
Abstract
Lactococcus lactis subsp. lactis L3A21M1, which produces the bacteriocin lacticin 481, was investigated for its ability to improve cheese ripening. Three laboratory-scale Cheddar-type cheeses were produced from pasteurized cow's milk in triplicate trials: control cheese made with a commercial starter culture containing Lactococcus lactis subsp. lactis and Lactococcus lactis subsp. cremoris; Bac + cheese produced with the same commercial starter culture plus Lactococcus lactis subsp. lactis L3A21M1 as a producer of bacteriocin (lacticin 481); and Bac− cheese produced with the same commercial starter culture plus Lactococcus lactis subsp. lactis L3B1M7 as a non-bacteriocin producer. The microbiological, physicochemical, textural, proteolytic, and peptide profiles of cheeses were evaluated during the 28-day ripening period. The bacteriocin-producing Lc. lactis L3A21M1 increased proteolysis throughout the ripening process, as shown by the higher (p < 0.05) values for nitrogen fractions and free amino acid content in the cheeses. The textural properties were also influenced by the addition of the Lc. lactis culture, resulting in a softer cheese. The use of the bacteriocin-producing strain Lc. lactis L3A21M1, as an adjunct culture, thus accelerates the proteolysis of Cheddar cheese by inducing the lysis of the starter culture and increasing the content of free peptides and amino acids during ripening. In addition, the amount of hydrophobic peptides decreased significantly (p < 0.05) during ripening in the cheese with the bacteriocin producer.
Highlights
A bacteriocin-producing strain, Lactococcus lactis L3A21M1, was used as an adjunct culture in cheese-making.
The bacteriocin-producing strain significantly accelerated proteolysis and reduced cheese hardness.
This strain is a promising adjunct culture for accelerating cheese ripening and improving cheese quality.
Introduction
Cheese is one of the fermented milk-based foods, which is characterized by its different texture, aroma, and taste (Alhelli et al., 2021). Cheese ripening is a complex process that mainly involves glycolysis, lipolysis, and proteolysis and plays an important role in the development of flavor characteristics (Tekin and Hayaloglu, 2023). Because ripening is a slow and expensive process, the dairy industry is keen to shorten this period by accelerating the development of cheese flavor characteristics (Alhelli et al., 2021; Martinez-Cuesta et al., 2001). Proteolysis is a rate-limiting process in the ripening of most cheeses and plays an important role in the development of organoleptic properties by hydrolyzing bitter peptides and releasing free amino acids, which are precursors of aroma compounds (Courtin et al., 2002). The main strategies considered to accelerate cheese ripening are high ripening temperatures, the addition of exogenous enzymes or cheese slurries, and the use of modified starters or adjunct cultures (Fox et al., 1996). Lactic acid bacteria (LAB) are an important source of enzymes such as proteinases, peptidases, amino acid degradation enzymes, and esterases that convert the milk components contained in the curd into low molecular weight compounds (Coelho et al., 2022). Since most enzymes produced by LAB are intracellular, cell lysis favors the access of enzymes to their substrates and most likely accelerates cheese ripening. Therefore, an increase in starter cell lysis will increase the rate of proteolysis (Morgan et al., 1997). One way to improve cell lysis is to use bacteriocin-producing strains added to early ripening during cheese production (Garde et al., 1997; Martinez-Cuesta et al., 2001). Various groups of bacteriocin-producing LAB have been used, especially lactococci (Ávila et al., 2005, 2006; Martinez-Cuesta et al., 2001; O'Sullivan et al., 2003a). The use of bacteriocin-producing strains offers a number of advantages, as there are no legal barriers to their use, no special equipment is required, the distribution in the curd is homogeneous, and the bacteriocin can ensure the safety of the final product (Martinez-Cuesta et al., 2001).
The lantibiotic lacticin 481 is a bacteriocin produced by Lactococcus lactis subsp. lactis. This peptide contains 27 amino acids, including two lanthionines, a 3-methyllanthionine, and a 2,3-didehydrobutyrine. This bacteriocin has a broad spectrum and is effective against a variety of lactic acid bacteria and also against pathogenic bacteria (Piard et al., 1992).
In previous studies, a bacteriocin producer—Lactococcus lactis subsp. lactis L3A21M1, which produces lacticin 481—was isolated from a traditional Azorean cheese and was shown to be effective against other LAB (lactobacilli, Lactococcus sp., and Enterococcus sp.) and Listeria monocytogenes (Ribeiro et al., 2016). In the present study, the potential of this bacteriocin producer to accelerate proteolysis during the ripening of Cheddar cheese was investigated. The aim of this study was therefore to evaluate the influence of the lacticin 481-producing strain (Lc. lactis subsp. lactis L3A21M1) as an adjunct culture on the gross composition, texture, and enhancement of proteolysis during the ripening of Cheddar cheese.
Materials and methods
Microorganisms and culture preparation
Two lactic acid bacteria isolated from Pico cheese were used as adjunct cultures. Lactococcus lactis subsp. lactis L3A21M1 (GeneBank KF193424) as a bacteriocin producer (lacticin 481) and Lactococcus lactis subsp. lactis L3B1M7 (GenBank KM079358) as a non-bacteriocin producer. A commercial starter consisting of Lc. lactis subsp. cremoris and Lc. lactis subsp. lactis, was used (DVS® R-703, CHR HANSEN, Denmark). Lc. lactis subsp. lactis L3A21M1 and Lc. lactis subsp. lactis L3B1M7 were initially cultured in MRS broth (Biokar, Germany) at 30 °C overnight for culture reactivation. Before adding the cultures for cheese-making, the overnight cultured strains were washed with PBS and added to reconstituted skim milk (10% w/v) and incubated at 30 °C for 24 h.
Laboratory-scale cheese-making
Bovine milk, 11 L (University of the Azores farm, Angra do Heroísmo, Portugal), was pasteurized at 65 °C for 30 min and cooled to 32 ± 2 °C. Cheddar-type cheeses were made according to Silva et al. (2023a). Three treatment cheeses were produced in each experiment: control cheese (starter only), cheese with the addition of a bacteriocin-producing culture—Lc. lactis subsp. lactis L3A21M1 (Bac+) and cheese with a non-bacteriocin-producing culture—Lc. lactis subsp. lactis L3B1M7 (Bac−), in three replicate experiments (blocks). To produce the control cheese, pasteurized milk was inoculated with the mesophilic starter culture (0.1U/L, DVS®R-703, CHR Hansen, Denmark). For the other cheese treatments, pasteurized milk was inoculated with the starter culture and the bacteriocin-producing culture Lc. lactis subsp. lactis L3A21M1 (Bac+) or the non-bacteriocin-producing culture Lc. lactis subsp. lactis L3B1M7 (Bac−). The Lc. lactis cultures were first activated in 100 mL of reconstituted milk and added at a constant bacterial concentration of ∼109 CFU/mL, with an inoculation ratio of 1%, resulting in an initial adjunct inoculation level of ∼107 CFU/mL of milk. The cultured milk was kept for 45 min at 32 ± 2 °C, curdled with 3.3 mL of animal rennet (Lusocoalho, Castelo Branco, Portugal), and left for 45–60 min to allow curd formation. The curd was then cut into ∼2 cm cubes and cooked at 37 ± 2 °C for 45 min. The curd was placed on a cheesecloth and pressed by hand for 20 min to remove most of the whey. The curd was salted (2% w/w) using sea salt, added manually and mixed uniformly, placed in cheese molds (400 g of curd into molds of 100 mm diameter and 40 mm high), and pressed for 15 min on each side, before moving to the maturation chamber. The cheeses were matured for 28 days at 10 °C–13 °C and 65%–70% RH. Samples were tested after 7, 14, 21, and 28 days of ripening. On each trial day, control and treatment cheeses were made from the same batch of milk. Four cheeses were made for each treatment, and the entire experiment was repeated three times independently, resulting in a total of 36 Cheddar-type cheeses.
Microbiological analysis and bacteriocin activity
Viable counts of Lactococcus sp. were determined in the different cheeses (control, Bac+, and Bac−) after 7, 14, 21, and 28 days of ripening. Cheese was grated, and 10 g was homogenized with 90 mL of peptone water (Biokar Diagnostics, France). Serial dilutions were plated on M17 agar (Biokar, Germany) and incubated at 30 °C for 48 h for lactococcal enumeration. Cell counts were expressed as log CFU g−1.
The antimicrobial activity of cheeses containing the bacteriocin-producing strain (Bac+) was evaluated using the agar well diffusion assay according to Ribeiro et al. (2014). The water-soluble extracts (WSE) were passed through a 0.22 µm sterile membrane filter before the well diffusion assay. WSEs obtained from the cheeses after 7, 14, 21, and 28 days were used to determine bacteriocin activity against the indicator strain Listeria monocytogenes 7644. Inhibition was recorded as positive when a translucent halo zone was observed around the well. All tests were performed in duplicate.
Chemical determinations
Cheese pH was measured in duplicate with a Crison penetration electrode (model 52-3,2; Crison Instruments, Barcelona, Spain). Titratable acidity (TA) was determined as g lactic acid (LA)/100 g (1 mL of 0.1 M NaOH equals 0.0090 g lactic acid) according to Kirk and Sawyer (1991). Fat was determined in cheese by the Gerber method (Kirk and Sawyer, 1991). Moisture content was determined by drying a sample (3 g) at 103 °C in an incubator (Memmert, Germany) for 24 h. Results were expressed as a percentage (g/100 g cheese) (AOAC, 1980). All analyses were performed in duplicate.
Hardness determinations
The hardness of the cheese samples was determined as fracturability (breaking force, expressed in N) after 7, 14, 21, and 28 days of ripening using a texturometer (TMS-PRO, Food Technology Corporation, USA), according to the method described by Primo-Martínet al. (2008) and Silva et al. (2023b). Cheeses were cut into blocks with dimensions 70 × 30 × 30 mm and were equilibrated at room temperature (20 °C) for 1 h before testing. The cheese blocks were fractured using a wedge-shaped aluminum probe (30° cutting angle and 25 mm wide) on a flat table at a speed of 100 mm/min. The penetration depth of the probe into the sample was set for 15 mm. All experiments were performed in triplicate.
Assessment of proteolysis
Proteolysis was assessed by analyzing the nitrogen fractions and the free amino acid content in the water-soluble nitrogen (WSN) fraction of the cheeses after 7, 14, 21, and 28 days of ripening.
In brief, water soluble extracts (WSEs) were prepared by homogenizing 50 g of grated cheese in 100 mL of Milli-Q water using a stomacher (5 min at 260 r/min, 400 Circulator Lab Blender, Seward, UK), followed by incubation at 40 °C for 60 min and centrifugation (30 min, 4000 ×g, 4 °C, Beckman Coulter J2-HC, Germany). The fat layer was removed, and the supernatant was filtered through Whatman No.1 paper. To 15 mL WSE, 5 mL 48% (w/v) TCA was added (12% TCA-SN). After 30 min at room temperature, the mixture was filtered through Whatman No. 42 paper. To 20 mL WSE, 14 mL 3.95 M H2SO4, and 6 mL 33.3% (w/w) phosphotungstic acid were added. The mixture was left overnight at 4 °C before being filtered through Whatman No. 42 paper. The nitrogen content of all fractions was determined by the Kjeldahl method using a Kjeltec System 2300 distillation apparatus (Tecator Technology, Foss, Spain) and expressed as a percentage of total cheese nitrogen.
The total concentration of free amino acids (FAAs) in the WSE was determined by the method of Radeljević et al. (2013). Results were expressed as grams of Leu/100 g of cheese. The standard curve of L-leucine (Fluka, Germany) was constructed, and the concentration of free amino acids in the samples was expressed as mM leucine.
Peptide analysis by HPLC
Hydrophilic and hydrophobic peptides were determined in the WSE by reversed-phase HPLC (VWR Hitachi, model Elite Lachrom, Japan) equipped with an automated injector, an RP-C18 column (Purospher® STAR, 5 μm particle size, 120 Å, 250 mm × 4 mm i.d.), and a diode array detector (DAD), according to the method described by Pavia et al. (2000). The separations were carried out at a flow rate of 1 mL min−1 with eluent A for 10 min and a linear gradient from 0% to 80% of eluent B for 80 min. Eluent A was 0.1% trifluoroacetic acid in water, and eluent B was 0.1% trifluoroacetic acid in acetonitrile and water (60:40, v/v). After running the samples, the integration area of the peptides (Abs 220 nm) was determined. The area of peptides eluted between 10 and 35 min was considered as the hydrophilic peptide area, while the area of peptides eluted between 35 and 80 min was considered as the hydrophobic peptide area (Gonzalez de Llano et al., 1995). The amounts of hydrophobic and hydrophilic peptides were expressed as units of chromatogram area.
Statistical analysis
At least three experiments (batches) were performed, and results were expressed as mean values ± standard error of the mean (SEM). A split-plot analysis of variance (ANOVA) design was used for comparisons between cheeses in each ripening time, with cheese treatments (control, Bac+, and Bac−) and ripening duration (days 7–28) as factors. For the chemical and texture analysis, a one-way ANOVA was used to assess the differences between the cheese treatments. When statistically significant differences were found (p < 0.05), the post-hoc Tukey test was used to distinguish between the cheese treatments. Statistical tests were performed using IBM SPSS Statistics, version 28 (IBM Corporation, New York, USA).
Results and discussion
Microbiological analysis and bacteriocin activity
The CFU of Lactococcus sp. in the different treated cheeses is shown in Figure 1. As observed, the Lactococcus counts were similar (p > 0.05) in all cheese treatments with values between 7 and 8 log CFU g−1. Levels of these bacteria remained > 7 log CFU g−1 throughout ripening in all cheeses. These results are in agreement with those of other authors who have observed the survival and maintenance of LAB counts during ripening (Garde et al., 2006; Kocak et al., 2020). Although the media used to determine CFUs could not distinguish between the Lc. lactis starter strains and the bacteriocin-producing strain (Lc. lactis subsp. lactis L3A21M1), the bacteriocin was detected in the cheese inoculated with this strain on days 7, 14, 21, and 28 (data not shown). WSEs from all cheese treatments were tested under identical conditions, and inhibition was observed exclusively in the cheese inoculated with the bacteriocin-producing strain. Despite the presence of bacteriocin (lacticin 481), the number of Lactococcus in this cheese was slightly higher and maintained throughout the ripening period. Some authors observed that lacticin 481 has a bacteriostatic mode of action on lactococcal cultures, with some cells suffering gradual lysis, whereas other cells continued to grow (O'Sullivan et al., 2002). In the present work, this particular mode of action of lacticin 481 would explain the growth of Lactococcus sp. in cheese with simultaneous activity of the bacteriocin during the first week. It should be noted that bacteriocin activity was evaluated from day 7 onwards, and samples from the early fermentation stage (day 0–1) were not analyzed in this study. Previous studies have shown that bacteriocins produced by Lactococcus lactis are actively excreted and reach their highest expression and activity during the early stages of cheese manufacture, followed by a decline during ripening due to reduced gene expression, diffusion limitations, adsorption to the cheese matrix, or proteolytic degradation (Noutsopoulos et al., 2017). In this context, the pronounced proteolytic activity observed in Bac+ cheeses at day 7 may reflect bacteriocin accumulation during early fermentation, which could have promoted early starter cell lysis and subsequent release of intracellular enzymes before the first sampling point.
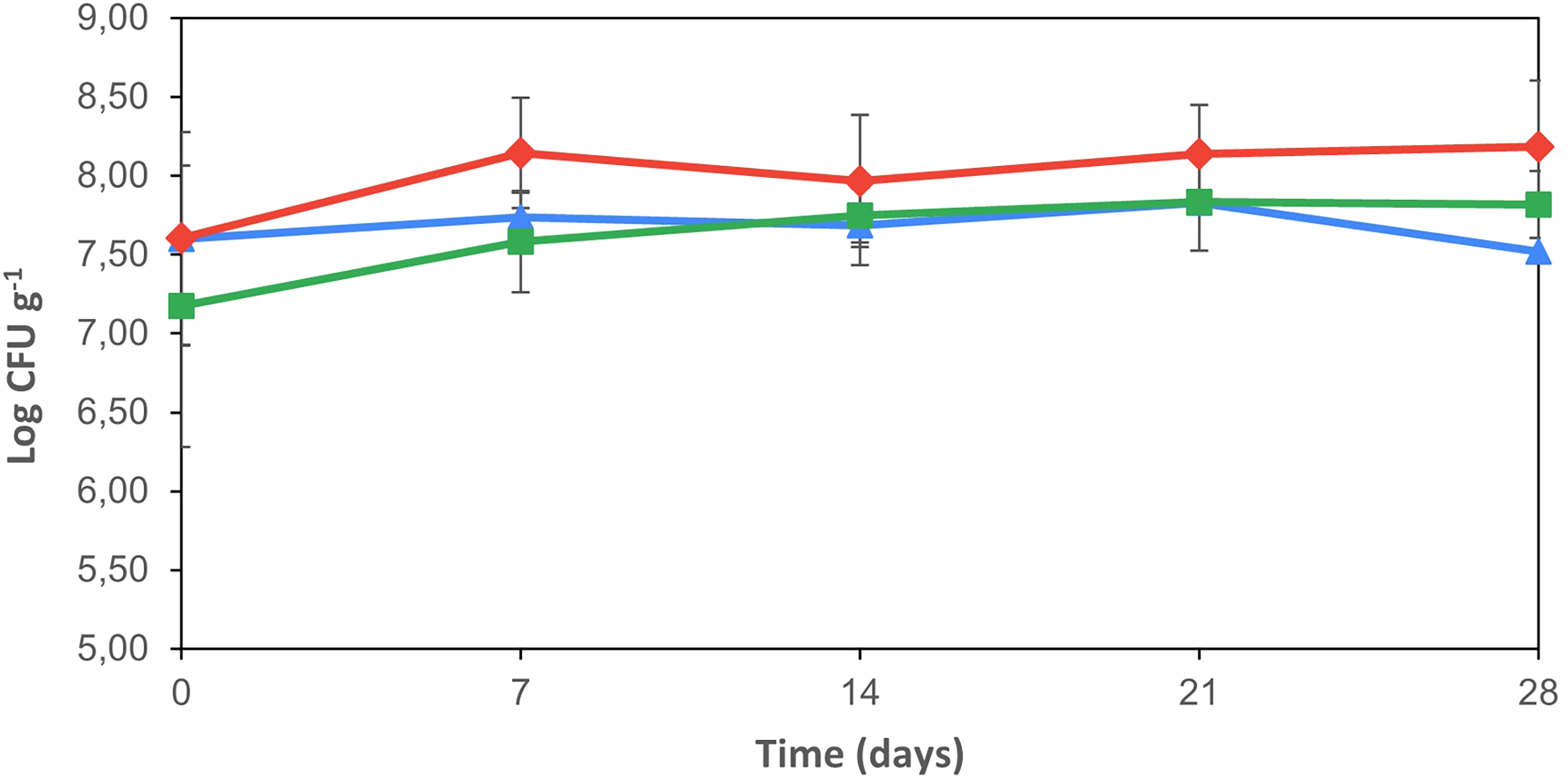
Counts of lactococci in the cheeses made with a commercial starter culture (control cheese Δ), commercial starter culture, and Lc. lactis L3A21M1 (bacteriocin-producing strain bac 481 ♦) and commercial starter and Lc. lactis L3B1M7 (no bacteriocin-producing strain ∎) over 28 days of ripening. Values are the means of three replicates, and error bars represent the standard error of the mean (SEM).
Chemical determinations
Cheese is a biochemically dynamic product and undergoes considerable changes during its ripening process (McSweeney and Sousa, 2000). The changes in the chemical composition during ripening at different ripening times are shown in Table 1. The pH of the cheeses was influenced by the treatment and ripening time of the cheese. The addition of a second Lactococcus strain resulted in an increase in more pronounced acidification (p < 0.05) with the bacteriocin producer (L3A21M1), suggesting that this strain is also contributing to the acid production. The pH values of all cheeses also decreased significantly (p < 0.05) from the first day to the end of ripening. Other authors found that the addition of bacteriocin-producing cultures can retard acid production by the starter culture, increasing the time of acidification (Garde et al., 2002; Morgan et al., 1997). This is not the case in the present study, since the addition of bacteriocin-producing culture (Bac+) increased acidification kinetics (Table 1). In addition, acid production (TA) was slightly higher for the cheese with the added bacteriocin producer (Bac+) and remained constant over the ripening period. Therefore, the inclusion of the lacticin 481-producing strain did not compromise the acid production by the starter as reported with the use of other bacteriocin producers (O'Sullivan et al., 2003a). Differences in fat-in-dry-matter (FDM) among treatments indicate variations in fat retention within the cheese matrix. FDM values observed in cheeses produced with the addition of a second Lactococcus strain (Bac+ and Bac−) are consistent with the reduced brittleness measured in these cheeses. In particular, Bac+ cheeses showed the highest FDM, which, together with the extensive proteolysis observed, suggests that both compositional and biochemical changes contributed to the softer texture. As expected, DM increased during the ripening period, but there were no significant differences (p > 0.05) between the cheese treatments. These results were also observed by other authors (Garde et al., 2006). However, in cheeses to which a second Lactococcus culture (Bac+ or Bac−) was added, the protein content was slightly reduced compared to the control (p < 0.05), while the fat content increased (p < 0.05). One possible explanation for these differences could be related to the higher amount of acid produced by the addition of the second Lactococcus culture. The acidification helps to trap the fat globules in the casein network so that more fat is retained by the casein, resulting in cheeses with a higher fat content. As the moisture content remained approximately the same, this was offset by a lower protein content. As this effect was observed in both bacteriocin-producing (Bac+) and non-bacteriocin-producing (Bac−) cheeses, it cannot be attributed to the production of bacteriocin itself.
Composition of cheeses produced with a commercial starter culture (control), commercial starter culture, and Lc. lactis L3A21M1 (Bac+), and commercial starter and Lc. lactis L3B1M7 (Bac−).

Values are the mean of three replicates ± SEM.
Different lowercase letters as superscripts in the same column indicate significant differences (p < 0.05) between ripening time (days) within each cheese treatment. Different uppercase letters as superscripts in the same column indicate a significant difference (p < 0.05) between cheese treatments at the same ripening time.
Cheese hardness
The hardness of the cheeses was assessed on the basis of the fracturability (breaking force) and is shown in Figure 2. The fracturability increased significantly (p < 0.05) in all cheese samples after 14 days of ripening (p ˂ 0.05). Similarly, other authors have found that semi-hard cheeses become harder during ripening due to moisture loss (Ávila et al., 2006). The addition of a bacteriocin-producing culture (Bac+) or a non-bacteriocin-producing culture (Bac−) resulted in a softer cheese with a significant (p < 0.05) reduction in brittleness on the 14th and 28th days of ripening. This reduction is probably due to the increase in total fat in these cheeses, observed on the 28th day of ripening (Table 1).
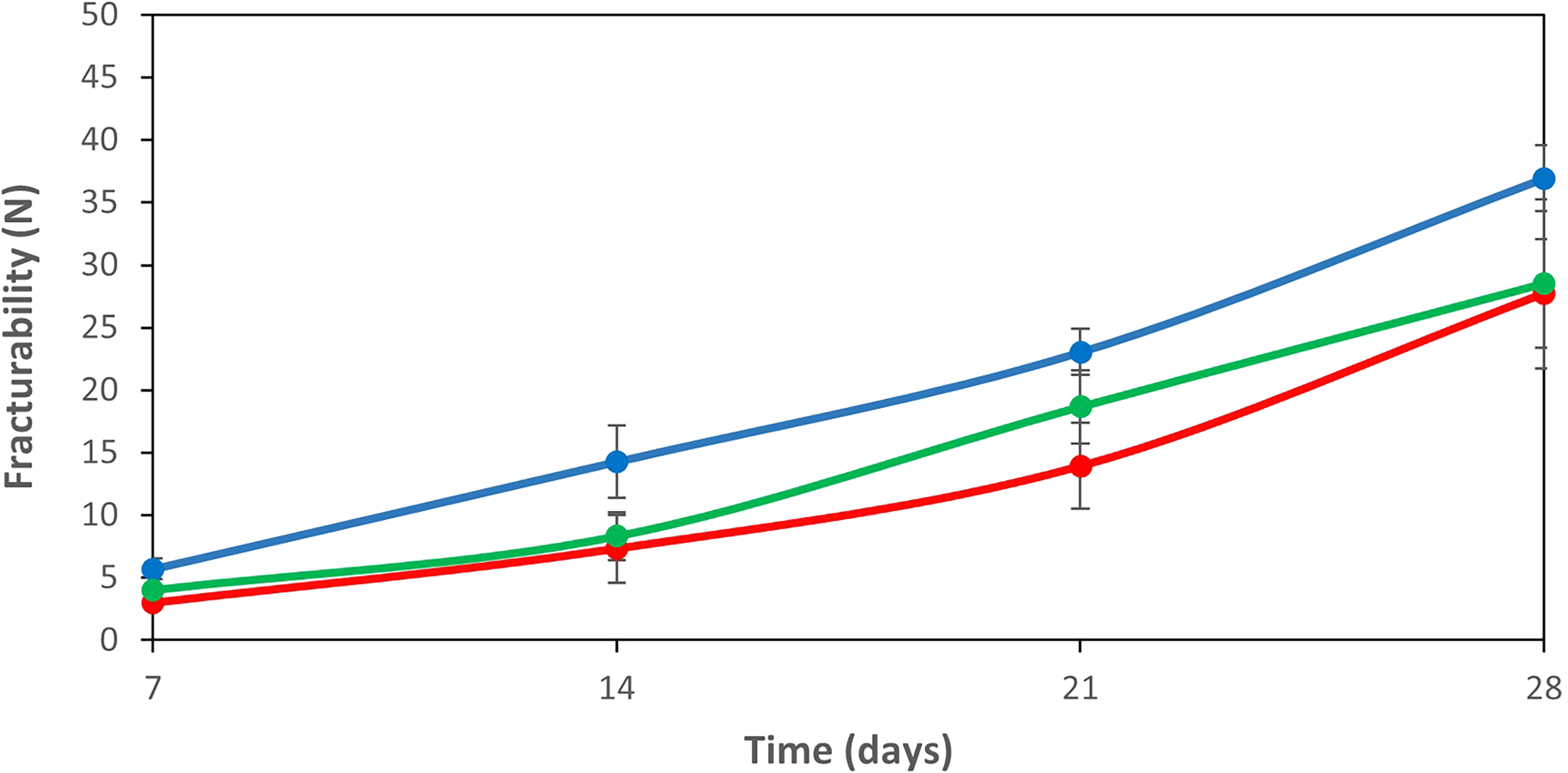
Fracturability (breaking force) of cheeses made with a commercial starter culture (control cheese—blue line), commercial starter culture, and Lc. lactis L3A21M1 (Bac+, bacteriocin-producing strain—red line) and commercial starter and Lc. lactis L3B1M7 (Bac−, no bacteriocin producer—green line). Values are the means of three replicates, and error bars represent the standard error of the mean (SEM).
Assessment of proteolysis
The WSN content in cheeses with the bacteriocin-producing strain (Bac+) was significantly higher (p < 0.05) compared to cheeses with the non-bacteriocin-producing strain (Bac-) or control cheeses. On day 7, the WSN concentration as a percentage of total N in Bac+ cheese was 7.51%, while control and Bac− cheeses had lower values of 2.67% and 4.38%, respectively (Figure 3(a)). These results showed that the lacticin 481-producing strain influenced primary proteolysis, probably by enhancing cell lysis and the release of intracellular enzymes of the starter culture. As reported previously (Ribeiro et al., 2016), the bacteriocin 481 exhibited antimicrobial activity against the species Lc. lactis subsp. lactis and Lc. lactis subsp. cremoris. However, the contribution of strain-specific physiological properties unrelated to bacteriocin production cannot be excluded (Sheehan et al., 2006).
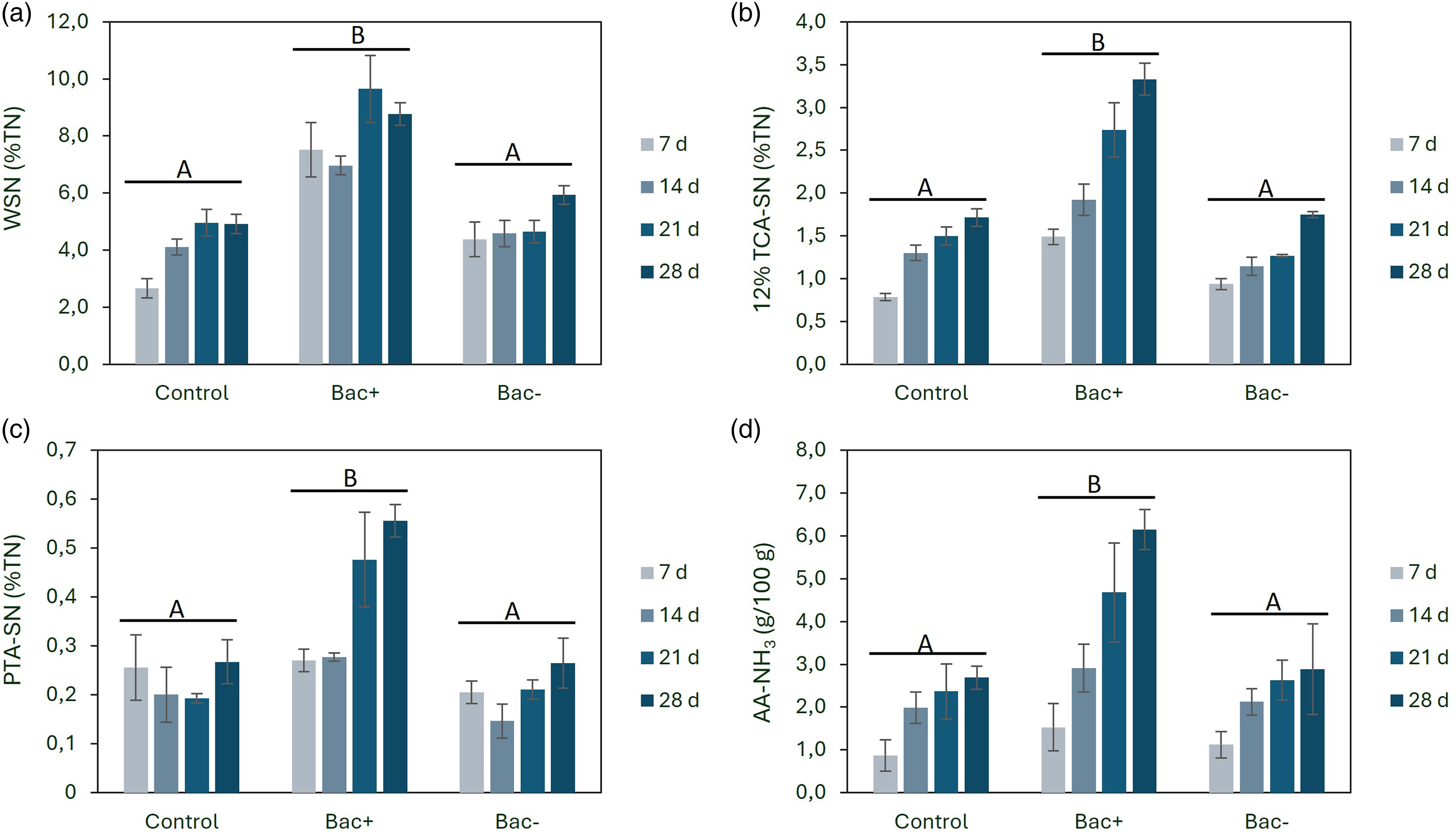
Age-related changes in levels of proteolysis measured as soluble nitrogen (N) in (a) WSN, (b) TCA 12%, (c), PTA, and (d) free amino acids (AA-NH3) in cheese made with starter only (control), cheese made with starter and Lc. lactis L3A21M1 (Bac+), and cheese made with starter and Lc. lactis L3B1M7 (Bac−) after 7, 14, 21, and 28 days of ripening. Values are the means of three experiments, and error bars represent the SEM. Different capital letters indicate a significant difference (p < 0.05) between the cheese treatments.
Proteolysis is considered one of the most important biochemical processes during cheese ripening, in which proteins are broken down to primary products and then to secondary products such as small and medium peptides and finally to free amino acids (Kongo et al., 2009; Sousa et al., 2001). Several factors influence proteolysis indices, including microbial activity and technological parameters such as rennet dosage, pH evolution, and salt content. Salt improves cheese texture by modifying it to a sticky texture in the hydrated state. Additionally, salt regulates the proliferation of microorganisms and enzyme activity (Bae et al., 2017). pH can also affect the type and extent of proteolysis by altering protein interactions, which in turn affects cheese functionality. The effect of pH on early cheese maturation has been shown to influence total and soluble calcium content, moisture content, the extent and pattern of proteolysis, and their interactions (Pastorino et al., 2003). Some authors have demonstrated that lower pH generally makes cheese softer, more crumbly, and less elastic by increasing casein protein solubility, while higher pH leads to firmer, more solid-like textures during early cheese maturation (Ramkumar et al., 1998). Coagulants are essential in cheese manufacture, as they initiate milk coagulation, with animal rennet traditionally being the most widely used option. Key findings indicate variations in coagulation patterns across the coagulants used, and the concentration levels affected the coagulation pattern, with a clear linear increase in dose influencing gelation and varying effects on other coagulation traits (Stocco et al., 2025). Although proteolysis indices may be affected by these technological parameters, these were kept constant across treatments in the present study, suggesting that the observed differences primarily reflect treatment-related effects.
The WSN, the nitrogen soluble in 12% trichloroacetic acid (12% TCA-SN), the nitrogen soluble in 5% phosphotungstic acid (5% PTA-SN), and the free amino acid content determined by the Cd-ninhydrin method are commonly used to characterize the proteolysis of cheese (McSweeney and Fox, 1997). Primary proteolysis was measured using the first two methods, while the degree of secondary proteolysis was quantified by measuring low molecular mass peptides (<15 kDa) and free amino acids using the last two methods.
According to several authors (McSweeney and Fox, 1997), the 12% TCA-soluble N content is a better index of maturity than WSN. In the present study, the level of 12% TCA-soluble N increased significantly (p < 0.05) in all cheeses during the ripening period (Figure 3(b)). The addition of the bacteriocin-producing strain (Bac+) also led to a doubling of the 12% TCA-soluble N content compared to the other cheeses (control and Bac−). These results suggest that the addition of a bacteriocin-producing strain has a marked effect on primary proteolysis by increasing the production of medium and small peptides.
The PTA-soluble N gives an estimate of the content of low molecular weight peptides, free amino acids, and other smaller nitrogen-containing compounds such as urea, amines, and ammonia. In the cheese with the bacteriocin-producing strain (Bac+), the PTA-soluble N showed a marked increase on days 21 and 28 and was twice the value of the other cheeses (Figure 3(c)). Therefore, the addition of the bacteriocin-producing strain has a marked influence on the release of small peptides, amino acids, and other smaller nitrogen-containing compounds during cheese ripening.
The Cd-ninhydrin method is very sensitive for estimating the content of free amino acids in cheese (AA-NH3 content). During ripening, the concentration of free amino acids increased independently of the cheese treatment (Figure 3(d)). However, the concentration of free amino acids was significantly higher in the cheese with the bacteriocin-producing strain (Bac+). Overall, these results show that the addition of the bacteriocin-producing strain (bacteriocin 481) increases the primary and secondary proteolysis of the cheese from the beginning of ripening. The results differ from the study by Ryan et al. (1996), who used a strain of Lc. lactis, which produces the bacteriocin lacticin 3147, as a starter in the production of Cheddar cheese. The authors found that the use of this bacteriocin producer reduced the levels of non-starter lactic acid bacteria but did not change the proteolysis of Cheddar cheese (WSN, AA-NH3 content, and free amino acid composition). In contrast, Ávila et al. (2007) showed that proteolysis (estimated by free amino acid levels) was significantly higher in Manchego cheese inoculated with the strain Lc. lactis subsp. lactis 415 (nisin Z and lacticin 481 producer). The authors also found that the textural and sensory properties of Manchego cheese were not significantly affected by the use of the bacteriocin-producing strain. These observations suggest that the impact of bacteriocin-producing strains on cheese proteolysis depends on the specific bacteriocin and the cheese matrix. Lacticin 481 is a small, heat-stable lantibiotic with a primarily bacteriostatic mode of action and a medium spectrum of inhibition, mainly active against other lactic acid bacteria. This mode of action may favor the controlled release of intracellular enzymes during early ripening, thereby enhancing proteolysis without drastically reducing starter viability. In contrast, lacticin 3147 has a broad inhibitory spectrum and is active against a wide range of Gram-positive bacteria, which may limit enzyme release or alter microbial dynamics differently (O'Sullivan et al., 2003b). Additionally, differences in cheese variety, milk composition, starter culture composition, and ripening conditions are likely to influence the extent to which bacteriocin-associated effects on proteolysis are expressed.
RP-HPLC peptide profiles
The analysis of peptide profiles by HPLC can be used to characterize the type of cheese and provides valuable information about proteolysis during cheese ripening (Vivar-Quintana et al., 2009). The average peak areas of the water-soluble extracts of the cheeses during the 28-day ripening period are shown in Figure 4. The analysis of the total peptide profiles shows an increase (p < 0.05) in peptides during the ripening period (control and Bac−) (Figure 4(a)). This tendency was not observed in the Bac+ cheese (bacteriocin-producing strain). Here, the amount of peptides was higher in the first 2 weeks of ripening (7 and 14 days) and decreased by half in the last weeks to reach values similar to the other cheese treatments. These results are consistent with the results of the WSN, where primary proteolysis was higher in cheeses with the bacteriocin-producing strain during the first weeks of ripening.
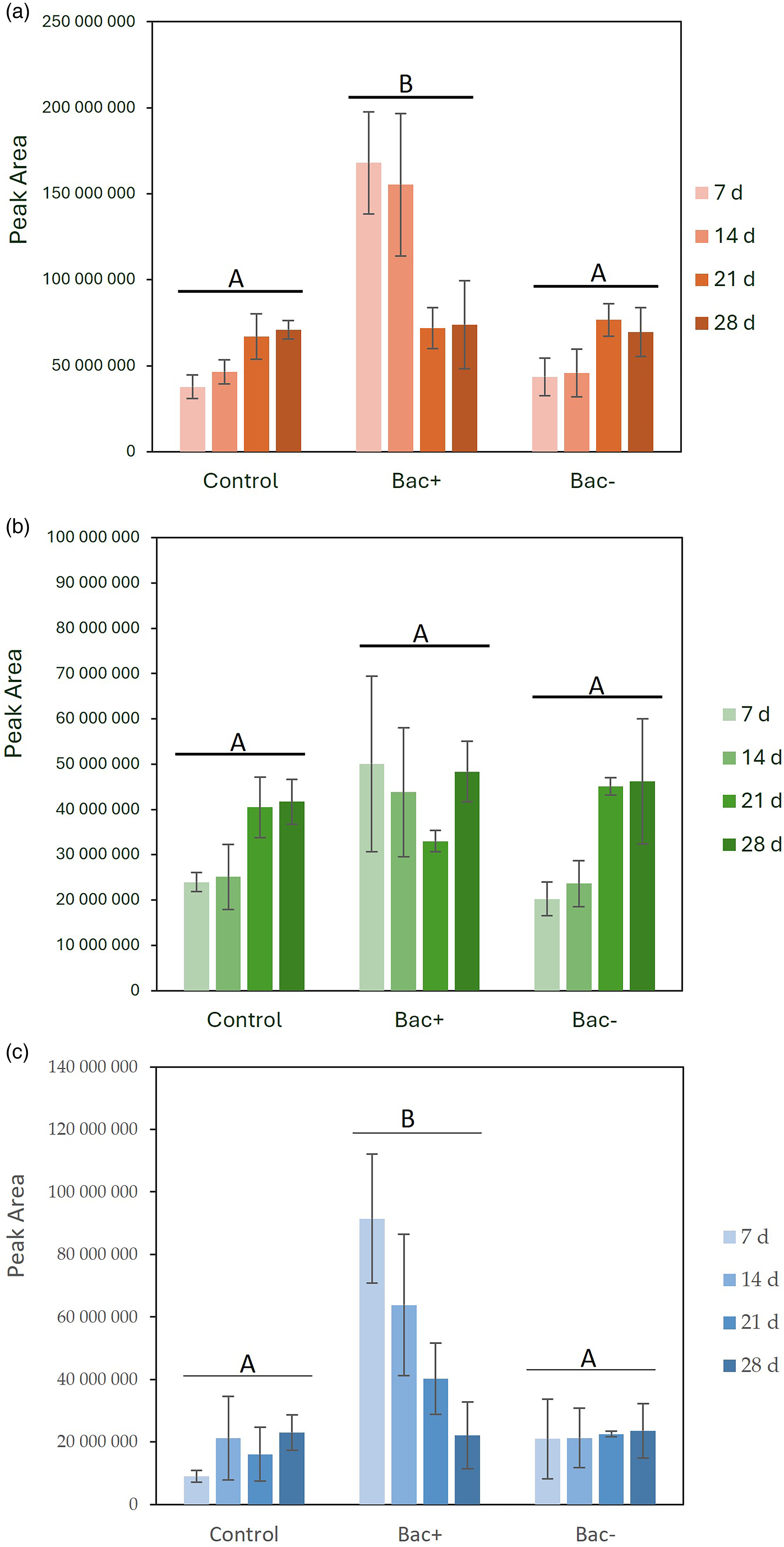
RP-HPLC peptide profiles of water-soluble fractions of cheese made with starter only (control), cheese made with starter and Lc. lactis L3A21M1 (Bac+), and cheese made with starter and Lc. lactis L3B1M7 (Bac−) after 7, 14, 21, and 28 days of ripening. (a) Total peptides, (b) hydrophilic peptides, and (c) hydrophobic peptides. Values are the means of three experiments, and error bars represent the SEM. Different capital letters indicate a significant difference (p < 0.05) between the cheese treatments.
The cheese peptides were separated according to their hydrophobicity, with the hydrophilic peptides eluting first and the hydrophobic peptides eluting later (De Llano et al., 1995). Overall, no significant differences (p > 0.05) in hydrophilic peptides were observed between the cheese treatments (Figure 4(b)). However, the cheese with the bacteriocin-producing culture (Bac+) showed a different pattern during ripening. While the hydrophilic peptide content of the control and Bac− cheeses increased from the second to the third week of ripening, the Bac+ cheese showed a high level of hydrophilic peptides from the beginning of ripening, which was maintained throughout the ripening time. As for hydrophobic peptides, although they were low in the control and Bac− cheeses and remained fairly constant, in the Bac+ cheeses, they were considerably higher (p < 0.05) from the beginning of ripening (Figure 4(c)), confirming the high level of proteolysis caused by the addition of the bacteriocin-producing culture. It is noteworthy that in these cheeses (Bac+), the amount of hydrophobic peptides decreased significantly (p < 0.05) during ripening, reaching similar levels to the control and Bac− cheeses on day 28. These results are in agreement with the previous study by Garde et al. (2006), which applied a lacticin 481-producing strain on a semihard Spanish cheese made with pasteurized cows’ milk and found a lower content of hydrophobic peptides. There are several factors that may contribute to casein breakdown and hydrophobic peptide release, such as the amount of rennet, temperature, curdling time, and addition of salt to the curd before molding (De Llano et al., 1995). As several researchers have found with different types of cheese, the peptide content increases with increasing age of the cheese, while the ratio of hydrophobic and hydrophilic peptides decreases (Garde et al., 2006). In the present study, the increase in peptide content during ripening was only observed in control and Bac− cheeses, as Bac + cheeses exhibited extensive primary proteolysis from the beginning of ripening. During the maturation of this cheese, primary proteolysis gave way to secondary proteolysis, and the content of hydrophobic peptides decreased as they were converted into free amino acids. Hydrophobic peptides were associated with the bitterness of the cheese by several authors (Garde et al., 2006; Morgan et al., 1997; Oumer et al., 2001). Therefore, although no sensory evaluation of these cheeses was performed, it was important that the hydrophobic peptides were reduced during the ripening of the Bac + cheeses and reached similar levels to the control cheeses. In addition, the higher level of proteolysis observed in Bac + cheeses could improve the sensory properties of these cheeses, as several authors have shown that a higher level of proteolysis correlates with a higher flavor intensity in cheese (Hayaloglu et al., 2005). For example, Garde et al. (2002) showed that cheese made with bacteriocin-producing Lc. lactis subsp. lactis as an adjunct culture resulted in significantly higher amino acid content and high scores of both flavor quality and intensity. Moreover, with the production of bacteriocins, the growth of non-starter LAB and pathogenic bacteria (e.g. Listeria monocytogenes) may be controlled, thus reducing the risk of off-flavor development and improving safety. Nevertheless, any implications regarding improvements in flavor quality remain speculative, and rand further studies on sensory and textural evaluation are required to confirm the observed results.
Conclusions
The results of this study suggest that the combination of a mixed starter culture of Lc. lactis (commercial starter mixture of Lc. lactis subsp. cremoris and Lc. lactis subsp. lactis) and the strain Lc. lactis subsp. lactis L3A21M1, a lacticin 481 producer, optimizes the release of intracellular components in the first days of ripening and thus influences the process of proteolysis of Cheddar cheese without altering its texture. The addition of this bacteriocin producer represents a cost-effective method for accelerating the ripening process of cheese. Further studies comparing cheeses produced with this bacteriocin-producing starter adjunct at reduced ripening times to conventionally ripened Cheddar cheeses are required to determine whether comparable compositional, textural, and sensory attributes can be achieved. The results of this study extend the knowledge about the changes during cheese ripening by the addition of a bacteriocin-producing strain, which could be of interest to the dairy industry.
Footnotes
Acknowledgements
This work was financially supported by the Portuguese Foundation for Science and Technology (FCT), Project UIDB/00153/2020. SC Ribeiro gratefully acknowledges financial support from the Portuguese Foundation for Science and Technology (FCT), Project UIDP/00153/2020.
Author contributions
Susana C Ribeiro: writing–original draft, methodology, investigation, supervision, formal analysis, and conceptualization. Agathe Blackway: methodology and investigation. Sofia PM Silva: methodology and investigation. Dominika Jurášková: methodology and investigation. Cristiana Maduro Dias: methodology. Célia CG Silva: writing–review and editing, supervision, formal analysis, and conceptualization.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Fundação para a Ciência e a Tecnologia (grant number UIDP/00153/2020).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Data will be made available on request.