
Abstract
Cannabinoids can mimic the infarct-reducing effect of early ischemic preconditioning, delayed ischemic preconditioning, and ischemic postconditioning against myocardial ischemia/reperfusion. They do this primarily through both CB1 and CB2 receptors. Cannabinoids are also involved in remote preconditioning of the heart. The cannabinoid receptor ligands also exhibit an antiapoptotic effect during ischemia/reperfusion of the heart. The acute cardioprotective effect of cannabinoids is mediated by activation of protein kinase C, extracellular signal-regulated kinase, and p38 kinase. The delayed cardioprotective effect of cannabinoid anandamide is mediated via stimulation of phosphatidylinositol-3-kinase-Akt signaling pathway and enhancement of heat shock protein 72 expression. The delayed cardioprotective effect of another cannabinoid, Δ9-tetrahydrocannabinol, is associated with augmentation of nitric oxide (NO) synthase expression, but data on the involvement of NO synthase in the acute cardioprotective effect of cannabinoids are contradictory. The adenosine triphosphate-sensitive K+ channel is involved in the synthetic cannabinoid HU-210-induced cardiac resistance to ischemia/reperfusion injury. Cannabinoids inhibit Na+/Ca2+ exchange via peripheral cannabinoid receptor (CB2) activation that may also be related to the antiapoptotic and cardioprotective effects of cannabinoids. The cannabinoid receptor agonists should be considered as prospective group of compounds for creation of drugs that are able to protect the heart against ischemia–reperfusion injury in the clinical setting.
A Brief History of the Discovery of Endogenous Cannabinoid System
The endogenous cannabinoid system has been discovered relatively recently. The central cannabinoid receptor (CB1) was discovered in 1990, and its structure was revealed when it was cloned. 1 The CB1 is coupled to Gi/o-protein and is widespread in the brain. 1,2 The CB1 was found in the myocardium of adult rats 3,4 and mice. 5 In 1993, a peripheral cannabinoid receptor (CB2) was identified and cloned. 6 The CB2 receptor is also coupled to Gi/o-protein and is most abundant in spleen. 7 –9 This receptor was also found in neonatal rat cardiomyocytes 10 and in the left ventricle of adult mice. 5 Activation of both CB1 and CB2 receptors promotes inhibition of adenylyl cyclase and voltage-gated Ca2+ channels. They also activate mitogen-activated protein kinases (MAPK) and phospholipase C (PLC). 2,9,11 Activation of both MAPK and PLC is associated with cardioprotection.
In 1999, an orphan G-protein-coupled receptor consisting of 319 amino acids was found in some human brain nuclei. 12 This receptor was named “GRP55.” It was later established that GRP55 is also a cannabinoid receptor and is expressed in the brain stem, frontal cortex, striatum, hypothalamus, cerebellum, and hippocampus. 13 GPR55 was later found in peripheral organs such as adrenals, spleen, jejunum, and ileum. Only trace quantity of GPR55 messenger RNA (mRNA) was found in other organs, and none has been found in the heart. 13 The GPR55 receptor is distinguished from other endogenous cannabinoid receptors by its differential affinity to CB ligands. It exhibits high affinity to CP55,940, anandamide, Δ9-tetrahydrocannabinol (Δ9-THC), palmitoylethanolamine, and HU-210, but it has low affinity to the CB1 antagonist SR141716 and does not interact with the CB1/CB2 agonist WIN55,212-2. 13 The GPR55 receptor couples to Gα13-protein but does not interact with Gi- and Gq-protein. 13 Selective antagonists of GPR55 have not yet been synthesized, so the regulatory function (if any) of GPR55 remains obscure.
In 2003, Fride et al 14 using CB1 receptor knockout mice obtained indirect evidence of the existence of a third (CB3) receptor. It is unique in that this receptor does not interact with the CB1/CB2 agonists WIN55,212-2 or CP55,940. 14 The Kunos laboratory 15,16 had also proposed the existence of another cannabinoid receptor in 1999. The receptor was present in the vascular endothelium and exhibited high affinity to the endogenous cannabinoid anandamide and methanandamide (synthetic analogue of anandamide) but did not interact with other CB receptor agonists. 16,17 This receptor can be inhibited by the CB1 receptor antagonist SR141716A but not by the CB2 receptor antagonist SR144528. 16,17 This receptor was named “SR141716A-sensitive anandamide receptor” to distinguish it from CB1 receptors. 16 In 2000, Di Marzo et al 18 discovered yet another G-protein-coupled anandamide receptor in the brain of CB1 knockout homozygote mice. This receptor was insensitive to SR141716A and SR144528 and apparently was different from the SR141716A-sensitive anandamide receptor found by Kunos’s group.
Kunos’s group discovered yet another Gi/o-protein-coupled endothelial receptor in 2003. This receptor exhibited a high affinity to the regioisomer of cannabidiol, abnormal cannabidiol. 19,20 This receptor promoted endothelium-dependent vasodilation of isolated rat mesenteric arteries via opening of Ca2+-dependent K+ channel. 19,20 This effect was abolished by the cannabidiol analog O-1918 but not by CB1 and CB2 antagonists. 19,20 Experiments on human umbilical vein endothelial cells showed that increase in outward K+ current, produced by abnormal cannabidiol, was blocked by KT-5823, an inhibitor of protein kinase G, or 1H-[1,2,4]oxadiazolo[4,3-a]quinoxaline-1-one, an inhibitor of soluble guanylate cyclase. 19 It has also been established that in endothelial cells, abnormal cannabidiol activates extracellular signal-regulated kinase (ERK) and Akt kinase. 20 Abnormal cannabidiol-induced vasorelaxation was maintained after inhibition of nitric oxide (NO) synthase by N-nitro-L-arginine methyl ester (L-NAME) (N-nitro-L-arginine methyl ester) or in combination with vanilloid VR1 receptor antagonist capsazepine. 20 The authors concluded that a Ca2+-activated K+ current in endothelial cells is potentiated by activation of a Gi/o-coupled receptor distinct from CB1 or CB2. This receptor signals through cyclic guanosine monophosphate (cGMP) and protein kinase G to increase the K+ channel availability or its sensitivity to voltage and/or Ca2+. 19
Thus, to date, the existence of at least 5 cannabinoid receptors has been confirmed: CB1, CB2, GPR55, anandamide receptor (there may actually be 2 distinct anandamide receptors), and cannabidiol receptor. It is likely that more cannabinoid receptors are yet to be discovered.
Devane et al discovered the first endogenous cannabinoid N-arachidonylethanolamine (anandamide). 21 We (Mechoulam group) subsequently found that 2-arachidonyl glycerol (2-AG) is also an endogenous cannabinoid, 22 and in 2001, we found endocannabinoid 2-arachidonyl glyceryl ether (noladin ether). 23 Another endocannabinoid, oleamide, was identified in 2004. 24 This was followed by a discovery of N-arachidonoyl-L-serine 25 and RVD-Hemopressin. 26 A list of the currently available cannabinoid ligands and their properties are presented in Table 1.
A List of Cannabinoid Ligands and its Properties.

Effect of CB1 and CB2 Receptors on Cardiac Tolerance to Ischemia–Reperfusion Injury
The ability of cannabinoids to increase the heart’s tolerance to the pathogenic impact of ischemia/reperfusion has now been demonstrated in a number of studies 27 –34 and are summarized in Table 2 of our review. The infarct-sparing effect of these compounds has been shown both in vivo 5,27,28,34,39 and ex vivo. 29 –33,39 These investigators suggest that the heart represents a direct target of exogenous cannabinoids in the implementation of their cardioprotective effects. 29 –33,39 We (Maslov group) found that treatment with CB1/CB2 receptor agonist HU-210 prior to ischemia promotes a decrease in the fraction of the ischemic zone that is infarcted (IS/AAR ratio) in vivo. 28,34 We have found that infarct-sparing effect of HU-210 (0.1 mg/kg) in vivo disappeared after blocking CB1 receptor with SR141716A but was maintained after CB2 receptor blockade with SR144528 29 or neural ganglion inhibition by hexamethonium. 41 Addition of HU-210 to the perfusion solution prior to 45 min global ischemia and 30 min reperfusion of isolated rat heart promoted a decrease in the amount of creatine kinase (CK) leaked into the coronary effluent. 29 The selective CB1 agonist arachidonylcyclopropylamide (ACPA) exhibited a similar effect in vitro, while the selective CB2 agonist JWH 133, anandamide, and methanandamide had no effect on cardiac tolerance to ischemia–reperfusion injury in vitro. 37,41 However, our recent in vivo experiments showed that JWH133 at high dose (20 mg/kg intravenously) was able to reduce the IS/AAR ratio. 42 Pretreatment with the CB2 receptor antagonist AM630 abolished infarct-reducing effect of JWH133. These results indicate the involvement of CB2 receptors as well in the infarct-limiting effect of JWH133. Thus, cardioprotective effect of cannabinoid receptors is well established, but the data on the role of particular cannabinoid receptors are contradictory and need to be further investigated.
Cannabinoid Receptor Agonists Reported to Protect the Heart Against Ischemia–Reperfusion Injury.a

Abbreviations: KC, protein kinase C; PI3K, phosphatidylinositol-3-kinase; Akt, kinase isolated from AKR thymoma cell line; p38 MAPK, p38 mitogen activated kinase; MEK1/2, mitogen-activated protein kinase kinase; ERK1/2, extracellular signal-regulated kinase; PKC, protein kinase C; NO, nitric oxide; CB, cannabinoid; KATP, ATP-sensitive K+ channel. aExperiment conditions: in vivo, experiments on isolated cells; ex vivo, experiments on isolated heart; in vivo, experiments on a whole animal. “No data” means that the data on this particular subject either absent or controversial.
Can Cannabinoids Postcondition the Heart?
A major clinical indication for a cardioprotectant is acute myocardial infarction when a coronary artery is suddenly blocked with a thrombus. Emergency recanalization of the artery is the treatment of choice but, unfortunately, this can seldom be accomplished before a significant fraction of the ischemic myocardium is irreversibly injured. A series of very brief reperfusion/occlusion cycles at the onset of reperfusion (termed ischemic postconditioning) offer protection to the heart similar to that with preconditioning. 43 Ischemic postconditioning in the clinical setting is difficult however, and there is a need for a pharmacological substitute. Two studies related to cannabinoids have addressed this. Defer et al 27 reported that a 5-minute injection of the selective CB2 agonist JWH133 (3 mg/kg) at the end of 1-hour coronary artery occlusion decreased the IS/AAR ratio in wild-type mice. This protection was not observed in CB2 receptor knockout mice. 27 In our experiments, 5-minute intravenous administration of the CB1/CB2 receptor agonist HU-210 (0.1 mg/kg) before reperfusion in isolated rat hearts reduced the IS/AAR ratio by half. 35 The above-mentioned results imply that stimulation of CB2 receptors by exogenous cannabinoids do promote an anti-infarct effect when administered just prior to reperfusion both in vivo and ex vivo. Cleary, more postconditioning studies are needed.
Cannabinoids and the No-Reflow Phenomenon
Another consequence of ischemia/reperfusion is decreased coronary reflow in the reperfused myocardium (the no-reflow phenomenon). Ischemic preconditioning improves reflow and the endogenous cannabinoids palmitoylethanolamide (PEA) and 2-AG mimicked the vasoprotective effect. 44 Gonzalez et al published novel results of experiments carried out on isolated perfused rat heart. 38 The investigators administered the CB1 and CB2 receptor agonist WIN 55,212-2 (1 mg/kg) intraperitoneally and then removed the hearts and performed global ischemia and reperfusion of the hearts ex vivo. They found that WIN 55,212-2 had no effect on the left ventricular developed pressure and end-diastolic pressure but significantly attenuated the rise in coronary resistance during reperfusion. The reduction in coronary resistance was interpreted by the authors as a vasoprotective effect. 38 The vasoprotective effect was unaffected by the blockade of CB1 receptors but disappeared after inhibition of CB2 receptors. 38 They concluded that exogenous cannabinoids prevent injury to the endothelium of coronary arteries evoked by ischemia/reperfusion via activation of CB2 receptors.
Cannabinoids and Remote Ischemic Preconditioning
Gho and colleagues found that a preliminary 15-minute occlusion of mesenteric arteries increases cardiac tolerance to ischemia/reperfusion with a reduced IS/AAR ratio. 45 Fifteen-minute ischemia of the left kidney also decreased the IS/AAR following coronary artery occlusion/reperfusion. 45 This cardioprotective phenomenon was named “remote ischemic preconditioning.” 45 –47 Interestingly, CB2 (but not CB1) receptor inhibition, eliminated the cardioprotective and antiarrhythmic effect of remote preconditioning with intestine ischemia in rats. 48 Because the authors did not determine the level of cannabinoids in the blood, it was not clear whether endogenous cannabinoids were the circulating cardioprotective factor or whether they only acted as intermediaries between cardiomyocytes and an unidentified circulating factor. Nevertheless, these data indicate that endogenous cannabinoids are involved at some level in the infarct-limiting effect of remote ischemic preconditioning. No data are yet available whether cannabinoids participate in remote preconditioning with limb ischemia.
Controversy as to Which Cannabinoid Receptors Protect the Heart
In experiments with isolated rat hearts, Underdown et al 40 have found that anandamide (1 μM) and methanandamide (1 μM) decrease the IS/AAR ratio after 30-minute global ischemia and 1-hour reperfusion by 2.5-fold. Because, the CB1/CB2 receptor agonist HU-210 (1 μM) did not exert a similar effect, they concluded that neither CB1 nor CB2 was the target. Accordingly, the authors did not observe an infarct-reducing effect of the selective CB1 receptor agonist ACPA (1 μM) or the selective CB2 receptor agonist JWH-133 (1 μM). 40 The beneficial effect of anandamide was also not affected by the blockade of CB1 or CB2 receptors. The authors concluded that the cardioprotective effect of anandamide is dependent upon a cannabinoid receptor different from CB1 or CB2. 40
The above-mentioned work contradicts to our data, where we (Maslov group) were able to show cardioprotective effect of HU-210 at the concentration of 0.1 μM and ACPA at the concentration of 1.0 μM. 29,33,37,41 We suggest that this discrepancy is due to the differences in the experimental protocol. Underdown and colleagues administered HU-210 and ACPA from 5 minutes before ischemia until the end of reperfusion. 40 In contrast, we mimicked the phenomenon of preconditioning by perfusion of the hearts with HU-210 and ACPA for 10 minutes followed by 10 minutes washout prior to ischemia. 29,33,41 Furthermore, Underdown et al. used a rather high concentration of HU-210 (1 μM). 40 According to our data, perfusion of hearts with HU-210 at the concentration of 0.1 μM for 10 minutes before ischemia gives maximal cardioprotective effect. 36 Further increase in HU-210 concentration did not enhance its protective effect. HU-210 at high concentration induced a marked bradycardia and decreased hemodynamic function of the isolated perfused rat heart. 49
One possible explanation of these differing conclusions is that the receptors could have developed tolerance to the high concentration of HU-210 causing the protective effect to wane during its prolonged exposure. Another is that HU-210 may protect through a process termed homesis. That is, HU-210 may actually have a noxious effect on the heart which happens to trigger a protective adaptation after its removal, much like ischemia does in ischemic preconditioning. If that is the case, then its continued presence during ischemia might actually be detrimental. In ischemic preconditioning, the protective signals can be divided into triggers and mediators. The triggers start the protective cascade prior to ischemia and their signal does not have to be sustained. An example would be a 5-minute pulse of adenosine, opioid or bradykinin, which puts the heart into a preconditioned state that lasts for about an hour. Mediators exert their protection at the time of reperfusion, and protection relies on their continuous signal during reperfusion. An example of a mediator would be the activation of ERK or inhibition of GSK3β (glycogen synthase kinase-3 Beta), which acts to inhibit mitochondrial permeability transition pore (MPTP) formation during reperfusion. 50 Triggers have to be administered prior to ischemia, while mediators can usually be started just prior to reperfusion which allows their use in the setting of acute myocardial infarction. The above-mentioned studies indicate that the schedule and dose are important, and these clearly need further exploration.
In the experiments on isolated rat heart, the Maslov group induced global ischemia (45 min) and reperfusion (30 min). Pretreatment of the hearts with a solution containing cannabinoid HU-210 (0.1 or 1.0 μM) for 10 min followed by 10 min washout resulted in a 2-fold decrease in CK activity in the coronary effluent. 36 The cardioprotective effect of this compound remained unchanged after NO synthase inhibition, which bracketed the HU-210 infusion. The HU-210 reduced cyclic adenosine monophosphate (cAMP) level in the myocardium by 2-fold during reperfusion but did not affect this parameter before or during ischemia. This cannabinoid also had no effect on the level of cGMP in the myocardium. 36 Thus, the cardioprotective effect of HU-210 is accompanied with a decrease in cAMP level in the myocardium during reperfusion, while cGMP and NO synthase are not involved in HU-210 triggering this effect ex vivo.
Another study was performed on mice with in situ coronary artery occlusion/reperfusion revealed that injection of the CB1/CB2 receptor agonist WIN 55,212-2 30 min prior to ischemia promotes a decrease in the IS/AAR ratio. This effect was maintained after the blockade of CB1 receptor by AM215 but disappeared after perfusing the heart with the selective CB2 antagonist AM630. 39 They concluded that the CB2 receptor was the protective one. AM630 produced a slight but significant (P < 0.05) increase in the IS/AAR ratio compared with the vehicle alone, while AM215 had no effect on the IS/AAR index. Ideally, the blocker should have no effect on the injury to the untreated heart. Perhaps, AM630 aggravates ischemia/reperfusion injury independent of its binding to CB2 receptor, and the CB2 receptor was not actually involved. But that does not explain the lack of effect of AM215. Infarct-reducing effect of WIN 55,212-2 was accompanied by a decrease in the activity of myeloperoxidase in cardiac tissue. 39 It is known that this enzyme is an indicator of neutrophils invasion in myocardial tissue during ischemia/reperfusion injury of the heart. 51 The authors concluded that CB2 receptor activation protects via a decrease in neutrophil accumulation in myocardium. It is possible that protection was associated with other mechanisms and the decreased neutrophil content simply reflected less tissue necrosis. Cause and effect are difficult to determine in this type of experiment, however, the fact that WIN 55,212-2 is protective in neutrophil-free isolated hearts would argue for the second hypothesis. 38
The above-mentioned data are strongly supported by the results of Wang et al 5 who also used mice. Hearts were subjected to either 30 min coronary artery occlusion and 24 h reperfusion or 24 h of continuous occlusion. The highly selective CB2 agonist HU-308 was administered intraperitoneally (2 mg/kg) 2 hours prior to ischemia and decreased the IS/AAR ratio 2-fold in the reperfused group. This effect of HU-308 was reversed by pretreatment with the CB2 selective antagonist AM630. 5 Perhaps mouse hearts are fundamentally different than rat hearts. In the continuous occlusion group, the IS/AAR ratio was also decreased 1.5-fold. This last result is the most surprising because, according to a widespread opinion, it is impossible to achieve a decrease in infarct size without reperfusion in a species that has negligible coronary collateral blood flow like the mouse. In this case, pharmacological agents or a preconditioning protocol can only delay cell death. 52 –54 The time required for ischemia to kill all of the tissue in the ischemic zone in experimental animals fluctuates between 3 and 6 hours. 52 –55 Accordingly, after 6 hours of ischemia, reperfusion does not promote a decrease in infarct size. 53,55 These data are also confirmed by the clinical observations. 56 A cannabinoid may delay cell death for an hour or two, but it is difficult to imagine that it could maintain viability for 24 hours in the absence of blood flow. The results of permanent occlusion shown by Wang’s group are difficult to explain.
CB2 protection has been reported in rats as well. Vogel’s group found that Δ9-THC could increase tolerance of rat neonatal cardiomyocytes to hypoxia. 10 Hypoxia was produced by incubating the cardiomyocytes in an argon-filled chamber with a glucose-free medium for 60 minutes. 10 The cannabinoid Δ9-THC was added to the medium 24 h before the hypoxia exposure. Hypoxia evoked lactate dehydrogenase (LDH) leakage from cardiomyocytes indicating cardiac cell necrosis. The Δ9-THC reduced LDH release. When the selective CB2 receptor antagonist SR144528 was added 1 h before incubation with Δ9-THC, protective effect was absent. 10 Since the selective CB1 receptor antagonist SR141716 did not attenuate the cytoprotective effect of Δ9-THC, they concluded that the CB2 receptor caused the protection. It has been reported that neonatal cardiomyocytes only express CB2 receptors 10 which is consistent with the above-mentioned observation.
Another study examined the interaction between CB1 and CB2 receptors and the mechanism of their cardioprotective effects. 30 Isolated perfused rat hearts were subjected to 2 h low-flow ischemia (flow rate 0.6 mL/min) and 20 min reperfusion. Necrosis was estimated by measuring of LDH and CK levels in the coronary effluent. One of the following (CB1/CB2/GRP55 receptor agonist anandamide, the selective GPR55 agonist PEA, or the CB1 receptor agonist 2-AG) was added to the perfusion solution 15 min before ischemia: and perfused throughout the ischemic period. Both PEA and 2-AG decreased LDH and CK levels in coronary effluent, while anandamide had no effect on these parameters. The CB2 receptor antagonist SR144528 reversed the cytoprotective effect of PEA and 2-AG. The CB1 receptor antagonist SR141716A abolished the cardioprotective effect of 2-AG. The authors concluded that cardioprotection may be achieved via stimulation of both CB1 and CB2 receptors. It should be noted, however, that this conclusion was challenged when it was later found that PEA has no significant affinity for either CB1 or CB2. In another publication from this group, isolated perfused rat hearts were subjected to 90-minute low-flow ischemia and 60-minute reperfusion. This time, more selective cannabinoid agonists (ACEA and JWH015) were present in the perfusion solution for 5 minutes before ischemia and throughout the ischemic period. 31 Both the selective CB1 receptor agonist ACEA (50 nM) and the selective CB2 receptor agonist JWH015 (50 nM) decreased the IS/AAR ratio 30,31 again supporting the hypothesis that both CB1 and CB2 can be involved in the cardioprotective effects of cannabinoids. Interesting that the infarct-reducing effect of ACEA was not observed after inhibition of NO synthase by NG-nitro-L-arginine. However, this NO synthase inhibitor did not attenuate the cardioprotective effect of JWH015, 31 suggesting that the cardioprotective effect of CB1, but not CB2 receptor stimulation, is mediated by endothelial NO synthase.
Both CB1 and CB2 Receptors are Present in the Heart
Some of the data presented earlier contradict our results that the infarct-reducing effect of HU-210 depended on CB1 but not CB2 receptor stimulation. 29,33 The expression of these receptors may differ among the various models. Vogel’s group found that the CB1 receptor is absent in cardiomyocytes of newborn rats, 10 indicating that the age of animals plays an important role. Message for the CB1 receptor has been reported to be in human myocardium. 7 Wagner et al found evidence of CB1 receptors in myocardium of adult rats. 4 Two years later, they confirmed an existence of this receptor in the adult rat heart by immunohistochemistry. 17 However, they could not find CB2 receptors in the myocardium of adult rats. Brown et al reported that mRNA for the CB2 receptor could not be found in the myocardium of adult rats. 57 In contrast, a Canadian group of investigators reported that mRNA and proteins of both CB1 and CB2 receptors are present in the heart of adult rats. 31 They have shown that CB1 receptors are localized almost exclusively on the arterial and capillary endothelial cells in the heart, whereas CB2 receptors are expressed in cardiomyocytes and endothelial cells of larger arteries. 31 Recently, both CB1 and CB2 receptors were found in the tissue of left ventricle of mice by Western blot analysis. 5
Much of the differences among the above-mentioned studies may well reflect differences in the various models of ischemia/reperfusion: in situ versus isolated hearts, whole hearts versus isolated myocytes, and mice versus rats. The dose and schedules of drug administration also vary widely among the studies ranging from pretreatment with the ligands to acute administration just prior to ischemia. In some studies, the drug was withdrawn prior to ischemia, and in others it was present during ischemia. In in vivo studies, the drug probably persisted during reperfusion. It is therefore not surprising that the results will be discrepant. While the experiments on receptor subtypes in myocardium are contradictory, we propose that the weight of evidence supports the hypothesis that under certain circumstances, cannabinoids can protect the hearts of adult animals through occupancy of CB1 and CB2 receptors.
Delayed Preconditioning
Protection from ischemic preconditioning wanes after an hour or two, but a second window of prolonged protection appears 24 h later. 58 This phenomenon was termed “delayed” ischemic preconditioning. The protection involves increased myocardial NO production via expression of inducible NO synthase (INOS). 59 Prolonged exposure to Δ9-THC mimics delayed ischemic preconditioning by increasing INOS content 4-fold. 10 We (Zhang group) have recently obtained data showing that the endogenous cannabinoid anandamide can mimic delayed ischemic preconditioning. 60 We found that anandamide, administered to rats intravenously (1 mg/kg), induced expression of heat shock protein 72 (HSP-72), which peaked 24 h later. Induction of HSP-72 is also a prominent feature of delayed ischemic preconditioning. 58,61 We induced 30 min occlusion of left anterior descending coronary artery (LAD) followed by 120 min reperfusion in vivo 24 h after the administration of anandamide and found a 2-fold decrease in the IS/AAR ratio. This cardioprotective effect was reversed by pretreatment with the CB2 receptor antagonist AM630 but not with the CB1 receptor antagonist AM251 indicating a CB2 effect. Anandamide’s elevation in HSP-72 was also eliminated by AM630 but not by AM251.
It was recently reported that pretreatment of mice intraperitoneally with an ultra-low dose (2 μg/kg) of Δ9-THC 48 or 24 h prior to coronary artery occlusion is cardioprotective. 62 They showed that this cannabinoid decreases infarct size, elevates fractional shortening, reduces troponin T leakage to the blood, and decreases the accumulation of neutrophils to the infarct area. Unfortunately, the authors did not measure area at risk in these hearts, which reduces the accuracy of estimation of the actual extent of the infarct size reduction induced by Δ9-THC. The current data indicate, however, that both anandamide and Δ9-THC can mimic delayed preconditioning in the intact heart.
Cannabinoids and Apoptosis
Recently, we (Zhang group) found that intravenous administration of the CB2 receptor agonist JWH133 (20 mg/kg) in rats prior to coronary artery occlusion and reperfusion decreases the apoptotic index in the area at risk 3-fold. 42 Pretreatment with the CB2 receptor antagonist AM630 abolished antiapoptotic effect of JWH133 implicating CB2 receptors. We established that ischemia/reperfusion increases caspase 3 and caspase 9 in the myocardium. Injection of JWH133 promoted a decreased caspase 3 approximately 5-fold and caspase 9 twelve-fold. 42 It is well known that opening of MPTP is a trigger of apoptosis. 63,64 Opening of this pore reduces potential on the inner mitochondrial membrane (Δψ) and releases cytochrome C and other apoptosis-inducing factors from the intermembrane space into the cytosol. 63,64 Cytochrome C together with apoptosis protease activating factor 1, procaspase 9, and adenosine triphosphate (ATP) form a supramolecular complex called the apoptosome. 64 Apoptosome catalyzes the proteolysis of procaspase 9, which results in transformation of procaspases to their active forms. Caspases catalyze the cleavage of other proteins, which ultimately leads to apoptosis. In the in vivo experiments on rats, we found that the CB2 receptor agonist JWH133 resulted in a 2.5-fold decrease in cytochrome C level and inhibited the fall in Δψ. Blocking CB2 with AM630 eliminated these effects. 42 We propose that the antiapoptotic effect of JWH133 is an outcome of MPTP inhibition via CB2 receptor stimulation.
Signaling Pathways of Cardioprotective Effect of Exogenous Cannabinoids
Oxidative stress is thought to play an important role in the pathogenesis of ischemia/reperfusion-induced heart injury. 65,66 Defer et al 27 tested whether cannabinoids can enhance resistance of cardiomyocytes to the detrimental effects of oxidative stress. Cardiomyocytes of wild-type (control) and CB2−/− mice were incubated for 12 h in the presence of H2O2 which induced death of approximately 90% of cells in both control and knockout groups. The selective CB2 agonist JWH133 (1 μM) increased cell survival during oxidative stress 5-fold. However, the protective effect was not observed in the presence of the selective CB2 antagonist AM630 (5 μM). 27 These data suggest that the cardioprotective effect of cannabinoids may be the result of an increased tolerance of the cardiomyocyte to oxidative stress. Current thinking is that preconditioning protects by preventing oxidative stress-induced opening of lethal MPTP in the heart’s mitochondria at reperfusion, 67 and the Defner et al. study suggests that cannabinoids protect through a similar mechanism.
A number of studies have shown that protein kinase C (PKC), p38MAPK, mitogen-activated protein kinase (MEK1/2)—extracellular signal-regulated kinase (ERK1/2) and phosphatidylinositol 3-kinase (PI3K)—Akt pathways are involved in the mechanisms of protection of ischemic preconditioning and its pharmacological mimetics. 68 –70 Lépicier et al investigated the signaling mechanism of the cytoprotective effect of PEA using a model of low-flow ischemia and reperfusion on isolated rat heart. 30 Pretreatment of the hearts with the PKC inhibitor chelerythrine, the MEK1/2 inhibitor PD98059, or the p38MAPK blocker SB203580 reversed the cardioprotective effect of PEA. 30 Furthermore, they showed that this endogenous cannabinoid induced phosphorylation of ERK1/2. 30 These data are consistent with our (Maslov group) study, which indicated that PKC is involved in the cardioprotective effect of HU-210. 29,33,41 One hour after injection of anandamide, which is able to mimic the effect of delayed ischemic preconditioning, resulted in an 8-fold increase in Akt phosphorylation, but 24 h later, Akt phosphorylation had returned to its control value. 60 Pretreatment of isolated rat hearts with the PI3K inhibitor wortmannin, Akt inhibitor MK-2206, and the CB2 receptor antagonist AM630 all attenuated early anandamide-induced phosphorylation of Akt and delayed expression of HSP-72. 60 More recently, we have found that JWH133 during in vivo ischemia/reperfusion promoted Akt phosphorylation (Figure 1) and had an anti-infarct and antiapoptotic effect in rat hearts. Pretreatment with wortmannin abolished the protection showing that Akt had a functional role in the protection. 42
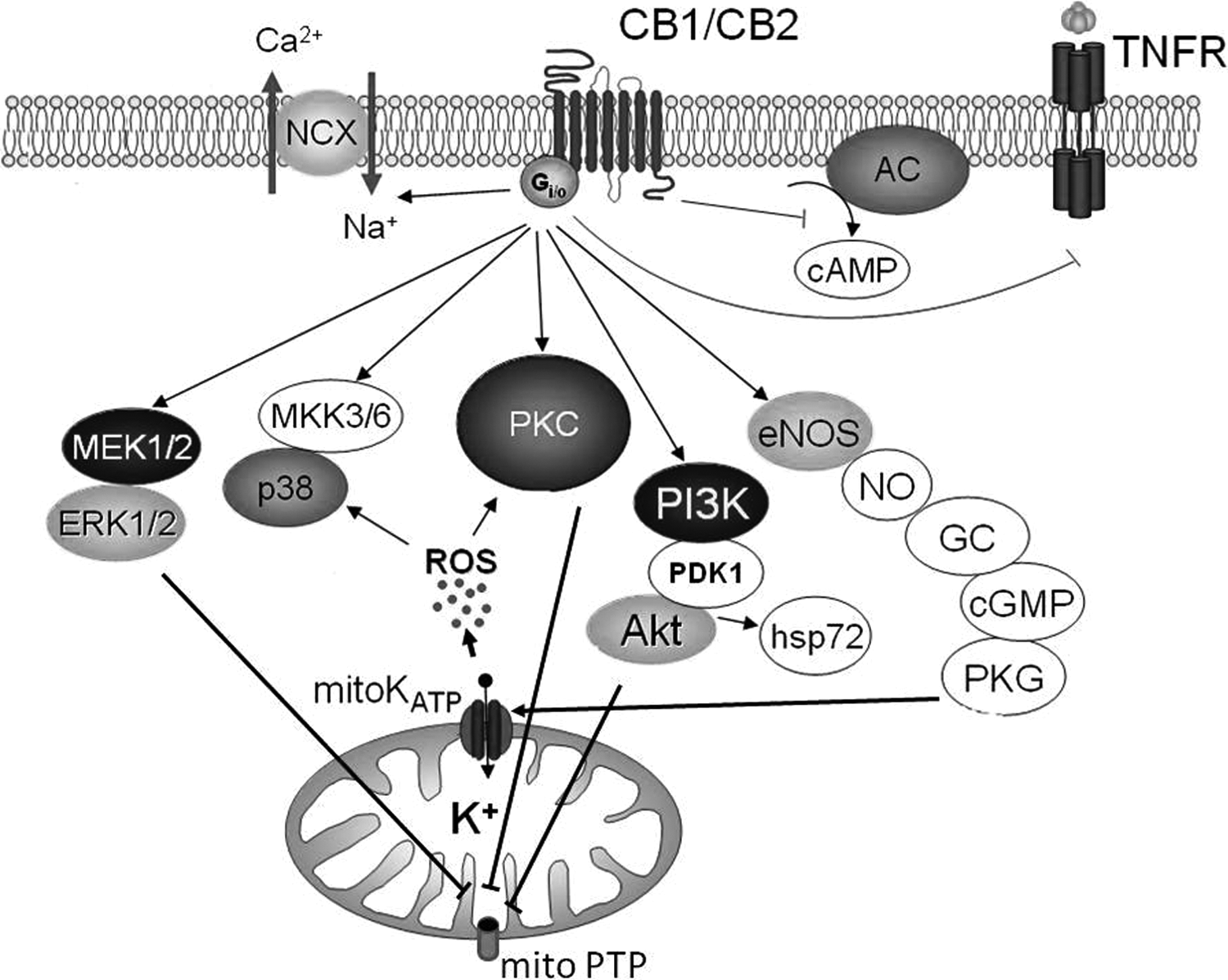
Proposed signaling scheme for the cardioprotective effect of cannabinoids. CB indicates cannabinoid receptor; AC, adenylyl cyclase; NCX, Na+/Ca2+ exchanger; TNFR, tumor necrosis factor receptor; mitoKATP, mitochondrial ATP-sensitive K+ channel; PKC, protein kinase C; ROS, reactive oxygen species, p38 MAPK, p38 MAP kinase; MEK1/2, mitogen-activated protein kinase; ERK1/2, extracellular signal-regulated kinase; PI3K, phosphatidylinositol 3-kinase; PDK1/2, 3’-phosphoinositide-dependent kinase-1/2; Akt, kinase isolated from AKR thymoma cell line; hsp72, heat shock protein; eNOS, endothelial NO-synthase; NO, nitric oxide; GC, guanylyl cyclase; cGMP, cyclic guanosine monophosphate; PKG, protein kinase G; MKK3/6, mitogen-activated protein kinase 3 and 6; mitoPTP, mitochondrial permeability transition pore. See text for details.
Recent data have suggested that an alteration in proinflammatory cytokines may also be involved in the cardioprotective effect of cannabinoids. This study revealed that 30-minute coronary artery occlusion and 24 h reperfusion or 24 h permanent occlusion evoked a 1.5-fold increase in the level of tumor necrosis factor α (TNF-α) in the blood of mice. 5 Intraperitoneal injection of the CB2 agonist HU-308 prior to coronary artery occlusion in mice abolished the rise in TNF-α. However, pretreatment with the CB2 receptor antagonist AM630 reversed the effect of HU-308. 5 Tumor necrosis factor α is known to be involved in the pathogenesis of inflammatory heart damage induced by ischemia/reperfusion. 71 These data support the hypothesis that a decrease in TNF-α is involved in HU-308-induced protection.
The above-mentioned data suggest that NO synthase could be involved in the cardioprotective effects of cannabinoids. Let us try to compare and analyze these data. It was established that the NO synthase inhibitor L-NAME abolished delayed cardioprotective effect of Δ9-THC. 10 Furthermore, it was found out that Δ9-THC increased NO production 3-fold and enhanced expression of INOS 24 hours after the pretreatment with Δ9-THC. 10 The early infarct-reducing effect in isolated rat heart of the selective CB1 agonist ACEA was reversed by coinfusing the NO-synthase inhibitor NG-nitro-L-arginine. However, this NO synthase inhibitor did not affect cardioprotective effect of the selective CB2 agonist JWH015. 31 We (Maslov group) also found that the inhibition of NO synthase with L-NAME did not affect the HU-210-induced increase in cardiac tolerance to ischemia and reperfusion in vivo 41 or ex vivo. 33 Thus, we conclude that NO synthase is involved in cannabinoid-induced delayed preconditioning and is suggested to be involved in the CB1 induced early preconditioning but is clearly not involved in CB2-induced early preconditioning.
Opening of the ATP-sensitive K+ channel (KATP) channel is an important step in the signaling of ischemic preconditioning. 72 We (Maslov group) have found that 10 min perfusion of isolated rat heart with buffer containing HU-210 (100 nM) 20 min prior to global ischemia is able to decrease CK release during reperfusion. 33 Pretreatment with the KATP channel blocker glibenclamide abolished the cardioprotective effect of HU-210. This indicates that the KATP channel is also involved in cannabinoid-induced cardioprotection.
It has been revealed that an enhancement of Na+/Ca2+ exchange (NCX) during ischemia/reperfusion aggravates ischemia/reperfusion heart injury. 73 Not surprisingly, inhibition of NCX during ischemia is highly protective. 73,74 Recently, in experiments on isolated rat cardiomyocytes, we (Zhang group) have established that anandamide (1-100 nM) dose dependently suppressed the increase in NCX. 75 Furthermore, anandamide (100 nM) significantly attenuated the increase in [Ca2+]i during simulated ischemia. Pretreatment of the cardiomyocytes with the CB1 receptor antagonist AM251 failed to inhibit the effects of anandamide on NCX and [Ca2+]i in these experiments. Meanwhile, the CB2 receptor antagonist AM630 eliminated the effects of anandamide on NCX and [Ca2+]i during simulated ischemia. The CB2 receptor agonist JWH133 (100 nM) also suppresses the increase in NCX and [Ca2+]i during simulated ischemia. In addition, pretreatment of the cells with the Gi/o inhibitor pertussis toxin abolished the above-mentioned effects of anandamide and JWH133. 75 These findings indicate that activation of Gi/o-protein coupled CB2 receptor inhibits NCX and prevents Ca2+ overload. It is well known that Ca2+ is one of the triggers of MPTP opening 64 and represents a major source or reperfusion injury in the heart. 73 These results imply that cannabinoid-induced NCX inhibition during ischemia/reperfusion may also contribute to the protection against ischemia/reperfusion.
Concluding Remarks
In summary, current data indicate that cannabinoids mimic the infarct-reducing effect of early preconditioning, postconditioning, and delayed preconditioning and are involved in at least some forms of remote preconditioning of the heart. Cannabinoids also exhibit an antiapoptotic effect. The acute cardioprotective effect of cannabinoids can be mediated via activation of PKC, ERK1/2, and p38 MAPK. The delayed cardioprotective effect of anandamide is mediated via stimulation of PI3K-Akt signaling pathway and enhancement of HSP-72 expression. As in preconditioning, the KATP channel is also involved in the cardioprotective effect of cannabinoids. The delayed cardioprotective effect induced by a cannabinoid Δ9-THC is mediated via enhancement of inducible NO synthase expression. However, unlike the case with ischemic preconditioning, data on the involvement of NO synthase in the acute cardioprotective effect of cannabinoids are contradictory. Anandamide and JWH133 inhibit NCX via CB2 receptor activation that may be related to the antiapoptotic effect of cannabinoids. In conclusion, agonists of CB receptors can be considered as an important group of chemical compounds that should be further explored for the creation of drugs that prevent injury from ischemia/reperfusion in the clinical setting.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors (Leonid N. Maslov, Andrey V. Krylatov and Natalia Naryzhnaya) were supported by Russian National Foundation grant 14-15-00008.