
Abstract
Endothelial protein C receptor (EPCR) is a candidate mediator in the pathogenesis of thrombosis, as several data in the literature indicate that polymorphisms such as EPCR 4678G/C and 4600A/G are associated with either protective effect or increased risk of thrombosis, respectively. We investigated the prevalence of these polymorphisms in patients with thrombotic disorders as well as their impact on the risk of thrombosis, the age of first thrombotic episode, and recurrence. The prevalence of the rare EPCR alleles 4600G and 4678C was comparable in patients and controls. However, in a subset analysis, we observed that 4600G allele was more prevalent among patients who developed thrombosis at younger age (<35 years). Moreover, the prevalence of 4678C allele was significantly lower in younger patients compared to older patients. Neither polymorphism seemed to have an impact on recurrence regardless of age. Soluble EPCR levels were elevated in 4600AG patients compared to controls while 4678CC patients presented with lower levels of soluble form of EPCR compared to carriers of at least 1 4678G allele. Our data suggest that either the lack of the protective EPCR 4678C allele or the presence of EPCR 4600G allele may be associated with earlier development of thrombosis.
Introduction
The protein C (PC) pathway is a natural anticoagulation system that plays a major role in maintaining the hemostatic balance and depends on the precise assemblage and interplay of at least 5 proteins: thrombin, thrombomodoulin (TM), PC, protein S (PS), and endothelial protein C receptor (EPCR) on the surface of vascular endothelial cells that result in generation of activated protein C (APC). Protein C is a vitamin K-dependent plasma glycoprotein, which is activated on the endothelial surface when thrombin binds to TM. Activated PC, with its cofactor PS, inactivates coagulation factors Va and VIIIa, thereby reducing thrombin generation. 1
Endothelial PC receptor, a transmembrane protein expressed on the endothelium of large vessels, is also involved in the activation of PC anticoagulant pathway by binding PC and enhancing its activation. 2 It increases 5-fold the activation efficiency of PC leading to markedly elevated anticoagulant activity. 3 Once activated, APC may either dissociate from EPCR or remain bound to EPCR and display an anti-inflammatory, antiapoptotic, and cytoprotective role. 4 The human EPCR gene is located on chromosome site 20 (q11.2), spans 8 kb, and comprises 4 exons. Exon 1 encodes the 5′-untranslated region (UTR) and the signal peptide; exons 2 and 3 encode most of the extracellular region; and exon 4, the remaining parts of the protein and the 3′-UTR. 5
A soluble form of EPCR (sEPCR), which lacks the transmembrane domain and cytoplasmic tail, circulates in plasma and is generated through proteolytic cleavage of membrane-bound EPCR by metalloprotease activity induced by thrombin. It binds PC and APC with similar affinity and inhibits PC activation on endothelium. 6 It also inhibits APC anticoagulant activity by blocking its binding to phospholipids and by abrogating its ability to inactivate factor Va. 7 Increased sEPCR levels may be associated with increased risk of thrombosis, as sEPCR can inhibit APC activity, as well as PC activation, by competing for PC with membrane-associated EPCR.
Venous thromboembolism (VTE) is a common multifactorial disorder, affecting 1 to 3 per 1000 people each year in the Western world. Both genetic and acquired factors are involved in the pathogenesis of the disease. The most common genetic risk factors for VTE in caucasians are associated with uncompromised control of thrombin formation. Factor V (FV) Leiden is caused by a 1691 G-A transition in the procoagulant FV gene and is associated with resistance to APC. 8 It is present in 3% to 8% of caucasians and in 15% to 50% of patients with VTE. 9 Clinical manifestations associated with FV Leiden exhibit a wide intra- and interfamilial variation, probably due to concomitant presence of FV Leiden with other risk factors. 10 The prothrombin factor II (FII) G20210A mutation is caused by a 3′UT G-A transition in the procoagulant FII gene and is associated with increased plasma prothrombin levels. The prevalence of the FII 20210A allele in the caucasian population is 2% to 3%, and its carriership is associated with a slight increase in the risk of VTE. Presence of other genetic or acquired risk factors further increases the risk of VTE in FII 20210A carriers. 11 Deficiency in the natural inhibitors of the coagulation pathway (PC, PS, and antithrombin [AT]) is also associated with increased risk of thrombosis but is very rare in caucasians. 7
Polymorphisms in other anticoagulant pathways may also modify the risk of thrombosis. 12,13 The involvement of EPCR in the anticoagulant pathway of hemostasis suggests a potential role in the pathogenesis of thrombosis. 14,15 Blocking of EPCR seems to accelerate thrombus development in vivo. 16 Two common polymorphisms in EPCR (4600A/G [rs867186] in exon 4, which predicts a substitution of Ser219 by Gly in the transmembrane region of EPCR and 4678G/C [rs9574] in the 3′-UT region) have been studied regarding their involvement in the development of thrombosis. There are several data in the literature indicating that the 4678C allele is associated with a protective advantage for thrombosis, increased plasma-activated PC levels, and reduced sEPCR levels, while the 4600G allele is associated with an increased risk of thrombosis and increased plasma sEPCR levels, 17 although these conclusions are questioned by others. 18,19
We investigated the implication of these 2 common polymorphisms of EPCR in the development of thrombosis by evaluating the relative contribution of the above-mentioned EPCR polymorphisms along with sEPCR levels in patients with thrombosis and hereditary thrombophilia or without known genetic predisposition. Furthermore, we studied the association of these 2 polymorphisms with the age of the first thrombotic event and the recurrence of thrombotic episodes.
Material and Methods
The study was performed in the Hematology Laboratory-Transfusion Unit of the General University Hospital of Athens “ATTIKON” from September 2009 until May 2012. We recruited 84 patients aged 22 to 83, as well as a control group of 100 healthy individuals matched for age and sex. All individuals were caucasians and completed a detailed questionnaire including data of personal and family history of thrombosis, malignancy, and drug intake. The study was approved by the local ethics committee, and all the patients gave their informed consent. Patients were studied for hereditary thrombophilia (FV Leiden and FII G20210A and deficiency of natural inhibitors of coagulation [PC, PS, and AT]). Automated hemostasis analyzer BCS System (Siemens GmbH, Germany) was used to determine the activity of natural inhibitors of coagulation by chromogenic and immunologic assays in plasma samples stored at −60°C. The sEPCR was measured with enzyme linked immunosorbent assay (Asserachrom-sEPCR; Stago Diagnostica, France).
Detection of FV Leiden and FII G20210A was performed in genomic DNA samples extracted from peripheral venous blood sample, anticoagulated with EDTA, and stored at 4°C, using standard procedures (Qiagen Inc, Valencia, California). The FV Leiden and FII G20210A mutations were assessed by polymerase chain reaction (PCR) and reverse hybridization that concurrently detects these 2 mutations (FV-PTH SripAssay; ViennaLab, Austria). The different target gene sequences were concurrently amplified and biotin-labeled in a single amplification reaction. The reaction consisted of 5 μL of DNA added to 15 μL already prepared PCR amplification mix, including primers and deoxynucleotide triphosphates in the presence of 1 U Taq polymerase. The amplification program consisted of an initial step at 94°C for 2 minutes, followed by 30 cycles at 94°C for 15 seconds, 58°C for 30 seconds, 72°C for 30 seconds, and a final extension step at 72°C for 3 minutes. The amplification products were selectively hybridized to a test strip that contained allele-specific oligonucleotide probes immobilized as an array of parallel lines. Bound biotinylated sequences are detected using streptavidine alkaline phosphatase and color substrates.
Real-time PCR (RT-PCR) was used for the identification of the 4600A/G and 4678G/C polymorphisms in EPCR gene (LightCycler 2.0 Instrument; Roche Diagnostics GmbH, Germany). Primers and probes were designed by TIB Molbiol, Berlin, Germany. The RT-PCR mixture was prepared in a total volume of 20 μL. The reaction mixture for the 4600A/G polymorphism of EPCR was optimized as follows: 2 μL DNA, 10× Fast start reaction mix, 3 mmol/L MgCl2, 0.5 μmol/L of forward primer S (5′-TCgggCTAACTCTTTgCATgTTC-3′), and 0.25 of reverse primer A (5′-ggggAgTCTggCTTCACAgTg-3′). The reaction mixture for the EPCR 4678G/C polymorphism consisted of 2 μL DNA, 10× Fast start reaction mix, 2 mmol/L MgCl2, 0.25 μmol/L of forward primer F (5′-CgCTTTggggTTTgACTCAAAT-3′), and 0.5 μmol/L of reverse primer (5′-CAgCCTCCATCAATCCAgTCC-3′). LightCycler capillaries were placed in a precooled centrifuge adaptor, in which DNA was loaded followed by 18 mL of the master mix and a quick centrifugation. The DNA was amplified by applying 10 minutes of hot start at 95°C, followed by 45 cycles of denaturation at 95°C for 5 seconds, annealing at 60°C for 10 seconds, and elongation at 72°C for 15 seconds. Subsequently, a melting curve program was applied with a temperature increment from 40°C to 85°C at 0.2°C/s. The fluorescence reading was continuously taken by the LightCycler at each temperature increment. Finally, the capillaries were cooled to 4°C. After completion of all programs, melting peak graphs were generated from the LightCycler Software 2.0.
Statistical analysis was performed using SPSS package, version 2.0 (SPSS software). Fischer exact test was used in order to detect differences in allele frequencies between groups (significance level: P = .05). t Test was used in order to compare differences in sEPCR levels.
Results
Eighty four patients (38 males and 46 females) aged 22 to 83 (mean age 47) were included in the study. The control group consisted of 100 healthy controls (40 males and 60 females) aged 45 to 87 (mean age 49), without personal or family history of thrombosis. In all, 38 (45.2%) patients were younger than 35 years and 46 (54.8%) patients were older than 35 years, when they experienced their first thrombotic episode. The mean age of the first thrombotic episode was 45 years. In all, 46 (54.8%) reported a family history of thrombosis and 40 (42.1%) were smokers. In all, 42 (50%) had experienced a unique episode of thrombosis, 24 (28.6%) patients reported 2 episodes, and 18 (21.4%) patients 3 or more episodes. All 84 patients had experienced at least 1 thrombotic episode such as VTE (58, 69%), stroke (15, 17.9%), and myocardial infarction (11, 13.1%). Demographic characteristics and clinical data of the study patients are shown in Table 1.
Demographic and Clinical Data of the Study Patients.
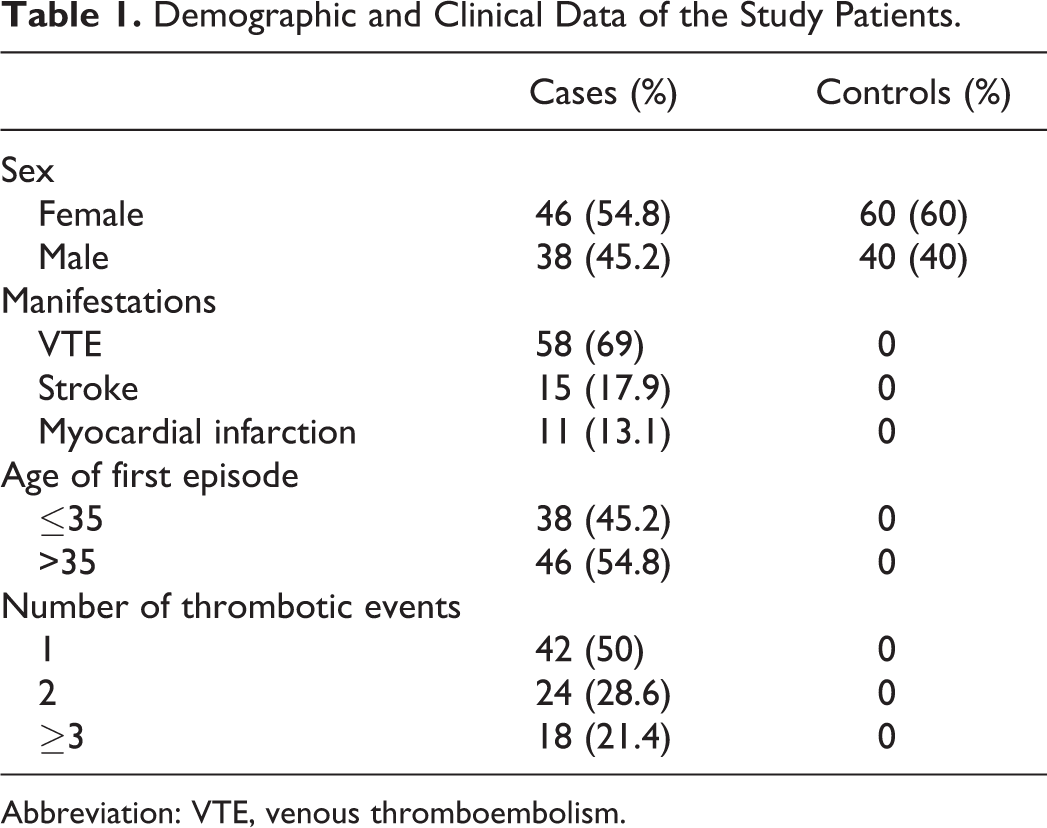
Abbreviation: VTE, venous thromboembolism.
No patients were detected with deficiency in the natural inhibitors (PC, PS, and AT). The FV Leiden was detected in 13 (15%) patients versus 8 (8%) controls (P = .05), while FII G20210A was present in 7 (8%) patients versus 3 (3%) controls (P = .02). Soluble EPCR levels were evaluated in patients (N = 58) who were not on oral anticoagulants (oral anticoagulants markedly decrease sEPCR levels). 20 Soluble EPCR levels in patients and controls according to their genetic background are depicted in Table 2.
Soluble EPCR Levels of 4600A/G and 4678G/C Polymorphisms in Patients and Controls.
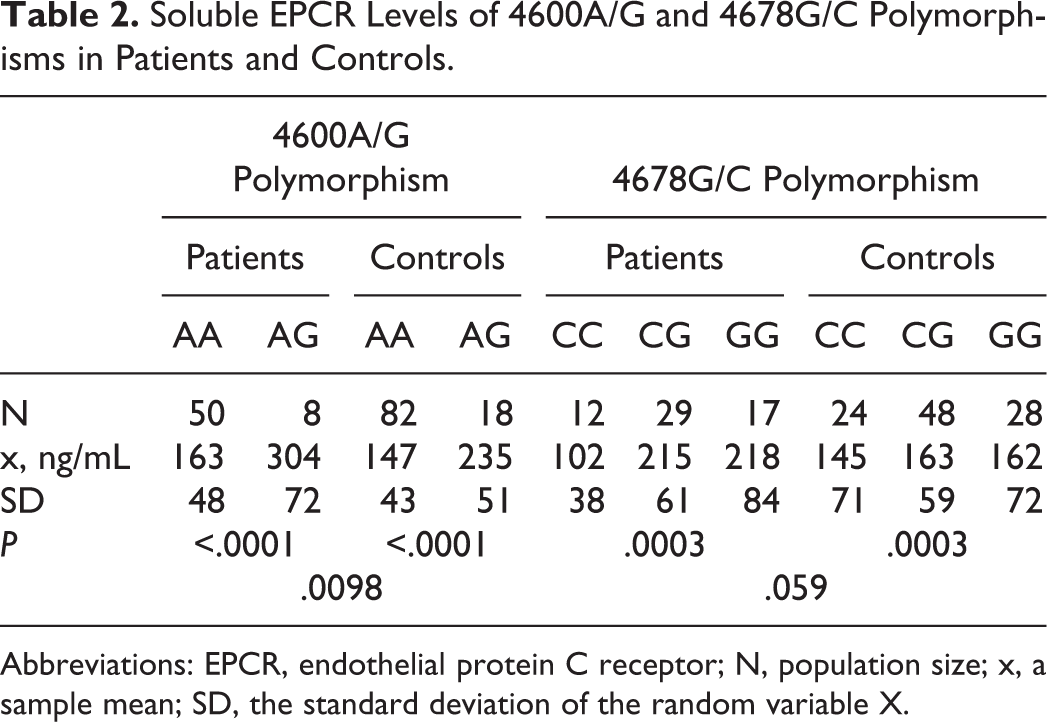
Abbreviations: EPCR, endothelial protein C receptor; N, population size; x, a sample mean; SD, the standard deviation of the random variable X.
Soluble EPCR was statistically elevated in carriers of the 4600G allele compared to noncarriers in both control (235 ± 51 vs 163 ± 43 ng/mL, P < .0001) and patient (304 ± 72 vs 147 ± 48 ng/mL, P < .0001) groups. Additionally, statistically significantly higher levels of sEPCR were detected in patient compared to control carriers of the 4600G allele (304 ± 72 vs 235 ± 51 ng/mL, P = .001). Concerning 4678CG polymorphism, 4678CC carriers presented with lower sEPCR levels when compared to 4678GG carriers in both control and patient groups (P = .0003). However, there was only a trend that 4678CC controls had lower sEPCR levels compared to patients (P = .06).
Regarding EPCR polymorphisms, 13 (15.5%) patients carried the EPCR 4600A/G genotype compared to 18 (18%) controls, while 71 (84.5%) patients carried the EPCR 4600A/A genotype (P = .7NS) compared to 82 (82%) controls. The allele frequency of the EPCR 4600G allele was 0.08 in patients versus 0.09 in controls (P = .7NS). No EPCR 4600G/G homozygous patient was found.
In all, 20 (23.8%) patients carried the EPCR 4678C/C genotype compared to 24 (24%) controls and 40 (47.6%) patients carried the EPCR 4678G/C genotype compared to 48 (48%) controls, while 24 (28.6%) carried the EPCR 4678G/G genotype compared to 28 (28%) controls. The allele frequency of the EPCR 4678C allele was 0.47 in patients versus 0.48 in controls. The prevalence of EPCR genotypes in patients and controls is depicted in Table 3.
Prevalence of EPCR 4678G/C and EPCR 4600A/G Polymorphisms in Patients and Controls.
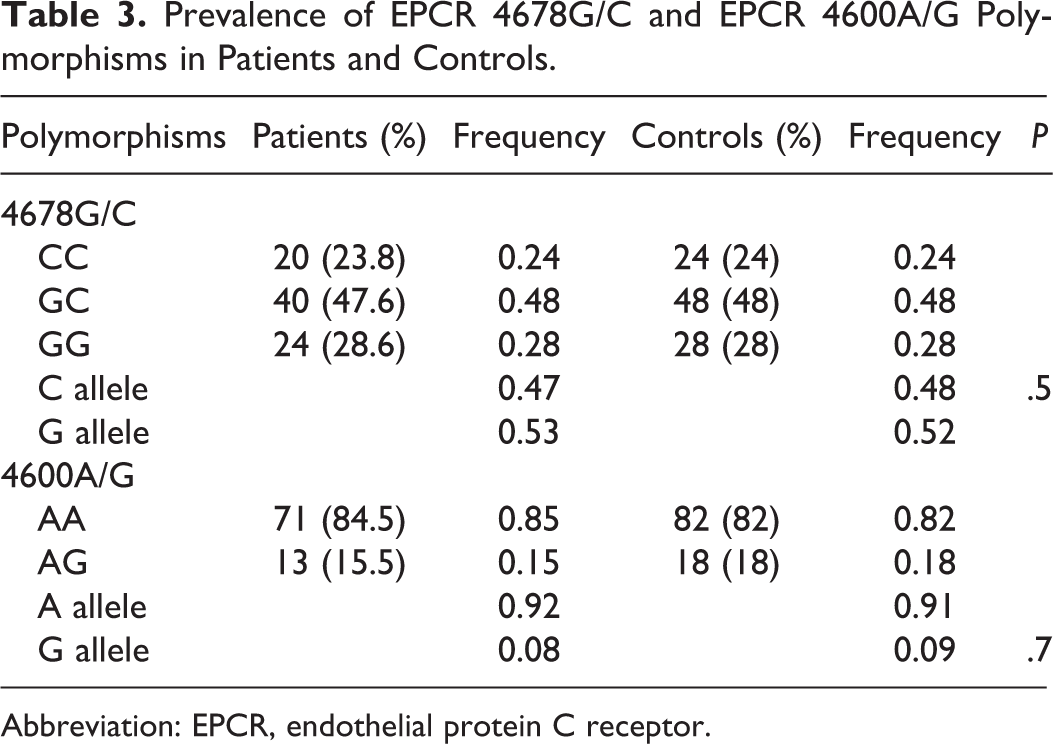
Abbreviation: EPCR, endothelial protein C receptor.
The distribution of EPCR 4600A/G and 4678G/C polymorphisms among the age-groups of patients is presented in Table 4. The EPCR 4600G allele was more prevalent (0.11) among patients who developed thrombosis at younger age (<35) compared to older (>35) patients (0.04), P = .08.
EPCR 4600A/G and EPCR 4678G/C in Relation to Age of First Thrombotic Event.
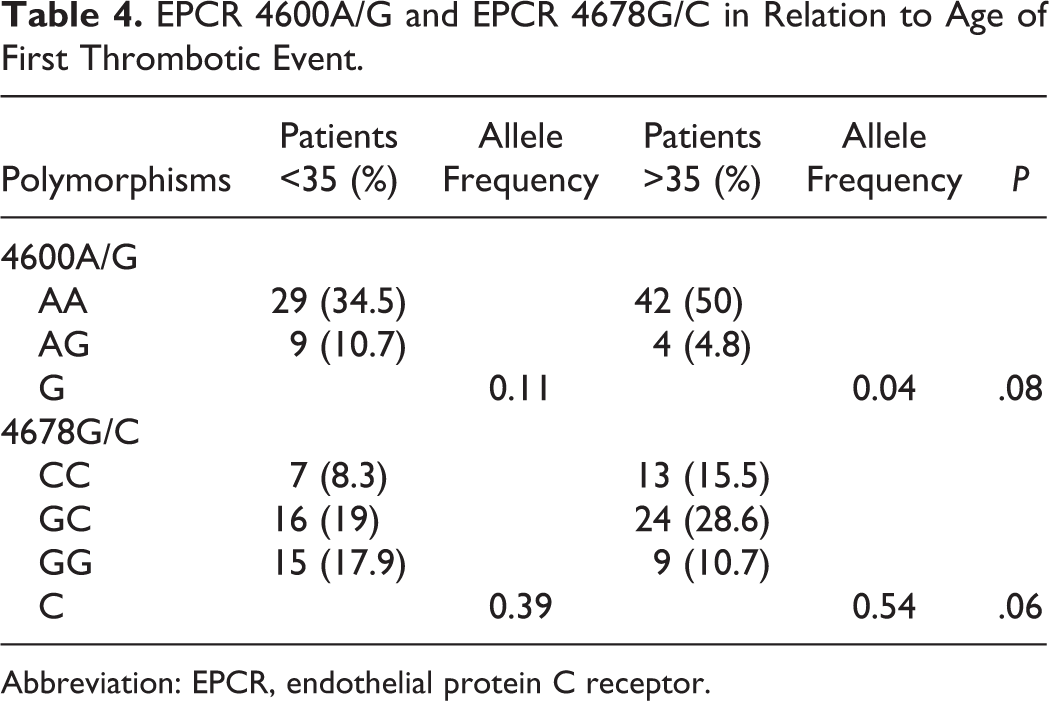
Abbreviation: EPCR, endothelial protein C receptor.
The EPCR 4678C allele was less prevalent in the <35 age-group compared to the >35 group (0.39 vs 0.54, P = .06). In the younger age-group (<35), EPCR 4678C/C genotype was less frequent (7 of 38 patients, 18.4%) compared to EPCR 4678G/C (16 of 38 patients, 42.1%) and EPCR 4678G/G (15/38 patients, 39.5%; P = .04 and P = .07, respectively). In the >35 age-group, the EPCR 4678C/C was also less prevalent (13 of 46 patients, 28.3%) compared to EPCR 4678G/C (24 of 46 patients, 52.2%; P = .03) and EPCR 4678G/G patients (9 of 46, 19.5%; P = .46).
Among the 13 EPCR 4600A/G patients, only 1 carried the EPCR 4678C/C genotype (female, 33 years old, superficial vein thrombosis, 3 events). Additionally, among EPCR4600AG patients who presented with early thrombosis (<35 years), no one carried the FV Leiden mutation and only 1 was a FII G20210A carrier (who had experienced upper limb thrombosis at the age of 19).
Table 5 depicts the distribution of EPCR 4600A/G and 4678G/C genotypes among patients in relation to number of thrombotic events. Among EPCR 4600A/G patients (N = 13), 8 (61.5%) experienced a unique thrombotic episode compared to 3 (23.1%) patients with 2 episodes and 2 (15.4%) who reported 3 or more events (P = .11, P = .04, respectively). The prevalence of EPCR 4678C/C was 50% (10 of 20) in patients with a unique episode compared to 30% (6 of 20) and 20% (4 of 29) in patients with 2 and more than 3 thrombotic episodes (P = .3 and .09, respectively). When comparing patients with a unique episode to those with relapses, we also observed comparable distribution of the genotypes in all subgroups (10 of 20 vs 10 of 20 in the 4678C/C, 20 of 40 vs 20 of 40 in the 4678C/G, and 12 of 24 vs 12 of 24 in the 4678G/G subgroups).
Frequency of EPCR 4600A/G and EPCR 4678G/C Polymorphisms in Relation to Relapses.
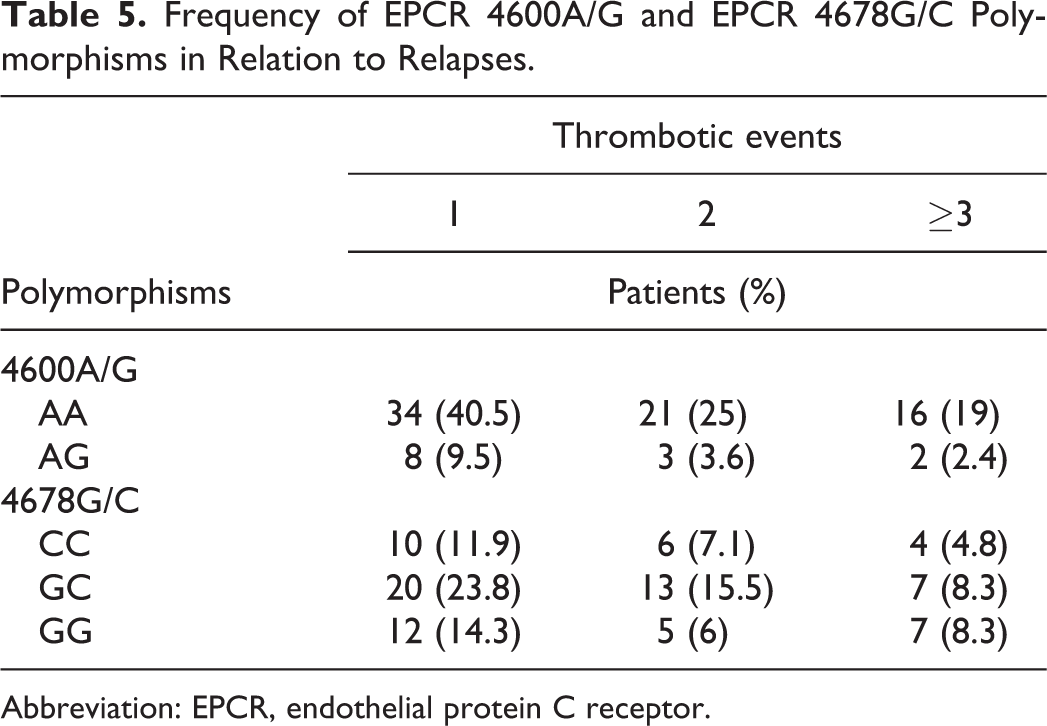
Abbreviation: EPCR, endothelial protein C receptor.
Discussion
There is convincing evidence that the risk of first thrombotic event and recurrence depends on the interaction between multiple congenital and exogenous interactions. 21 As a result, the risk may vary widely both within and among individuals. We studied a group of patients with a range of thrombotic manifestations in order to evaluate the implication of “classic” hereditary background (FV Leiden, FII G20210A, and deficiency of AT, PC, and PS) as well as the additional involvement of polymorphisms of the EPCR gene in the development and recurrence of thrombotic events.
Functional polymorphisms in the EPCR gene have been shown to modulate the risk of thrombosis, especially in carriers of prothrombotic mutations. 20 Four haplotypes of EPCR (H1, H2, H3, and H4), 3 of which contain 1 or more single-nucleotide polymorphisms (SNPs), specific for each haplotype have been described so far. 22 The 4600AG SNP predicts a conformational change (Ser 219-Gly) that renders EPCR more susceptible to metalloprotease cleavage and thus leads to impaired PC–EPCR interaction. The sEPCR is connected to the PC and APC with the same affinity as transmembrane EPCR. Thus, it can compete and prevent activation of PC to APC but also prevents the anticoagulant pathway of PC. Generation of sEPCR is a result of cleavage of EPCR in the transmembrane encoded by the exon 4 near the 4600 position. Additionally, it was recently reported that 4600AG is associated with reduced membrane bound EPCR and increased sEPCR in human umbilical vein endothelial cell-conditioned medium, higher levels of a truncated messenger RNA isoform, and lower rate of PC activation. 18
In our study, the frequency of the 4600G allele was comparable in patients and controls (P = .7), suggesting that the presence of the G allele per se does not increase the risk of thrombosis. These results agree with those previously reported 4,23 but are in disagreement with those by Saposnik et al 19 who found that 4600G allele was overrepresented in patients with thrombosis. However, it is worthwhile noticing that in an analysis of a subset of patients in terms of age, we observed that the EPCR 4600G allele is more prevalent among patients who developed thrombosis at younger age (<35) compared to older age (>35) patients and that its presence is associated with increased sEPCR levels.
Soluble EPCR levels in patients and controls were comparable to those described previously. 12,13,16 Carriers of the 4600G allele presented with statistically higher levels of sEPCR in both control and patient groups compared to carriers of the 4600A allele. Moreover, higher levels of sEPCR were detected in patient carriers of the 4600G allele compared to control carriers of 4600G allele.
In terms of the EPCR 4678C/G polymorphism, the H1 haplotype (4678C allele) is considered to be associated with a lower risk of thrombosis, irrespectively of hereditary thrombophilic status of the patients. 24 It has been observed that 4678C/C homozygous patients have higher levels of APC than carriers of 1 4678C allele, which suggests that H1 haplotype is protective probably because of high levels of APC (gain of function mutation). 18 Carriers of both FV Leiden mutation and EPCR 4678C/C genotype (H1/H1) present thrombosis later than carriers of FV Leiden mutation who don’t carry the EPCR 4678C/C genotype. Furthermore, these patients have greater likelihood not to experience thrombosis until the age of 40. The mechanism by which it reduces the risk of VTE may be similar to that suggested in venous thrombosis: increased circulating APC identified in carriers of the C allele may protect the organism from vascular damage and prolong not only endothelial, but also cellular and organ survival. 13
In our study, the prevalence of the 4678C EPCR allele was comparable in patients and controls. Our results agree with those reported by Galligan et al. 25 However, the prevalence of EPCR 4678C allele was significantly lower in younger patients compared to older patients. Although this difference did not reach statistical significance, there is a clear trend suggesting that the lack of the protective effect of EPCR 4678C/C may lead to earlier development of thrombotic manifestations. Additionally, among 24 patients who lacked the 4678C allele (4678G/G genotype), 15 experienced their first episode before the age of 35 compared to 9 who developed thrombosis later (P = .14). These data suggest that the presence of EPCR 4678C allele might contribute to the delay of the development of a thrombotic event. As far as sEPCR is concerned, 4678CC patients presented with lower levels compared to other genotypes. In our study, neither the EPCR 4600A/G nor the 4678G/C polymorphisms seem to be associated with relapses in any patient subgroup.
Coexistence of FV Leiden with either other rare congenital deficiencies (PC, PS, and AT) or acquired risk factors is already known to increase the risk of thrombosis. 26,27 A Spanish study shows that the risk of thrombosis in carriers of FII G20210A mutation is influenced by 4600G allele, through increased plasmatic levels of sEPCR. 12 In our study, 2 patients carried FV Leiden and EPCR 4600A/G simultaneously. They both presented with a first episode of PE; a male patient presented with a unique episode at the age of 36 and a female presented with PE at the age of 50 followed by 2 DVT episodes. Coexistence of FIIG20210A and EPCR 4600AG was detected only in 1 patient with a first episode of superficial vein thrombosis at the age of 19 followed by 2 relapses.
The FV Leiden and FII G20210A mutations were statistically more frequent in our patient group than in controls as expected (P = .05, P = .02, respectively). Additionally, we observed that only 3 (7.5%) FV Leiden heterozygotes versus 37 (92.5%) wild-type patients and 2 (5%) FII G20210A heterozygotes versus 40 wild type (95%) have experienced relapse. This finding further confirms previous reports in the literature, 28 suggesting that these common risk factors are not associated with higher risk of relapse. The absence of patients with deficiency in natural inhibitors of coagulation (PC, PS, and AT) further confirms the hypothesis that contribution of these congenital risk factors in the development of thrombosis cannot be evaluated in small patient groups due to the fact that the incidence of these deficiencies is very rare in caucasian populations. 9
A unique feature of our study is that 45.2% (38 of 84) of patients enrolled experienced the first thrombotic episode at a very young age <35. Studying a group that develops thrombosis very early can be a valuable tool for highlighting interaction between genetic factors since young patients have not yet been exposed to environmental and age-dependent stimuli. The insights gained from studies that evaluate the involvement of EPCR polymorphisms in the risk of thrombosis may not only elucidate underlying pathophysiological mechanisms but also their clinical significance and the interplay between these polymorphisms and other congenital and/or epigenetic risk factors. It has been previously suggested that sEPCR levels may be used as a marker of vascular disease. 29 The limitations set by the small number of patients in our study would not allow us to draw firm conclusions but only to suggest possible interactions. Further studies including larger number of patients as well as the evaluation of markers such as endogenous thrombin potential and sEPCR along with EPCR polymorphisms, not only at the onset of thrombosis but also during relapses and symptom-free periods, might further elucidate the pathogenesis of thrombotic events.
Conclusion
We retrospectively studied 84 patients with thrombotic events, 45.2% of whom were younger than 35 years at the first manifestation. We found that although the prevalence of 4600G and 4678C alleles was comparable between patients and controls, carriers of the 4600G were more prone to develop a thrombotic event earlier in life, while carriers of the 4678C alleles seemed protected against early development of thrombosis. 4600G allele was associated with higher sEPCR levels compared to 4600A allele in both controls and patients. Additionally, sEPCR levels were lower in patients with 4678C/C genotype compared to carriers of other genotypes.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.