
Abstract
Specific viral proteins enter the nucleus of infected cells to perform essential functions, as part of the viral life cycle. The integrase (IN) molecule of human immunodeficiency virus (HIV)–1 is of particular interest in this context due to its integral role in integrating the HIV genome into that of the infected host cell. Most IN-based antiviral compounds target the IN/DNA interaction, but since IN must first enter the nucleus before it can perform these critical functions, nuclear transport of IN is also an attractive target for therapeutic intervention. Here the authors describe a novel high-throughput screening assay for identifying inhibitors of nuclear import, particularly IN, based on amplified luminescent proximity homogeneous assay (AlphaScreen®) technology, which is high throughput, requires low amounts of material, and is efficient and cost-effective. The authors use the assay to screen for specific inhibitors of the interaction between IN and its nuclear transport receptor importin α/β, successfully identifying several inhibitors of the IN/importin α/β interaction. Importantly, they demonstrate that one of the identified compounds, mifepristone, is effective in preventing active nuclear transport of IN in transfected cells and hence may represent a useful anti-HIV therapeutic. The screen also identified broad-spectrum importin α/β inhibitors such as ivermectin, which may represent useful tools for nuclear transport research in the future. The authors validate the activity and specificity of mifepristone and ivermectin in inhibiting nuclear protein import in living cells, underlining the utility of the screening approach.
Introduction
A
Molecules >45 kDa generally require specific signals known as nuclear localization signals (NLSs) to gain nuclear entry. Classically, nuclear protein import requires the recognition of an NLS in the cargo protein by members of the importin (IMP) superfamily of nuclear import receptors, of which there are multiple α and β types. NLSs are most commonly recognized by the IMP α subunit of the IMP α/β heterodimer, whereas the IMP β subunit mediates docking to and translocation through the NPC (for reviews, see Pemberton and Paschal 5 and Poon and Jans 6 ). More recently, however, IMP β itself and many of its homologues have been shown to mediate nuclear import of NLS-containing cargoes independent of IMP α. 7-10 In all cases, the binding of the guanine nucleotide binding protein Ran in its guanosine triphosphate (GTP)–bound form to IMP β dissociates the complex, releasing the cargo into the nucleus. Nuclear protein export occurs in an analogous fashion, requiring nuclear export signals (NESs) to be recognized by exportin proteins (IMP β homologues). Currently, the only commercially available compound for inhibiting nuclear transport is leptomycin B (LMB), which is a highly specific inhibitor of the exportin CRM-1. Other inhibitory compounds have been suggested, including structurally related compounds such as ratjadone, peptide-based inhibitors, and several small-molecule inhibitors, 11-15 but these are almost exclusively limited to inhibition of nuclear export mediated by CRM-1. Due to the critical role played by viral protein nuclear import, it is clear that the NLS-IMP interaction itself is a very attractive target for the development of small-molecule inhibitors.
Human immunodeficiency virus (HIV) is one of the most widespread human diseases, with an estimated 6000 new infections daily. 16 The high mutation rate of the HIV virus leads to a marked prevalence of resistance among patients receiving antiretroviral drugs, resulting in a dire need for new targets and classes of effective antiviral therapeutics. A critical part of the HIV replicative cycle is the transport of newly synthesized viral cDNA coated with viral and cellular proteins in what is known as a preintegration complex (PIC) into the nucleus. 17,18 Once inside, the PIC is disassembled, and the HIV integrase (IN) protein, a critical component of the PIC, integrates the viral cDNA into the host cell genome, from which viral mRNA is then synthesized. Due to this critical role of IN, there is already 1 IN-targeted antiviral therapeutic licensed for clinical use and several others under development, but these compounds usually target the IN-DNA interaction, and there is significant cross-resistance between these various IN inhibitor drugs, highlighting the urgent need for the development of IN-targeting drugs with alternative mechanisms of action. Since IN must first enter the nucleus to perform these critical functions, and this is dependent on strong (µM) interactions with IMP α/β, 19 nuclear import of IN is a highly attractive target for therapeutic intervention.
In this study, we describe a high-throughput screening (HTS) assay to identify inhibitors of NLS-IMP interactions and document its utility by performing a pilot screen for inhibitors of IN-IMP α/β binding. The assay is based on our previous work examining the strength of IMP-NLS interactions using AlphaScreen® (amplified luminescent proximity homogeneous assay) technology (PerkinElmer, Waltham, MA). 20 The AlphaScreen® technique eliminates many of the technical difficulties associated with other protein-protein binding assays. 20,21 The assay is performed in solution with proteins in native conformation, and the energy transfer between binding partners is via an excited oxygen singlet diffusing through solution over distances up to 200 nm. This means that protein conformation and complexation geometry are not limiting factors. The assay is performed in a final volume of only 25 µL with nanomolar amounts of each reagent used and is compatible with most solvent and protein buffer systems. 20,21 Here we modify the assay for high-throughput library screening to identify small-molecule inhibitors of critical biological interactions. Through our screening/counterscreening strategy, we identify several specific inhibitors of IMP α/β-IN binding, as well as more general inhibitors of IMP α/β-mediated nuclear transport. One of each is validated for activity in living cells, underlining the utility of the screening assay.
Materials and Methods
Generation of GFP-fusion protein bacterial and mammalian expression plasmids
Bacterial or mammalian cell expression vectors encoding green fluorescent protein (GFP)–IN and GFP-Op-T-NLS, containing the full-length HIV-1 IN (from the pNL4-3 proviral HIV-1 clone) or the optimized protein kinase CK2 site-enhanced T-ag NLS (Op-T-NLS: SSDDEATADAQHAAPPKKKRKV; single-letter code), respectively, were generated using the Gateway cloning technology (Invitrogen, Carlsbad, CA) and vector pGFP-attC 20,22 for GFP-fusion protein expression in bacteria or pDest53 (Invitrogen) for GFP-fusion protein expression in mammalian cells. 23
Protein purification and IMP α/β dimerization
His6-tagged IN protein was expressed from the pINSD. His.Sol plasmid in BL21 (DE3) bacteria as described.
19,24
GFP-tagged Op-T-NLS fusion protein was purified from bacteria as a His6-tagged protein under denaturing conditions as described previously.
20
The IMP proteins were purified from bacteria as GST-fusion proteins under native conditions as previously described.
20,25,26
Where appropriate, IMP α and β were predimerized as previously described
19,20,22,23,26-29
at 13.6 µM for 15 min at room temperature in intracellular buffer (IB; 110 mM KCl, 5 mM NaHCO3, 5 mM MgCl2, 1 mM EGTA, 0.1 mM CaCl2, 20 mM HEPES, 1 mM dithiothreitol [DTT], pH 7.4) to generate the IMP α/β heterodimer for binding studies. The efficiency of dimerization was monitored using a gel mobility shift assay as per
Biotinylation of IMP proteins
IMP α was biotinylated (α-B) as previously described using the Sulfo-NHS-Biotin reagent (Pierce, Rockford, IL). 20 Briefly, 3.5 mg of IMP was incubated with 250 µL of Sulfo-NHS-Biotin (1 mg dissolved in 150 µL H2O) on ice for 2 h. Free biotin was removed via a PD-10 column (Amersham, Buckinghamshire, UK), and the resulting biotinylated protein was concentrated to between 100 and 250 µM in an Amicon-30 concentration device in phosphate-buffered saline (PBS). No protein stability or precipitation issues have been observed in this concentration range.
AlphaScreen® binding assay
The AlphaScreen® assay was performed in triplicate in 384-well white opaque plates (PerkinElmer).
20
Briefly, 2 µL of 375 nM (30 nM final concentration) His6-IN was added to each well, followed by 20 µL of the appropriate IMP concentration (generally 0-60 nM), prepared by serial dilution in PBS, and incubation for 30 min at room temperature. All subsequent additions and incubations were made in subdued lighting due to photosensitivity of the beads. Then, 1 µL of a 1:10 dilution (in PBS) of the acceptor beads and 1 µL of 2.5% bovine serum albumin (BSA) were added simultaneously and incubated for 90 min at room temperature. Next, 1 µL of a 1:10 dilution of the donor beads was added to give a final sample volume of 25 µL and the mixture incubated at room temperature for 2 h. The assay was quantitated on a PerkinElmer FusionAlpha plate reader, triplicate values averaged, and titration curves (3-parameter sigmoidal fit;
Library screening for inhibitors of the IN/IMP α/β interaction
In total, 480 compounds selected at random from the LOPAC1280 (Library of Pharmacologically Active Compounds; Sigma, St. Louis, MO) were used for this proof-of-principle screen. The compounds (dissolved in 100% DMSO) were diluted to 50 µM with PBS (10% DMSO final concentration), with a no-compound control sample (PBS/10% DMSO only) included in 6 replicate wells per plate. The high-throughput assay was carried out in triplicate in 384-well microplates, essentially as above, except that following addition of 2 µL His6-IN to the wells, 5 µL of each diluted compound or 5 µL PBS/10% DMSO alone (no compound control) was added, followed by 15 µL of IMP α/β at the predetermined optimal concentration. The remaining additions and incubations times were all carried out as above. Appropriate control samples were also included in triplicate on each plate, including 25 mM imidazole (positive control for assay inhibition) and a control for AlphaScreen® signal generation (Op-T-NLS binding to IMP α/β). Results are determined as percent inhibition relative to the no-compound control from the same assay plate, taking into account the background signal. Assay robustness was confirmed through calculation of the Z′ factors as per Zhang et al. 31
Library counterscreening and cross-screening
A counterscreen was carried out as described above to exclude false-positive compounds by replacing His6-IN with 5 nM His6-Biotin. Results of the library screen were normalized to the results of the counterscreen to generate fold inhibition data. Similarly, a library cross-screen was performed on positive compounds identified from the IN screen to identify inhibitors specific for the IN-NLS interaction rather than general NLS-IMP interaction inhibitors, whereby His6-IN was replaced with 30 nM His6-GFP-Op-T-NLS.
Cell culture and transfection
HeLa (human cervical adenocarcinoma) cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS), 1 mM L-glutamate, 1 mM penicillin/streptomycin, and 20 mM HEPES at 37 °C in 5% CO2. Twenty-four hours prior to transfection, cells were seeded onto glass coverslips (15 × 15 mm). LipofectAMINE 2000™ (Invitrogen) was used according to the manufacturer’s instructions to transfect DNA into the HeLa cells. Cells were imaged live 24 h posttransfection by confocal laser scanning microscopy (CLSM; Bio-Rad 1024ES [Bio-Rad, Hercules, CA] or Olympus FV1000 [Olympus, Tokyo, Japan]) using a 60× oil or a 100× oil lens as described previously. 23,32 Digitized images were analyzed using the ImageJ v1.43g public domain software (National Institutes of Health, Bethesda, MD) to determine the ratio of nuclear (Fn) to cytoplasmic (Fc) fluorescence (Fn/c) according to the formula Fn/c = (Fn − Fb)/(Fc − Fb), where Fb is background autofluorescence. 8,33,34 P values were determined via Welch’s test using GraphPad Prism 5.0 (GraphPad Software, La Jolla, CA).
Results
Assay development and validation
This study aimed to establish a high-throughput assay to screen for inhibitors of the initial step of protein nuclear transport, the recognition of NLS sequences by IMPs. To this end, we modified our established AlphaScreen®-based assay for analyzing IMP binding to NLS-containing proteins 20 and chose to investigate HIV-1 IN and IMP α/β as proof-of-principle interacting proteins. The optimal concentration of His6-IN and the IMP α-B/β heterodimer for the assay was initially determined by titrating increasing concentrations of IMP α-B/β against 60 nM His6-IN ( Fig. 1A ) and fitting a sigmoidal curve to the experimental data. A ratio of 1:2 IN:Imp α/β was chosen for the subsequent assays as it was well below the binding capacity of the beads but still provided a robust signal that was not within the hooking zone (see Materials and Methods).
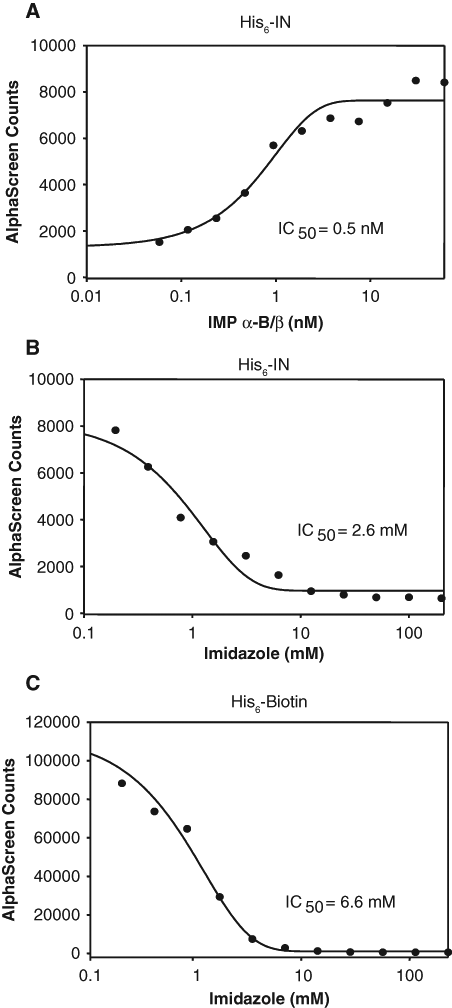
Validation of the integrase (IN)–importin (IMP) α/β inhibition AlphaScreen® assay. (
To test whether the assay was suitable for HTS for inhibitors of the interaction through detection of reduced signal, we tested optimized His6-IN:IMP α-B/β binding in the presence of increasing concentrations of imidazole ( Fig. 1B ). Imidazole prevents coupling of the His6-tag to the nickel-chelate acceptor beads, thus blocking signal generation. Imidazole inhibited the signal production with an IC50 of 2.6 mM, indicating that the assay can robustly detect decreases in signal and is therefore suitable for a screening application.
Nonspecificity counterscreen validation
To be able to discard false-positive compounds that interfere with the ability of the assay to generate a signal rather than the binding interaction itself, a counterscreen assay was developed. For this purpose, we used a biotinylated-His6 control (His6-Biotin) that binds the donor and acceptor beads together, generating an extremely strong signal. Any compounds that prevent coupling of proteins to either bead or interfere with the production/half-life of reactive singlet oxygen species will result in a decreased signal in the counterscreen assay. To verify that the counterscreen assay was suitable for detection of false-positive compounds, the assay was performed (see Materials and Methods) in the presence of increasing concentrations of imidazole ( Fig. 1C ), with a significant loss of signal observed (IC50 of 6.6 mM). These results indicate that the counterscreen assay can effectively identify compounds that interfere with AlphaScreen® signal production, which can then be discarded from further testing.
Screening of the LOPAC1280 library for inhibitors of the IN:IMP α/β interaction
The optimized His6-IN:IMP α-B/β binding assay was used to screen a partial library of pharmacologically active compounds (LOPAC1280; Sigma) for inhibitors of the interaction. Compounds were screened in triplicate as described in Materials and Methods, with appropriate comparison controls included on every plate. The set of inhibitors was in turn screened 2 times for a total of 6 replicates across 2 separate days. Hits were scored as those compounds that showed >70% inhibition of the signal ( Fig. 2A ) in both assays, with compounds generating a positive signal in only 1 assay discarded as outliers. The assay gave a consistent and robust response throughout the screening process with a median Z′ factor of 0.80 ( Fig. 2B ), reflecting the excellent signal/noise ratio achieved and the high reproducibility of the assay. Fourteen positive hits were identified from this initial screening process.
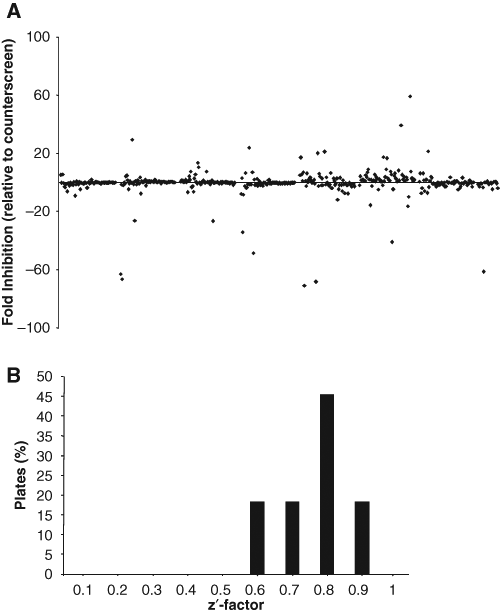
Screening of the LOPAC1280 library for inhibitors of the His6-IN:IMP α-B/β interaction. (
The entire compound library was then counterscreened using the His6-Biotin counterassay, with hits identified as those compounds able to reduce the counterscreen assay signal by more than 50% ( Fig. 2A ). Ten compounds were identified as significant false positives, including 2 imidazole ring–containing compounds, as expected. Six of the 14 positive hits above proved to be false-positive compounds and were excluded from further screening. Other compounds were identified that were able to significantly increase the counterscreen signal (>200% signal; false negatives), including compounds that generate excited singlet oxygen species when excited, such as protoporphyrin IX. False-negative compounds were not automatically removed from the screen as a proportion of these compounds were still identified as hits in the His6-IN:IMP α-B/β screen, indicating that they may be even more potent inhibitors of the interaction than initially suspected.
After taking the results of the counterscreen into consideration, 8 of the 480 initial compounds were identified as hits from the pilot screen, with compounds showing >70% inhibition of the signal representing a hit rate of 1.7%.
Cross-screening of the positive hits to determine specificity of inhibition
To determine the specificity of the compounds, a cross-screen was carried out to identify those compounds that generally inhibit IMP α/β binding to NLS sequences from those specific for the IN:IMP α/β interaction. To validate the stringency of our assay, the top 89 compounds (including the 8 true hits) identified from the initial pilot screen as inhibiting the IN:IMP α/β interaction >50% were cross-screened for the ability to inhibit GFP-Op-T-NLS binding to IMP α/β ( Fig. 3 ). Compounds showing >70% inhibition of the Op-T-NLS/IMP α/β interaction were deemed “nonspecific” inhibitors that either prevent IMP α/β recognition of NLS-containing proteins in general or may be interfering with formation of the IMP α/β complex itself; 5 of these were selected for further investigation. Of the compounds with no significant effect on GFP-Op-T-NLS:IMP α-B/β binding, 3 were considered specific inhibitors of the IN/IMP α/β interaction and selected for further investigation.
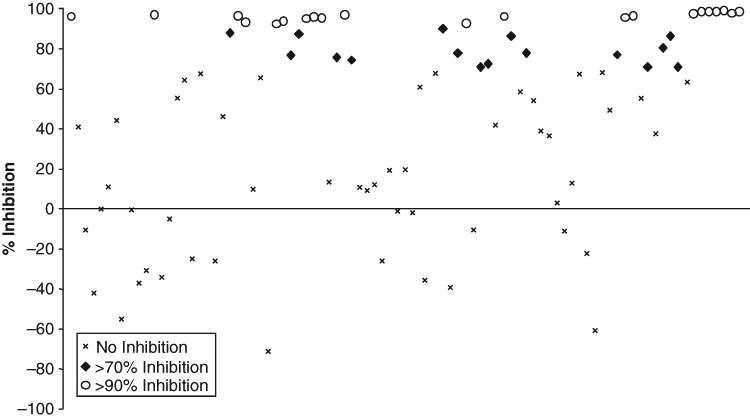
Determination of the specificity of the His6-IN:IMP α-B/β inhibitors identified from the library screen. Compounds identified as positive from the His6-IN:IMP α-B/β library screen were cross-screened as described in Materials and Methods for their ability to inhibit/enhance the binding of His6-GFP-Op-T-NLS to IMP α-B/β to determine the specificity of the inhibition of the compounds.
IC50 values were determined for 1 compound identified as a specific inhibitor of the IN/IMP α/β interaction (mifepristone) and 1 that reduced NLS binding to IMP α/β in general (ivermectin; Fig. 4 ). Concentration-response curves were determined using 30 nM (not shown) or 7 nM ( Fig. 4 ) IN, with almost identical results, both mifepristone and ivermectin being able to inhibit the IN:IMP α/β interaction at low µM concentrations (IC50s of 27.1 µM and 4.8 µM, respectively).
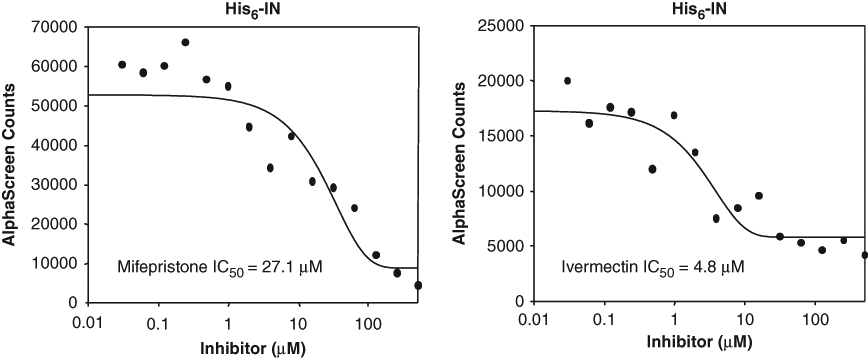
Determination of the strength of inhibition of mifepristone and ivermectin on integrase (IN). IC50 values determined using 7 nM IN and increasing concentrations of mifepristone (in phosphate-buffered saline [PBS]/2% EtOH) or ivermectin (in PBS/10% DMSO) as indicated; similar results were obtained at 30 nM IN (not shown).
Mifepristone and ivermectin inhibit IN nuclear import in cultured cells
To verify that compounds identified from the screen are indeed able to inhibit IN nuclear import in vivo, HeLa cells were transfected with GFP-IN or GFP-Op-T-NLS and treated with mifepristone or ivermectin at concentrations below their reported cellular toxicity levels. 35,36 Both compounds significantly inhibited the nuclear accumulation of GFP-IN compared to untreated controls (p = 0.0011 and 0.0030, respectively; Fig. 5A , B ). However, only ivermectin was able to reduce the nuclear accumulation of GFP-Op-T-NLS significantly (p < .0001), consistent with the idea that mifepristone is a specific inhibitor of the IN/IMP α/β interaction, whereas ivermectin is not. Neither compound was able to inhibit the nuclear accumulation of telomere repeat factor-1 (GFP-TRF), which is known to be imported into the nucleus via IMP β1 alone, 7 with results indicating that although ivermectin is not specific for the IN-IMP α/β interaction, it does appear to be a specific inhibitor for cargoes dependent on the IMP α/β heterodimer for their nuclear import. The fact that neither compound was able to generally inhibit all nuclear transport in these experiments was a clear indication that the observed effects on inhibition of the nuclear import of specific cargoes could not be attributed to a nonspecific cytotoxic or other effect in the short 3-h incubation period used.
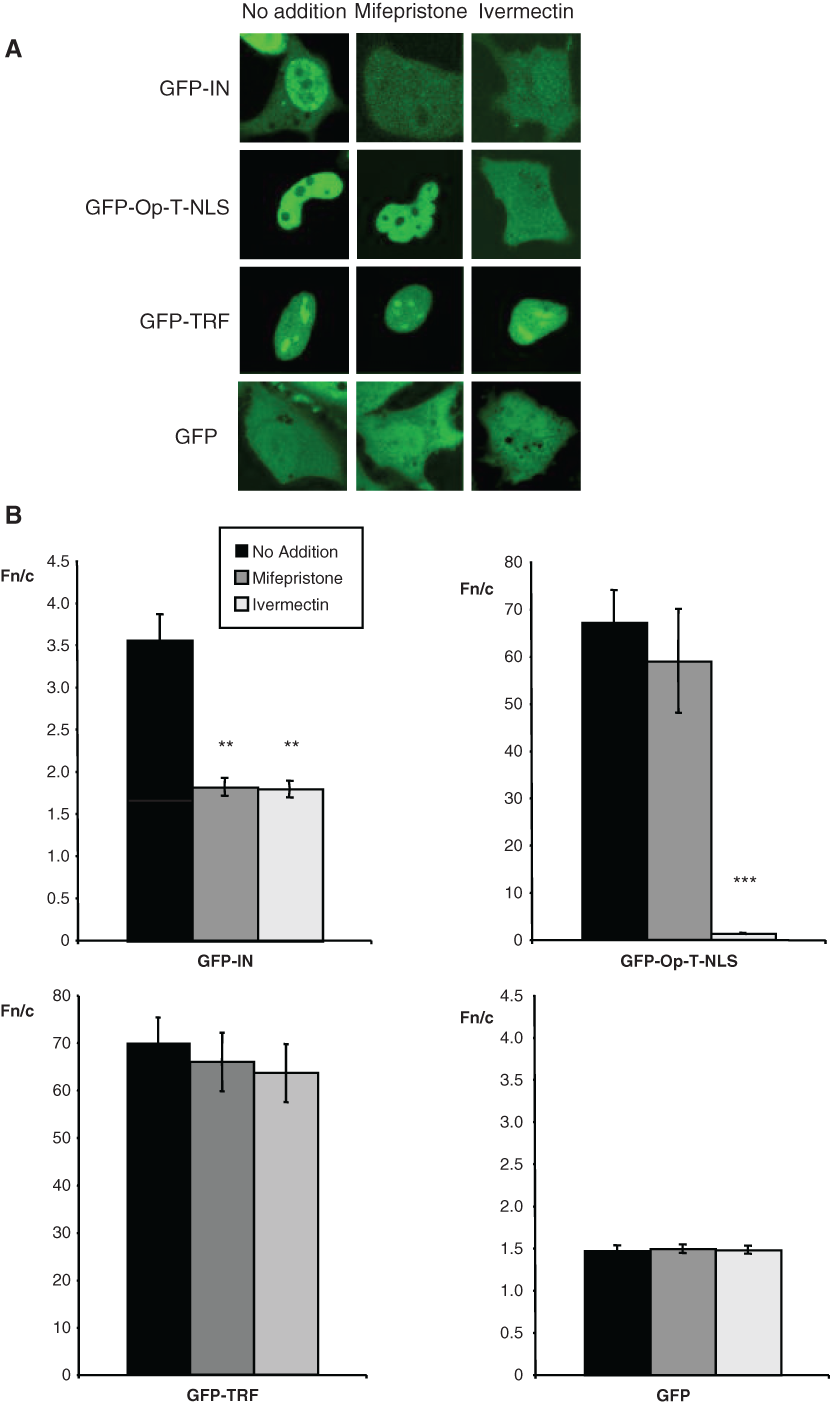
Mifepristone specifically inhibits integrase (IN) nuclear accumulation. (
Discussion
In this study, we investigated viral protein nuclear import as a potential target for antiviral intervention, describing a novel high-throughput assay for screening for inhibitors of protein nuclear transport and its use to screen for inhibitors of HIV-1 IN nuclear import, specifically for compounds that inhibit the binding of IN to its nuclear import receptor IMP α/β. Using a cross-screening approach, we were able to identify inhibitors specific for the IN-IMP α/β interaction and not other viral protein-IMP α/β interactions, as demonstrated in live-cell experiments. The assay is based on our previously described AlphaScreen® assay for examining protein-protein interactions, particularly NLS-IMP interactions. 20 The true power of this assay lies in the fact that it is focused on identifying inhibitors of a specific protein-protein interaction rather than compounds that bind to and inhibit a single protein activity. This enables compounds to be directed at a completely novel series of targets such as the interactions involved in viral protein nuclear import, which would otherwise be unsuitable for traditional therapeutic intervention; clearly, developing inhibitors that globally shut down cellular nuclear import pathways would not be generally desirable, but we have shown here that it is possible to identify compounds that inhibit IN nuclear import, without affecting T-ag or cellular proteins such as TRF-1. The fact that the assay is miniaturized (performed in 384-well microplates) also makes it ideally suited for HTS application, as indicated by the robust Z′ factor values. The assay was used to screen a small library for compounds that could effectively inhibit the binding of HIV-1 IN to IMP α/β. The screen was followed by screening for false-positive interactions, with several nonspecific compounds identified and removed from the assay, including 2 compounds containing imidazole rings, which actively compete with His6 for binding to the nickel-chelate acceptor beads, preventing any interaction from generating a signal. Clearly, the counterscreen assay can easily identify false-positive compounds contributing to the robustness of the screening regimen.
From 8 true hits, 5 were found to also actively inhibit the binding of IMP α/β to another well-characterized cargo, the Op-T-NLS, indicating that these compounds are not specific for the IN-IMP α/β interaction. These results demonstrate the utility of our approach to effectively separate hits into specific and nonspecific compounds, both of which may be useful. Specific hits are potential lead candidates for antiviral drug development, whereas nonspecific compounds can potentially be used in cell biology applications to enable the investigation of nuclear import pathways. The latter group is particularly intriguing as currently the only nuclear transport inhibitor in general use is LMB, 32,37,38 which specifically inhibits the nuclear export–mediating protein CRM-1 and has proven extremely useful in elucidating nuclear export signals/pathways for numerous proteins. In terms of nuclear import inhibitors, a single peptide (designated M9M) has been described through structure-based design 39 to bind to IMP β2 (transportin) and inhibit β2-mediated nuclear import, but at present, there are no commercially available inhibitors of nuclear import. Importantly, this study has identified ivermectin as a potential general IMP α/β-mediated nuclear import inhibitor, which does not appear to affect nuclear import mediated by IMP β1 alone. Although ivermectin is a broad-spectrum antiparasitic agent, which functions by modulating α7 sodium channels, the results here suggest it may also prove useful in future studies examining IMP α/β–mediated nuclear import.
Of the specific compounds identified in this study, mifepristone was demonstrated to inhibit the IN-IMP α/β interaction with µM affinity. Furthermore, mifepristone significantly reduced the nuclear accumulation of IN but not the Op-T-NLS or TRF-1 in living transfected cells, indicating that mifepristone selectively inhibits the IN-IMP α/β interaction, rather than IMP α/β–dependent nuclear import in general. Interestingly, the level of nuclear accumulation of IN in the presence of mifepristone demonstrated in this study is similar to the observed level of GFP (Fn/c ≈1.7; Fig. 4 ), which does not contain an NLS and only enters the nucleus through passive diffusion through the NPC. This implies that, although active IN nuclear import appears to be completely inhibited by mifepristone and ivermectin, there is no effect on the NPC itself, consistent with the fact that mifepristone does not affect other nuclear import pathways such as those for Op-T-NLS and TRF-1. Mifepristone is likely to have anti-HIV activity because IN is involved in nuclear import of the HIV-1 PIC, 18,19,40 which is well above the molecular weight cutoff for diffusion through the NPC, and nuclear import of the PIC is absolutely required for viral replication and long-term infectivity of HIV. 17,18,41
Finally, it is interesting to note that mifepristone (a competitive receptor antagonist of the progesterone receptor) also inhibits the glucocorticoid receptor, which has led to its investigation as an antiretroviral therapeutic agent due to the use of the glucocorticoid receptor by the HIV accessory protein Vpr during HIV infection. Mifepristone has been shown to significantly inhibit Vpr transactivation through the glucocorticoid receptor in a dose-dependent fashion, correlating with a strong dose-dependent antiviral effect, 42 and it is already being studied in early stage clinical trials, although with somewhat disappointing initial results. 43 That we have now demonstrated that mifepristone may have antiviral effects on a second HIV protein, IN, may prove useful in its further development as an antiviral therapeutic agent.
In conclusion, we have developed a highly sensitive and effective screening method, based on AlphaScreen® technology to identify inhibitors of IMP-NLS interactions, and have successfully identified several compounds that may prove useful in this regard. We have shown that one of these compounds, mifepristone, is a highly specific and effective inhibitor of HIV-IN nuclear import and may therefore serve as the lead compound for a new class of antiviral therapeutics against this novel yet overlooked import target of viral replication.
Footnotes
Acknowledgements
This work was supported by a grant from the NHMRC, Australia (#384109). The LOPAC1280 library was kindly donated by Dr. Roger Bosse (PerkinElmer-BioSignal, Montreal, Canada). The following reagents were obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, NIAID, NIH: pINSD.His.Sol was from Dr. Robert Craigie and pNL4-3 was from Dr. Malcolm Martin.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.