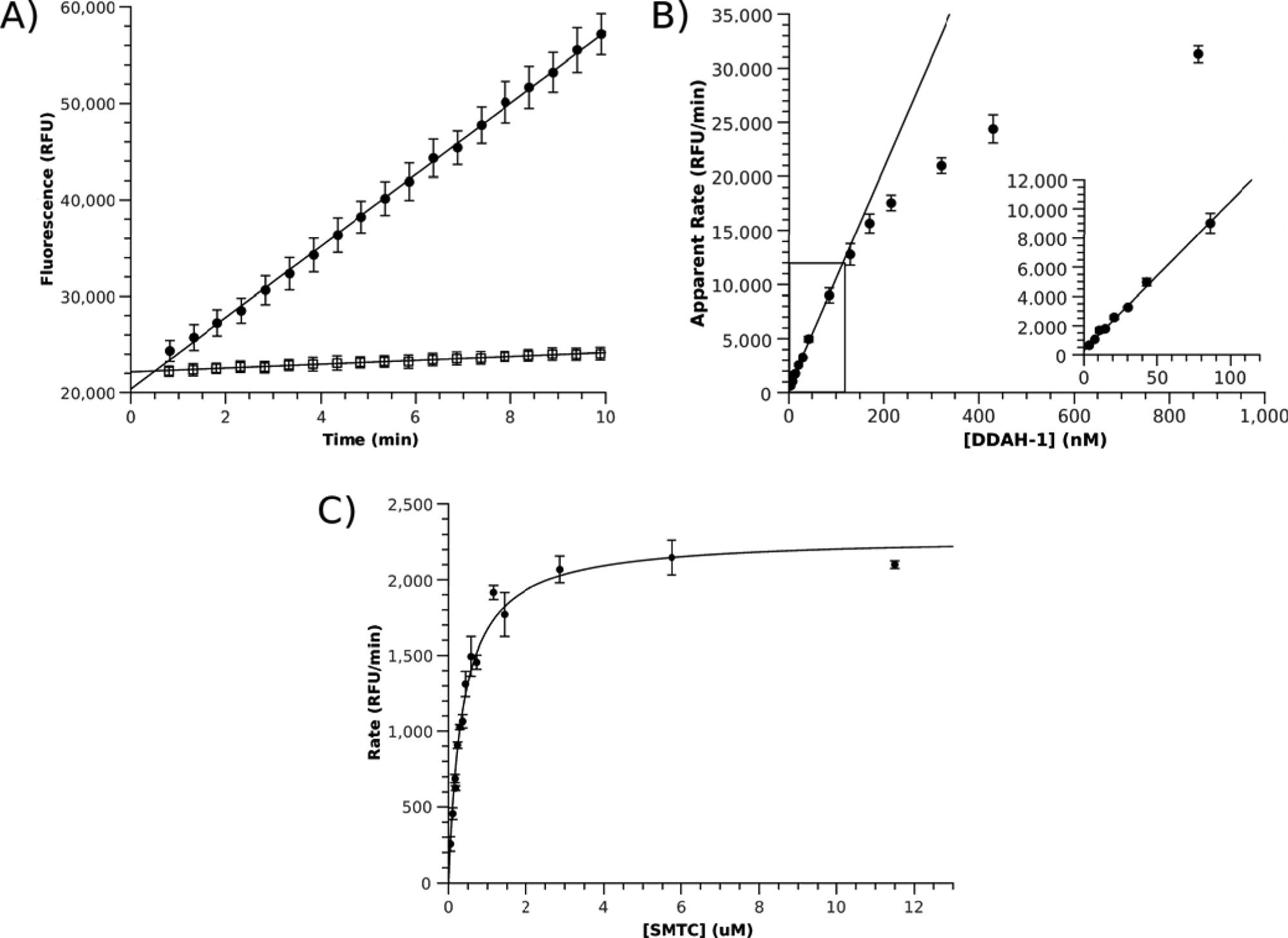
Development and optimization of CPM-based assay for DDAH-1. (
The assay was then optimized to ensure that the initial increase in fluorescence was linear with time. Without detergent or blocking protein present, DDAH-1 exhibited significant time-dependent loss of activity upon addition of enzyme to wells. Albumin showed a background reaction with CPM, but addition of beta-casein (0.02 mg/mL) or Tween-20 (0.02% w/v) successfully prevented loss of activity due to enzyme adsorption without background reaction. Ultimately, Tween-20 was chosen to include in the HTS because it has also been shown to minimize false-positive hits by nonspecific aggregators 16 and because it is known not to adversely affect DDAH-1 activity. 10
Although DDAH-1 contains a catalytic active-site Cys residue, fluorescence due to product formation increased linearly with time under kcat conditions for >20 min, indicating that during this time frame, CPM does not significantly inactivate DDAH-1 and that a significant fraction of CPM is not consumed. To ensure that initial rates were measured under the [S] = KM conditions used in HTS, the increase in fluorescence was measured and found to increase linearly with time for >10 min, indicating that a significant portion of substrate is not consumed.
CPM-based detection was then demonstrated to be more sensitive than existing DDAH assays. First, apparent reaction rates were measured at varying DDAH-1 concentrations under saturating substrate conditions. Rates varied linearly with DDAH-1 concentrations from about 120 nM to 3 nM, suggesting that the enzymatic reaction, rather than the reaction of CPM with methanethiol, is rate limiting in that concentration range. To further ensure that the reaction of CPM with methanethiol is not rate limiting, we tested the assay with a twofold higher CPM concentration and found that the rates observed with saturating substrate (4.6 µM SMTC; 60 µM DDAH-1) were within error of each other (6600 ± 200 RFU/min with 3.6 µM CPM versus 6200 ± 300 RFU/min with 7.2 µM CPM). The low enzyme concentrations used represent a significant improvement over previous assays that typically use 1 µM of DDAH-1 (
Using the optimized HTS conditions, steady-state Vmax (2280 ± 60 RFU/min) and KM (0.37 ± 0.03 µM) values for human DDAH-1 (20 nM) were determined using a Victor Wallac Plate Reader (
CPM HTS assay development
The benchtop-optimized assay was then optimized for HTS by testing its compatibility with DMSO cosolvent and the robotic liquid-handling steps and by stabilizing the assay components so that reagents can be prepared and stored for several hours before use. Consistent with a previous report,
5
human DDAH-1 is shown to be quite tolerant to DMSO, showing retention of nearly 100% activity in 15% DMSO and 50% activity in 50% DMSO (
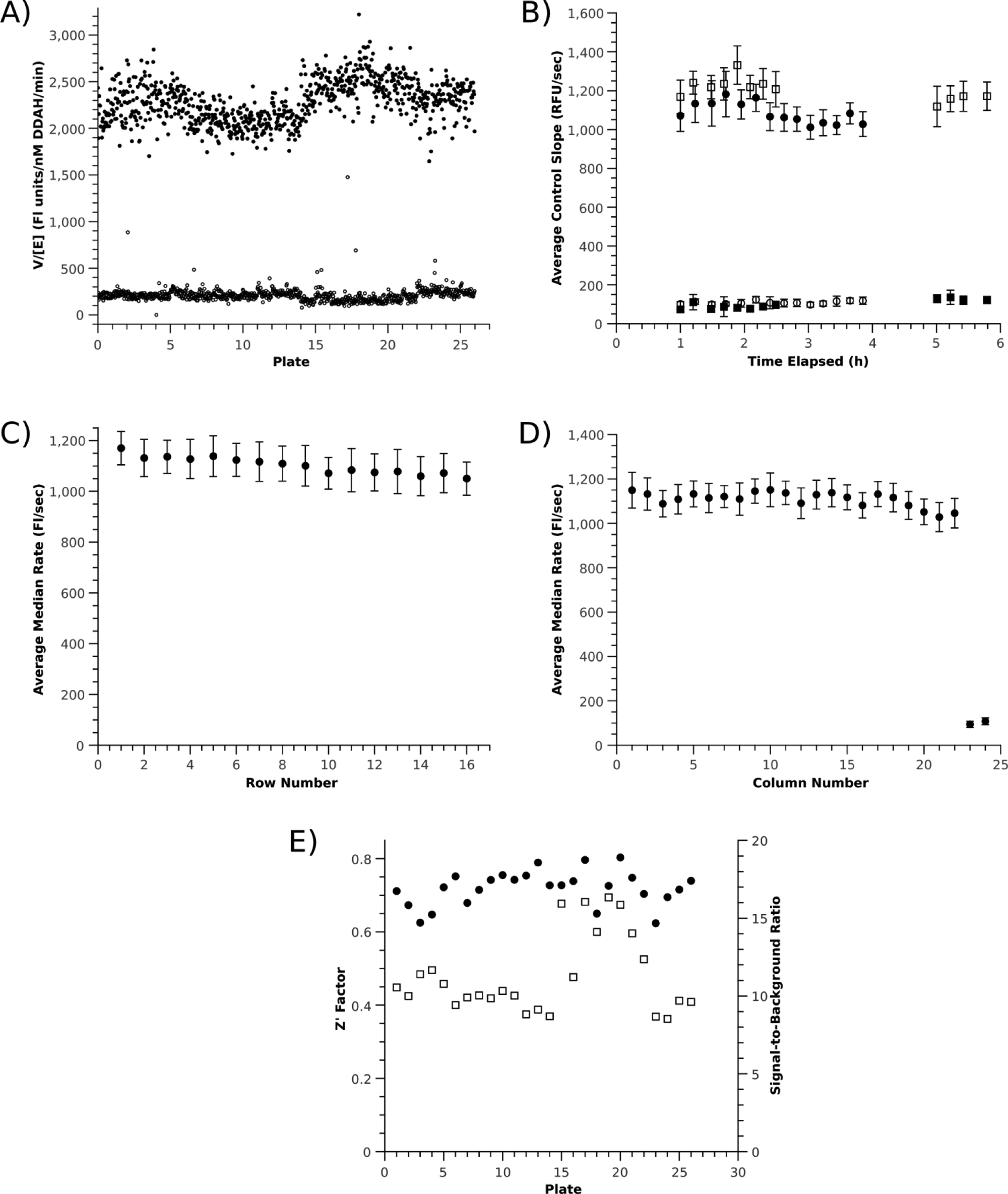
Controls from high-throughput screening of the Chembridge Fragment Library. (
The stability of assay reagents was also shown to be adequate. The CPM assay retained approximately 80% of its fluorescence for at least 4 h. The small loss in fluorescence did not appear to affect Z′ factors adversely, which remained high (0.86 as measured using manual pipetting to 24 wells) after 4 h. This loss of activity could be eliminated by mixing fresh CPM+substrate solution, and CPM+substrate solution kept sealed and protected from light overnight at room temperature showed almost 100% activity, suggesting that exposure to air or light during screening may slowly degrade the fluorophore.
HTS performance
The assay proved to be robust for HTS (
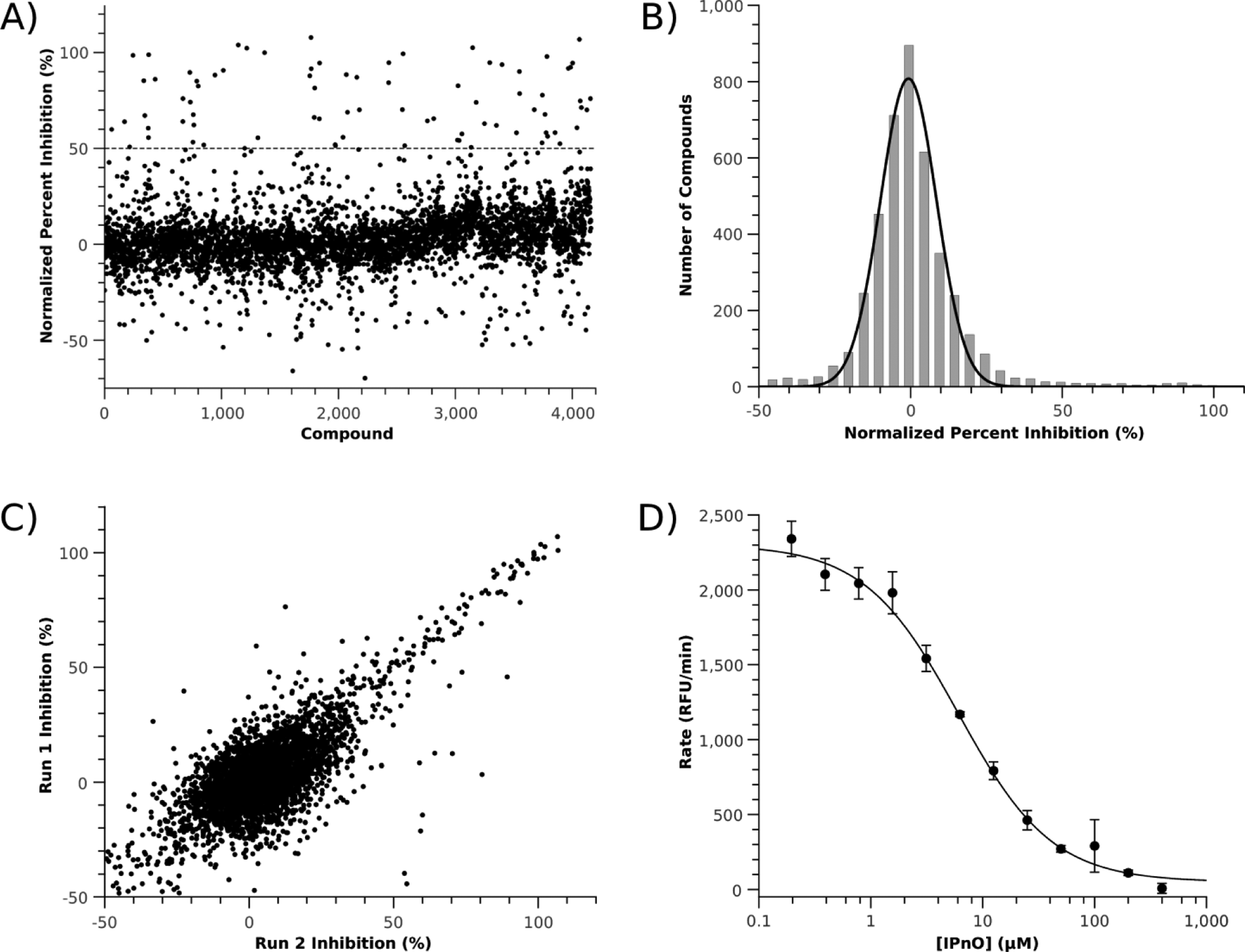
High-throughput screening results. (
Fluorescent HTS assays can be prone to interference by library compounds though quenching or high background fluorescence, resulting in false hits that show reduced rates of fluorescence increase even when the enzyme is fully active. Therefore, to minimize false-positives, library compounds with very high (i.e., beyond the detector’s limits) or low (possible quenching) background fluorescence were eliminated from consideration as follows: background fluorescence for each well was first estimated by extrapolating the linear fit of fluorescence versus time to a time point at which the positive and negative control lines intercept, which represents the time of mixing substrate with enzyme. The estimated fluorescence of each well at this time point is taken to be the background fluorescence. Compounds that exhibited >10-fold or <5-fold fluorescence relative to the background controls were discarded from consideration. Furthermore, compounds with observed rates 1.5-fold greater than the positive control mean were eliminated, as it is possible that they slowly reacted with CPM or otherwise enhanced the fluorescence of the CPM-thiol product. Of the 4446 compounds screened, 143 were eliminated using these metrics (3.2%). To further reduce false-positives from less efficient quenchers, we applied the fluorescence quenching corrections that have been previously described to compounds with greater than 50% inhibition, thus eliminating 40 false-positive compounds (0.9%). 13 The relatively high number of false-positives may be due to the high concentration of inhibitors used in the screen.
Post-HTS processing
Because a thiol-reactive reagent was used for screening, false hits could be a result of compounds that scavenge the methanethiol product or compounds that directly react with CPM. Therefore, to avoid unnecessary characterization of these compounds, all primary hits showing >50% inhibition of human DDAH-1 were retested for inhibition by using a preexisting orthogonal, lower throughput, secondary assay, which uses the naturally occurring substrate Nω-methyl-L-arginine (kcat = 0.79 min−1; KM = 90 µM) 12 and a discontinuous colorimetric derivatization of the urea group in the resulting product L-citrulline. 10
Starting with the NCC library, two compounds showing >50% inhibition were identified, ebselen (109% inhibition) and rabeprazole (53% inhibition). Rabeprazole did not show any inhibition of DDAH-1 when assayed using the secondary assay, indicating it is likely a false-positive. It is presumed that the false signal derives from the known conversion of this prodrug to a thiol-reactive species, 17 which could directly react with the methanethiol product. In contrast, the other hit, ebselen, has already been mechanistically characterized and demonstrated to be a bioavailable inhibitor of DDAH-1, with an in-cell IC50 value of 36 ± 7 µM. 9 Therefore, as a proof of principle, this HTS assay is shown to successfully identify a known DDAH-1 inhibitor. Starting with the fragment library, 81 hits were identified as primary screening hits. Of these compounds, 49 compounds were selected for further validation, and 42 have been verified as human DDAH-1 inhibitors to date by using the orthogonal secondary assay and will be described in full elsewhere. The higher percentage of primary hits derived from the fragment library (2.0%) compared with the NCC library (0.4 %) is consistent with expectation.
As an additional validation that the assay is useful for generating dose-response data, the concentration dependence of N
5
-(1-iminopropyl)-L-ornithine, a known DDAH-1 inhibitor, was studied (
In summary, a continuous fluorescent assay for human DDAH-1 has been developed, validated, and used to screen two commercial libraries. In the assay’s current form, one 384-well plate containing 320 compounds can be screened every 10 min with a Z′ value of 0.71, and the throughput of the assay can likely be increased by substituting continuous reads with single endpoint reads. To our knowledge, this is the most sensitive DDAH assay reported to date and is the only assay amenable for HTS of human DDAH-1 under [S] = KM conditions. The sensitivity of the assay is such that it likely could be tailored for detection of weak-binding compounds using [S] < KM conditions. This assay should greatly facilitate the search for novel molecular probes and therapeutic agents for the control of NO production.
Footnotes
Acknowledgements
We thank Dr. Eun Jeong Cho and the Automation and High Throughput Screening Facility at the Texas Institute for Drug and Diagnostic Development (TI-3D, University of Texas, Austin) for assistance with the HTS. This work was supported in part by grants from the National Institutes of Health (GM69754 to W.F.) and the Robert A. Welch Foundation (F-1572 to W.F.) and from a seed grant from the CTT/TI-3D Chemistry & Molecularly-Targeted Therapeutic Development grant program.
Three figures showing Gaussian distribution and separation of positive and negative controls for an example screening plate, per-plate coefficient of variation, and DMSO tolerance of DDAH-1.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.