
Abstract
Dantrolene, which is primarily used to treat malignant hyperthermia, has recently been suggested for the prevention of arrhythmogenesis in various animal models. In this study, the effects of dantrolene treatment on electrophysiological properties and ventricular arrhythmias (VAs) in rats with chronic β-adrenergic receptor (β-AR) activation were investigated. Rats were randomized to treatment with saline (control group), isoproterenol (ISO; ISO group), or ISO + dantrolene (ID group) for 2 weeks. An electrophysiological study was performed to assess action potential duration restitution (APDR) and induce action potential duration (APD) alternans or VA in vitro. The protein levels of Cav1.2, sarcoplasmic reticulum Ca2+-ATPase (SERCA2a), and ryanodine receptor 2 (RyR2) were detected by Western blot. Compared with the control group, chronic administration of ISO significantly increased APD, the maximum slope (Smax) of APDR curve, and the spatial dispersions of Smax and APD (all P < .01), and all effects were attenuated by dantrolene treatment (all P < .05). Additionally, chronic ISO administration significantly reduced the protein levels of SERCA2 and RyR2, but increased the Cav1.2 protein expression (all P < .05). However, compared with the ISO group, dantrolene treatment preserved SERCA2a and RyR2 protein levels and decreased Cav1.2 protein levels in the ID group (all P < .05). The intracellular Ca2+ ([Ca2+]i) levels measured by incubating isolated cardiomyocytes with Fluo-3/alveolar macrophages were significantly increased in the ISO group compared with the control group (P < .01). Dantrolene treatment markedly reduced the rise of [Ca2+]i levels caused by chronic administration of ISO (P < .05). Dantrolene treatment also prevented the reductions in the APD alternans and VA thresholds induced by chronic ISO stimulation (all P < .05). These data suggest that dantrolene stabilizes ventricular electrophysiological characteristics and increases the expression of key sarcoplasmic reticulum calcium cycling proteins to reduce vulnerability to VA in rats with chronic β-AR activation.
Introduction
Chronically elevated catecholamine levels have been detected in several cardiovascular diseases, such as congestive heart failure and acute myocardial infarction. 1,2 In these cardiovascular diseases, elevated catecholamine can activate the cardiac β-adrenergic receptor (β-AR), which is a powerful regulator of cardiac function. Although the initial activation of β-AR by catecholamine is beneficial to cardiovascular function, its long-term effects on the heart are harmful. In animals with β-AR overactivation, distinct alterations in myocardial structure and in contractile and electrophysiological properties have been observed. 3 –5 As a nonselective β-AR agonist, isoproterenol (ISO) has been widely used to investigate the effects of activating cardiac β-AR in experimental studies. The ISO acts through the β-AR/cyclic adenosine monophosphate/protein kinase A pathway to increase intracellular Ca2+ ([Ca2+]i) levels and eventually lead to [Ca2+]i overload. 6,7 There is evidence to suggest that the aberrant [Ca2+]i homeostasis is a contributor not only to cardiac dysfunction but also to arrhythmogenesis. 8,9 Thus, certain therapeutic interventions to restore the proper [Ca2+]i regulation may potentially prevent the cardiotoxic effects induced by chronic β-AR activation.
Dantrolene, a drug primarily used to treat malignant hyperthermia, has recently been shown to improve cardiac function and prevent arrhythmogenesis via stabilization of ryanodine receptor 2 (RyR2) function and restoration of [Ca2+]i homeostasis in various animal models. 10 –12 Furthermore, a previous study demonstrated that dantrolene treatment has a significant role in protecting the heart against ISO-induced myocardial injury. 13 However, the effects of dantrolene treatment on electrophysiological properties and ventricular arrhythmia (VA) in animals with chronic β-AR activation remain unclear.
In the present study, we mimicked β-AR overactivation using chronic administration of ISO and examined the effects of dantrolene treatment on ventricular repolarization, action potential duration restitution (APDR), threshold levels of action potential duration (APD) alternans or VA, [Ca2+]i, and expression of key sarcoplasmic reticulum (SR) calcium cycling proteins in rats.
Materials and Methods
Experimental Animals
All experimental procedures conformed to the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication #85-23, revised 1996), and the protocol was approved by the Animal Care and use Committee of Renmin Hospital of Wuhan University, China.
Adult male Sprague-Dawley rats (weighing from 200 to 250 g) from the Experimental Animal Center of the Medical College of Wuhan University were used in this study. The animals were randomly divided into 1 of the following 3 groups: the control group where rats received an equal volume of 0.9% saline; the ISO group where rats received ISO alone (5 mg/kg/d, intraperitoneally [ip]); or the ISO + dantrolene group (ID group) where rats received ISO (5 mg/kg/d, ip) and dantrolene (2 mg/kg/d, ip). Drugs or saline treatments were administered for 14 consecutive days (1/3 total doses every 8 hours). Dantrolene treatment was initiated 30 minutes prior to ISO administration.
Surface Electrocardiogram Recording and Echocardiography Analysis
After the 2-week drug treatment period, surface electrocardiogram (ECG) recordings (lead II) were obtained. Rats were anesthetized with pentobarbital sodium (40 mg/kg, ip) and electrodes were positioned subcutaneously. Chart 7.0 software (AD Instruments, Bella Vista, New South Wales, Australia) was used to analyze the recorded ECG data. Means were calculated for the RR interval, QRS duration, and QT interval. The QT interval was corrected for heart rate using Bazett formula 14 QTc = QT/(RR/100)1/2.
Additionally, echocardiography analysis was also performed to assess left ventricular ejection fraction (LVEF), left ventricular endocardial fractional shortening (LVEFS), left ventricular end-diastolic diameter (LVEDD), left ventricular end-systolic diameter (LVESD), left ventricular posterior wall dimensions (LVPWD), and interventricular septum thickness at diastole (IVSD) in this study.
Histological Analysis
Rat hearts (n = 5 in each group) were excised and transversely cut close to the papillary muscles to visualize the left ventricles. Heart tissue sections were fixed in 10% phosphate-buffered formalin and embedded in paraffin. Paraffin sections were stained with Masson trichrome to visualize collagen deposition. Fibrosis was quantified in each heart section as the average myocardial interstitial collagen volume fraction (CVF) in 4 random areas of the left ventricular free wall. The CVF was measured by 2 independent observers who were blinded to experimental group assignment, and the mean CVF for each sample was calculated.
Isolation of Ventricular Myocytes and Measurement of [Ca2+]i Levels
As previously described, 15 the ventricular myocytes were obtained from rat hearts (n = 5 in each group) through enzymatic digestion with collagenase. The dissociated cells were used for experiments within 5 hours of preparation. 16 To avoid the cell clumping during the flow cytometry studies, the isolated myocytes were filtered with a 300-μm filter. In addition, cells were stained with propidium iodide (5 μg/mL) to exclude dead cells. 17 Intracellular Ca2+ levels were measured as previously reported. 15,16,18 In brief, [Ca2+]i levels were determined using the Ca2+-sensitive fluorescence probe, Fluo-3/alveolar macrophage (fluorescence lifetime analysis [FLA]; Molecular Probes; Invitrogen Company, California) by flow cytometry (FACScan, Becton-Dickinson, Massachusetts). The myocytes were incubated with 10 μmol/L FLA at 37°C for 60 minutes. Cells were then washed 3 times with Ca2+-free phosphate-buffered saline to remove extracellular FLA before resuspension in Ca2+-free media supplemented with 10 mg/mL bovine serum albumin (Gibco Company, California; 145 mmol/L NaCl, 5 mmol/L KCl, 1.0 mmol/L MgCl2, 1.0 mmol/L ethylene glycol tetraacetic acid [EGTA], 5.6 mmol/L glucose, and 10 mmol/L HEPES, pH 7.4 adjusted with NaOH). Probenecid (2 mmol/L) was added to the media to prevent loss of FLA via the anion transporter. 19 During flow cytometry, FLA fluorescence intensity was measured with a spectrophotometer (Hitachi F-4500, Japan) by exciting the cells at a wavelength of 488 nm and recording the emission signal at a wavelength of 530 nm. Approximately 2 × 103 cells in each sample were analyzed to calculate the mean fluorescence intensity (Fmean) of FLA, which is positively correlated with the [Ca2+]i level. Additionally, the average [Ca2+]i concentration was calculated by the following equation 16,18,19 : [Ca2+]i = Kd[(F − Fmin)/(Fmax − F)], where Kd is the dissociation constant of FLA (400 nmol/L at 37°C). 19 Fmax is the maximal Fmean obtained by adding 0.1% Triton X-100 and 1 mmol/L CaCl2, and Fmin is the minimal Fmean determined by adding 10 mmol/L EDTA. F is the Fmean measured without the addition of Triton-X-100 and CaCl2 or EGTA.
Western Blotting Analysis
Rat hearts (n = 5 in each group) were immediately excised following euthanasia. The left ventricular tissue was then collected and stored in tubes at −80°C until Western blotting analysis was performed. Western blotting was performed to determine the relative expression levels of Cav1.2, RyR2, sarcoplasmic reticulum Ca2+-ATPase (SERCA2a), and phospholamban (PLB). In this study, the protein levels were normalized to glyceraldehyde 3-phosphate dehydrogenase.
Preparation of Langendorff-Perfused Hearts
The rats were anesthetized with pentobarbital sodium (40 mg/kg, ip) and heparinized with heparin sodium (400 U, ip). After a thoracotomy, the heart was quickly isolated, rapidly cannulated in a Langendorff perfusion system (AD Instruments) and secured for retrograde perfusion with 37.0°C ± 0.5°C oxygenated Tyrode solution at a rate of 6 to 8 mL/min. The composition of Tyrode solution was as previously described. 20 Prior to further analysis, the isolated hearts were perfused for 20 minutes. Those hearts that had irreversible myocardial ischemia or did not recover regular spontaneous rhythm were discarded. A total of 36 hearts (n = 12 in each group) were used for further experimental procedures in this study.
Data Acquisition and Stimulation Protocol
Monophasic action potential recording
The monophasic action potential (MAP) was recorded in 4 epicardial sites using a custom-made Ag-AgCl electrode, which was comprised of two 0.25-mm Teflon-coated silver wires (99.99% purity). 20 The 4 recording sites included the left ventricular anterior and posterior base and the left anterior apex and left posterior apex (LPA). All MAP signals were recorded and analyzed by the PowerLab system and Chart 7.0 software (AD Instruments). The amplifier bandwidth signal was between 0.3 and 1 kHz.
S1 dynamic pacing, inducibility of APD alternans or VA, and construction of APDR curves
The paired platinum-stimulating electrodes were positioned on the basal surface of the right ventricle, and the pacing stimuli were 2-ms pulse widths with 3 times the diastolic threshold current. The S1-S1 pacing protocol was performed with a series of pulse trains at a regular pacing cycle length (PCL). Starting at 300 ms, the PCL was shortened to 160 ms in 20-ms intervals and then to 40 ms in 10-ms intervals. The regular pacing at each PCL lasted for 15 seconds to ensure a steady rhythm, and each pacing was separated by at least 30 seconds to minimize the pacing memory. The VA episode was defined as lasting >2 seconds. 4 If VA was induced, a 5- to 10-minute rest period after VA termination allowed for cardiac electrical recovery. The APD at the PCL of 300 ms in each site was determined at 90% repolarization (APD90) to calculate the average APD90 of the left ventricle. The APD alternans was assessed at each site by subtracting the APD90 for 2 consecutive beats when the alternate APD90 differed by >5% over 10 beats. 21 The threshold was defined as the maximal PCL (PCLmax) that induced APD alternans or VA by PCL decrease. When the PCL was decreased to 40 ms, but the APD alternans or VA was not elicited, the threshold of APD alternans or VA was defined as less than 40 ms.
The dynamic APDR curves were constructed by plotting each APD90 against the preceding diastolic interval (DI), and the data points were fitted with a single exponential trend as in previous reports. 21,22 The DI was measured between the APD90 point in the preceding MAP and the onset of the subsequent MAP. The maximal slopes (Smaxs) of the APDR curves were determined at the shortest DI. Traces in which a reproducible sequence of consistently shaped waveforms could not be obtained were not used to construct APDR curves.
Statistical Analysis
SPSS 18.0 for Windows (SPSS Inc, Chicago, Illinois) was used for the statistical analyses. The coefficients of variation (COVs, standard deviation [SD]/mean) of the APD90 and Smax curves were calculated to represent the spatial dispersions of APD90 and Smax in the left ventricle. Continuous data, that is, APD90, COV, and CVF, were expressed as the mean ± SD and analyzed by 1-way analysis of variance followed by Tukey post hoc test to determine the differences among groups. The median threshold values of APD alternans or VA were compared by the nonparametric Wilcoxon signed rank test or the Kruskal-Wallis test as appropriate. The nonlinear curve fit to construct the APDR curve was created with Origin7.5 (Microcal Co Inc, Northampton). A value of P < .05 was considered statistically significant.
Results
Changes in Cardiac Structural and Contractile Function
Table 1 summarizes the body and organ weights of the rats in each group. With respect to body weight, no differences were found among the 3 groups (all P > .05). However, chronic ISO administration significantly increased the heart weight (HW) and left ventricular weight (LVW; all P < .01 compared to the control group). By contrast, significant reductions in HW and LVW were found in the ID group when compared to the ISO group (all P < .01). Additionally, echocardiography showed that chronically ISO-stimulated rats exhibited significantly increased LVEDD, LVESD, IVSD, and LVPWD, while the LVEF and LVEFS values were significantly decreased (all P < .01, compared to the control group). However, dantrolene administration ameliorated the abnormal echocardiographic indices induced by chronic ISO stimulation (all P < .05; Table 1). Consistent with these results, histopathology of the heart sections showed that the average myocardial interstitial CVF was also markedly increased in the ISO group when compared with the control group (P < .01). In contrast, the CVF was lower in the ID group than in the ISO group (P < .01; Figure 1).
The ECG, Anatomic, and Echocardiographic Parameters in Each Group.

Abbreviations: BW, body weight; ECG, electrocardiogram; HR, heart rate; HW, heart weight; ID, ISO + dantrolene; ISO, isoproterenol; IVSD, interventricular septum thickness at diastole; LVEDD, left ventricular end-diastolic diameter; LVEF, left ventricular ejection fraction; LVEFS, left ventricular endocardial fractional shortening; LVESD, left ventricular end-systolic diameter; LVPWD, left ventricular posterior wall dimensions; LVW, left ventricular weight.
a P < .01 versus control group.
b P < .05 versus ISO group.
c P < .01 versus ISO group.
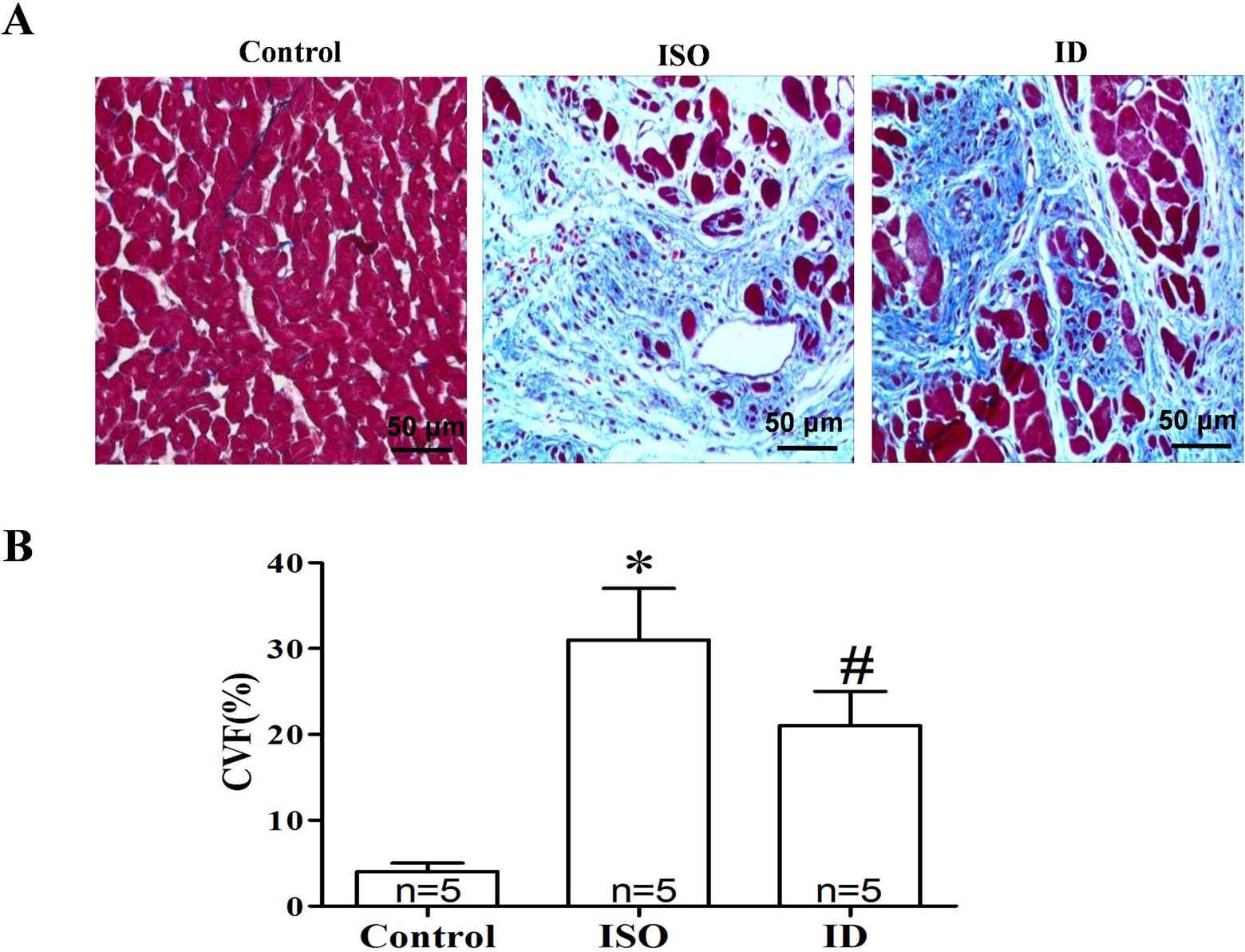
Histological analysis of myocardial fibrosis in 3 groups. A, Masson trichrome stain shown (magnification of ×400) in the left ventricles; (B) summarized data of myocardial fibrosis in each group. Chronic ISO administration significantly increased the myocardial interstitial average collagen volume fraction (CVF) in the ISO group. In contrast, dantrolene treatment markedly reduced the CVF in the ISO + dantrolene (ID) group.*P < .01 versus control group; # P < .01 versus ISO group. ISO indicates isoproterenol.
Characteristics of Ventricular Repolarization
Compared with the control group, the ISO group displayed abnormal ECG waveform morphologies, including widened QRS complexes and prolonged QTc and RR and QT intervals (all P < .01). However, dantrolene treatment markedly attenuated these ECG alterations in the ID group compared to the ISO group (all P < .01; Figure 2 and Table 1). Additionally, chronic ISO administration significantly increased the average APD90 and COV-APD90 (all P < .01). In contrast, treatment with dantrolene significantly reduced the average APD90 and COV-APD90 in rats that received chronic ISO administration (all P < .01; Figure 3).

Three representative electrocardiograms recorded from each group showed the measurements of QRS complex and QT intervals. Dantrolene treatment prevented the electrocardiogram (ECG) waveform abnormalities in the chronically isoproterenol (ISO)-stimulated rats.

The comparisons of ventricular repolarization among 3 groups. A, Three representative APD recorded from each group; (B) summarized data of average APD90 in 3 groups; (C) the APD90 at 4 recording sites of each group; and (D) the spatial dispersion of APD90 was evaluated by coefficient of variations (COV-APD90) among 4 recording sites. Dantrolene treatment significantly reduced the increase in APD90 and COV-APD90 induced by chronic ISO administration. *P < .01 versus control group; # P < .01 versus ISO group. APD90 indicates action potential duration at 90% repolarization; LAB, left anterior basic ventricle; LAA, left anterior apex; LPB, left posterior basic ventricle; LPA, left posterior apex; ISO, isoproterenol.
Alterations in [Ca2+]i Levels and the Expression of Key SR Calcium Cycling Proteins
In this study, the [Ca2+]i level was analyzed by the Fmean and [Ca2+]i concentration. Compared with the control group, the Fmean and [Ca2+]i concentration were markedly augmented in the ISO group (all P < .01). However, the ID group exhibited significant decreases in Fmean and [Ca2+]i concentration when compared to the ISO group (all P < .01; Figure 4).

The results of [Ca2+]i level in each group. A, The examples of fluorescence lifetime analysis (FLA) fluorescence intensity in each group; (B and C) the comparisons of average fluorescence intensity (Fmean) and [Ca2+]i concentration in each group. Compared to the control group, both the Fmean and [Ca2+]i concentration were significantly increased in the ISO group. However, dantrolene treatment markedly reduced the Fmean and [Ca2+]i concentration in the ISO + dantrolene (ID) group. *P < .01 versus control group; # P < .01 versus ISO group. [Ca2+]i indicates intracellular Ca2+; ISO, isoproterenol.
Furthermore, as shown in Figure 5, chronic ISO administration significantly reduced the protein levels of SERCA2 and RyR2 but increased the expression of Cav1.2 (all P < .01 compared to the control group). However, compared with the ISO group, dantrolene treatment preserved SERCA2a and RyR2 protein levels and decreased the Cav1.2 protein level in the ID group (all P < .05). There was no significant difference in the PLB protein levels among the 3 groups (all P < .05).

The expression of Cav1.2, RyR2, SERCA2a, and PLB was assayed by Western blotting. Chronic ISO stimulation significantly decreased SERCA2a and RyR2 protein levels, while increased the Cav1.2 protein levels. However, compared with the ISO group, dantrolene treatment preserved SERCA2a and RyR2 protein levels and decreased Cav1.2 protein levels in the ISO + dantrolene (ID) group. No significant differences in the expression of PLB were found among 3 groups (all P > .05). *P < .01 versus control group; # P < .05 and ## P < .01 versus ISO group. ISO indicates isoproterenol; PLB, phospholamban; RyR2, ryanodine receptor 2; SERCA2a, sarcoplasmic reticulum Ca2+-ATPase.
Changes in the APDR Curve and the APD Alternans or VA Thresholds
Figure 6 shows the dynamic APDR curves, and Table 2 summarizes the Smax at the 4 recording sites in each group. Chronic ISO administration remarkably steepened the dynamic APDR curves at each site (all P < .01). In contrast, the dynamic APDR curves were flattened by dantrolene treatment in the ID group compared to the ISO group at each recording site (all P < .01). Chronic ISO administration also increased the COV-Smax compared with the control values (ISO group: 0.18 ± 0.01 vs control group: 0.09 ± 0.01, P < .01). However, dantrolene treatment significantly reduced the increase in COV-Smax induced by chronic ISO administration (ID group: 0.13 ± 0.01 vs ISO group: 0.18 ± 0.01, P < .01).
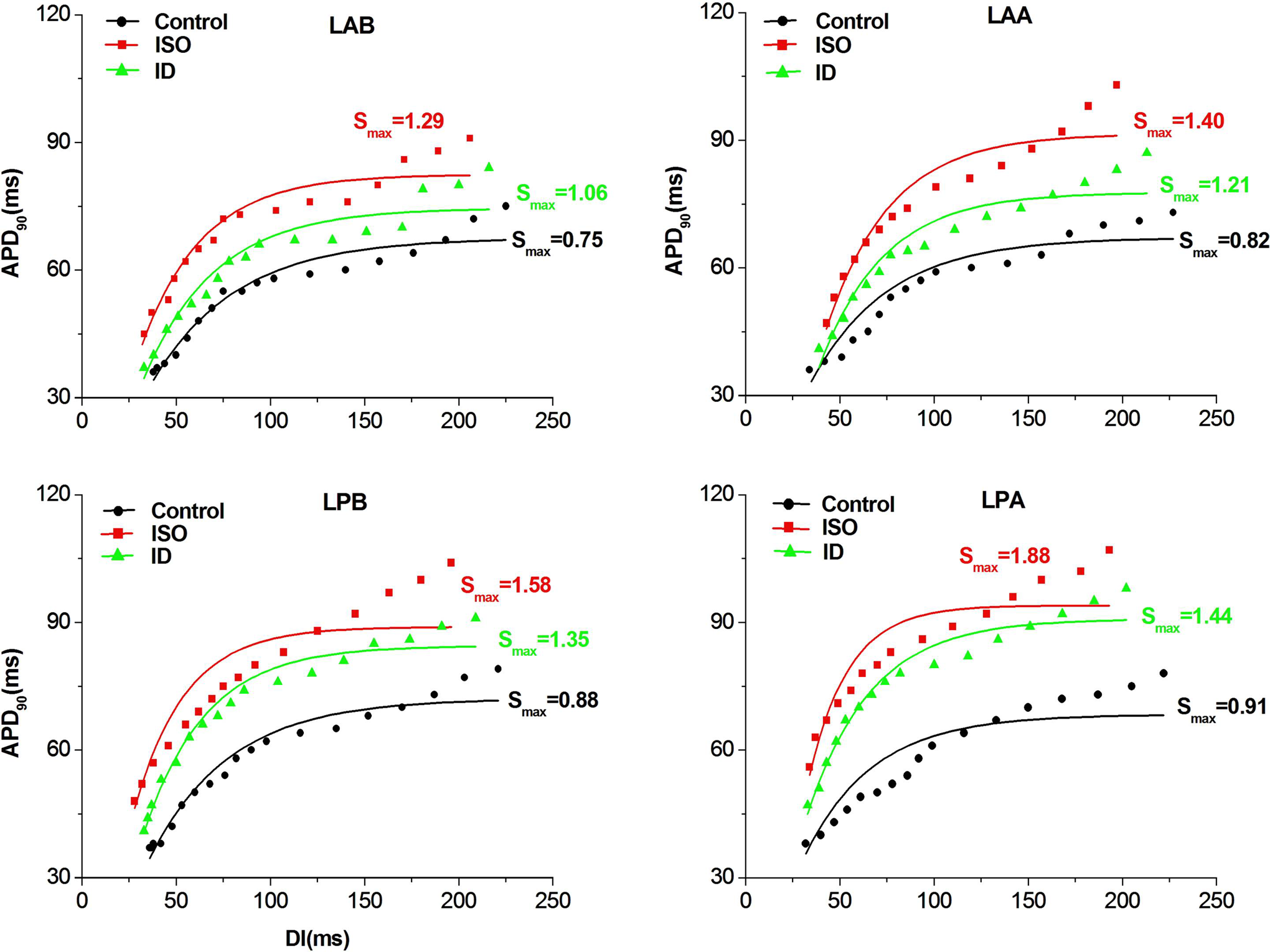
The examples of APD restitution curve in each group. Compared to the control group, the APD restitution curves were shifted upward and steepened in the ISO group at all sites. However, the APD restitution curves were shifted downward and flattened in the ISO + dantrolene (ID) group compared to the ISO group at all sites. ISO indicates isoproterenol.
The Smax of APDR Curve at 4 Recording Sites in Each Group.

Abbreviations: APDR, action potential duration restitution; ID, ISO + dantrolene; ISO, isoproterenol; LAA, left anterior apex; LAB, left ventricular anterior base; LPA, left posterior apex; LPB, left ventricular posterior base; Smax, maximum slope.
a P < .01 versus control group.
b P < .01 versus ISO group.
Meanwhile, the ISO group exhibited significant increases in the PCLmax inducing APD alternans or VA at each recording site (all P < .01 compared to the control), indicating that the APD alternans and VA thresholds were markedly reduced by chronic ISO administration. However, the PCLmax inducing APD alternans or VA was shorter in the ID group than in the ISO group (all P < .01), indicating that dantrolene treatment prevented the reduction in APD alternans and VA thresholds induced by chronic ISO stimulation (Figures 7 and 8). Because the PCL inducing VA differed, we did not compare the incidence and duration of VA among the 3 groups.

The APD alternans was induced in 3 groups. A, The MAPs were recorded in each group at pacing cycle length (PCL) of 200 ms; (B) MAP tracings during the APD alternans in 1 rat of each group. The numbers in all the panels represent APD90; (C) the median of maximal PCL induced APD alternans at 4 sites in each group. Dantrolene treatment prevented the reductions in the APD alternans threshold induced by chronic isoproterenol (ISO) stimulation at all recording sites. APD indicates action potential duration; MAP, monophasic action potential.

Ventricular arrhythmia (VA) was elicited by S1-S1 dynamic pacing in each group. A, The examples of VA in 3 groups; (B) the median of maximal pacing cycle length (PCL) induced VA at 4 sites in each group. Compared to the control value, chronic ISO stimulation significantly decreased the VA threshold at 4 recording sites. However, dantrolene treatment increased the VA threshold in the ISO + dantrolene (ID) group compared to the ISO group (all P < .01). *P < .01 versus control group; # P < .01 versus ISO group. ISO indicates isoproterenol.
Discussion
Main Findings
In the present study, we evaluated the effects of dantrolene treatment on ventricular electrophysiology and arrhythmogenesis in rats with chronic β-AR activation. We found that (1) treatment with dantrolene stabilized the cardiac electrophysiological properties of rats treated with chronic ISO stimulation, including shortened APD, flattened the APDR curves, and decreased spatial dispersions of APD90 and Smax; (2) dantrolene administration significantly preserved SERCA2a and RyR2 protein levels and decreased the expression of Cav1.2 and [Ca2+]i levels in chronically ISO-stimulated rats; and (3) dantrolene treatment significantly increased the APD alternans and VA thresholds in rats that received chronic ISO administration.
The Abnormal Changes in ECG After Chronic β-Adrenoceptor Stimulation
The widening of QRS complex and prolongation of QT interval have been reported in this study. Krandycheva et al 23 also showed the widened QRS complex and prolonged QT interval in the ISO-induced heart failure rat model. They demonstrated that the widened QRS complex induced by ISO stimulation was ascribed to the prolongation of epicardial activation duration. 23 In the current study, cardiac fibrosis induced by chronic ISO stimulation might attenuate the electrotonic interaction between ventricular myocytes and hence to increase epicardial activation duration. 23 Therefore, it is possible that cardiac fibrosis induced by ISO might lead to prolongation of QRS duration. Additionally, previous studies reported that ISO administration preferentially damage the subendocardial and midmyocardial layers of the heart ventricles, which could result in the transmural delay of activation and finally create a widened QRS complex. 23 –25 The prolonged QT interval induced by chronic ISO administration in the present study might be due to the prolongation of APD, which probably resulting from downregulation of outward K+ currents. 3
Dantrolene Attenuated the Abnormal Changes in Ventricular Repolarization Induced by Chronic ISO Stimulation
Soltysinska et al 3 reported that chronic ISO stimulation prolonged ventricular repolarization in guinea pig hearts. In this study, we also showed extended ventricular repolarization by chronic ISO stimulation, as evidenced by prolonged APD90 and QT intervals. In addition, similar to the previous studies, 26,27 we found that sustained β-AR stimulation increased the expression of Cav1.2 in ventricular tissue, which may be implicated in the prolonged ventricular repolarization caused by chronic ISO stimulation. However, dantrolene administration was found to attenuate a prolongation of ventricular repolarization induced by chronic ISO stimulation. Dantrolene can reportedly behave as verapamil, an L-type calcium channel blocking agent, to inhibit inward calcium current at adequate dose in cardiac cells. 28 –30 Although a patch-clamp experiment was not performed in this study, we observed that dantrolene treatment decreased the Cav1.2 protein levels in rats that received chronic ISO stimulation, which may result in the reduction of L-type Ca2+ current. Therefore, we hypothesize that the effects of dantrolene treatment on the L-type Ca2+ current may be implicated in the abbreviated ventricular repolarization observed in this study.
Furthermore, a posterior-to-anterior and apex-to-basal APD90 gradient was present in the ISO group, with the longest duration observed at LPA, resulting in an increased COV-APD90 and suggesting an increased spatial dispersion of APD90. In contrast, the COV-APD90 was significantly decreased by dantrolene treatment in the ID group. The increased spatial dispersion of APD90 has been suggested in previous studies to favor micro reentry and discordant APD alternans and VA. 21,31 Thus, we suggest that the reduction in spatial dispersion of APD90 may be associated with the antiarrhythmic effect of dantrolene treatment.
Dantrolene Prevented APD Alternans via Increased Expression of Key SR Calcium Cycling Proteins in Rats That Received Chronic ISO Stimulation
Previous studies suggested that APD alternans predisposed the ventricle to fibrillation, while a delayed APD alternans was associated with lower susceptibility to VA. 32,33 In this study, we observed that dantrolene treatment significantly increased the APD alternans threshold in rats chronically stimulated with ISO, indicating a potential role for dantrolene in reducing vulnerability to VA. Although the mechanisms of APD alternans have not been completely illuminated, the dysfunctional SR calcium handling hypothesis has been proposed to explain the cellular mechanism of APD alternans. 34 –36 It states that cellular alternans occurs when the heart rate exceeds the ability of myocytes to properly control calcium cycling. The RyR2 and SERCA2a are key SR calcium cycling proteins and critical factors in the generation of APD alternans. Previous studies demonstrated that dysfunctional SR calcium cycling caused by partial inhibition of RyR2 and SERCA2a function can cause [Ca2+]i cycling alternans, which directly implicated calcium cycling in the mechanism of APD alternans. 36 –38 Wan et al 34 showed that the expression levels of RyR2 and SERCA2a were lower in alternans-susceptible myocytes than in alternans-resistant myocytes. Moreover, overexpression of SERCA2a in the intact heart has been associated with significant resistance to APD alternans. 39 In the current study, dantrolene administration preserved the expression of RyR2 and SERCA2a, which were reduced by chronic ISO stimulation. Thus, we suggest that suppression of APD alternans by dantrolene treatment may be attributed to increased expression of key SR calcium cycling proteins in chronically ISO-stimulated rats.
Dantrolene Stabilized Ventricular APDR Properties to Suppress VA in Rats That Received Chronic ISO Stimulation
The electrical restitution hypothesis suggests that a steep APDR curve slope may precipitate oscillation in APD and electrophysiological instability during rapid pacing, thereby leading to the gradual development of conduction block, spiral wave break, and ventricular fibrillation. 40,41 According to the electrical restitution hypothesis, an increased APDR curve slope is associated with increased vulnerability to VA. Some antiarrhythmic drugs or other interventions that can flatten the APDR curve have been shown to prevent or suppress the occurrence of VA. 42 –44 In the present study, we showed that dantrolene treatment reduced the Smax of the APDR curves obtained in the chronically ISO-stimulated rat model, suggesting a potential antiarrhythmic effect for dantrolene. Intracellular Ca2+ dynamics have been proposed to play an important role in APDR. 45 During periods of fast heartbeat, increased [Ca2+]i accumulation can remarkably reduce the APD at a short DI, owing to [Ca2+]i accumulation induced inactivation of the L-type Ca2+ current or increases in the Ca2+-dependent outward K+ current, or both. 3,46 The balance of APD and previous phase DI can be shifted by [Ca2+]i accumulation and finally result in a steep APDR curve. In this study, dantrolene reduced the [Ca2+]i levels and preserved the expression of SERCA2a and RyR2 in chronically ISO-stimulated hearts, which may reduce the [Ca2+]i accumulation during rapid pacing. Therefore, we presume that the dantrolene-induced decrease in the Smax of APDR curves in chronically ISO-stimulated hearts was partly due to improved [Ca2+]i cycling. However, the detailed mechanism by which dantrolene flattens the APDR curve in chronically ISO-stimulated rats requires further investigation.
Additionally, it has been demonstrated that the APDR curve obtained at a single site may not represent the kinetics of APDR restitution in the whole heart. 47 Nash et al 48 showed that the spiral wave break could be caused by the increased spatial differences in Smax. In addition, Lu et al 49 reported that vagal nerve stimulation precipitated atrial fibrillation associated with an increased spatial difference in Smax. These studies demonstrated that the increased spatial difference in Smax also played an important role in arrhythmogenesis. In the current study, the spatial dispersion of Smax was evaluated by calculating the COV-Smax. We determined that dantrolene treatment decreased the COV-Smax in the chronically ISO-stimulated rat model, also suggesting that dantrolene exerts a protective effect during VA.
Dantrolene Improved the Cardiac Remodeling Induced by ISO Chronic Stimulation
Sustained ISO stimulation causes the excessive production of free radicals, cytosolic Ca2+ overload, and severe stress in the myocardial tissue. As shown in previous studies, 50,51 we also observed that chronic ISO stimulation induced cardiac hypertrophy accompanied by enhanced fibrosis among the interstitial cardiac tissue. Both abnormal Ca2+ handling and cardiac oxidative stress have been proposed to induce changes in cardiac structure following chronic ISO stimulation. 50,52 Shibata et al 52 demonstrated that the upregulation of SERCA2a protein levels, which improved the Ca2+ handling dysfunction, inhibited the development of cardiac remodeling in chronically ISO-infused rats. Thus, we suggest that the enhanced levels of SERCA2a and RyR2 induced by dantrolene were implicated in ameliorating the cardiac remodeling induced by chronic ISO stimulation. Additionally, Buyukokuroglu et al 53 reported that dantrolene has antioxidant properties in a myocardial infarction model. However, whether the antioxidant effects of dantrolene prevent the development of cardiac remodeling requires investigation.
Dantrolene Alleviated the [Ca2+]i Overload Induced by Chronic ISO Stimulation
In this study, dantrolene treatment markedly decreased the elevated [Ca2+]i induced by chronic ISO stimulation, suggested that dantrolene alleviated the [Ca2+]i overload induced by chronic ISO stimulation. Many studies reported that ISO treatment could increase diastolic [Ca2+]i and decrease the SR Ca2+ content by reducing the SERCA2a activity and enhancing the Ca2+ leak from SR during diastole, these effects may finally result in [Ca2+]i overload. 4,6,7 Bers 54 demonstrated that SERCA2a pump, which could remove 92% of the activator Ca2+ during diastole, plays a pivotal role in regulating the [Ca2+]i cycling in rat cardiac cells. In this study, dantrolene treatment prevented the reduction in SERCA2a protein expression caused by chronic ISO administration, which may be the important action of dantrolene to alleviate the [Ca2+]i overload induced by chronic ISO stimulation in this study. Additionally, recent evidences have suggested that as a stabilizer of RyR function, dantrolene could inhibited diastolic Ca2+ leak from SR to restore the proper [Ca2+]i handling in heart failure model. 11,12 Thus, we suggest that dantrolene alleviated [Ca2+]i overload may also via reducing the diastolic Ca2+ leak from SR in the present study.
Clinical Implications
Progression of chronic heart failure and myocardial infarction is accompanied by electrical remodeling, which could promote the development of potentially lethal VA and sudden cardiac death. It has been shown that catecholamine levels are chronically elevated in patients with chronic heart failure and myocardial infarction. Increased plasma catecholamine levels can activate the β-AR and hence affect cardiac function. Both our previous report and other studies have demonstrated that chronic activation of the β-AR can distinctly alter cardiac electrophysiological properties and facilitate VA. 3,4 Thus, the increased catecholamine levels might well play a pathogenic role in electrical remodeling during chronic heart failure and myocardial infarction. Meanwhile, certain therapeutic interventions that attenuate the electrical remodeling caused by chronic β-AR activation may be beneficial for reducing the incidence of VA and sudden death in patients with chronic heart failure and myocardial infarction. In this study, we showed that dantrolene treatment stabilizes the ventricular electrophysiological properties to reduce the vulnerability to VA in rats with chronic β-AR activation, indicating that dantrolene may be a promising antiarrhythmic drug for preventing VA in these cardiovascular diseases.
Study Limitations
There are several limitations to the present study. First, we determined only electrophysiological characteristics at epicardial sites, but the electrophysiological properties between the epicardium to endocardium are different, and whether dantrolene treatment affects endocardial ventricular electrophysiological properties in the chronically ISO-stimulated model remains unknown. Second, although previous studies have demonstrated that dantrolene can inhibit the L-type Ca2+ current at adequate dose, whether the dose of dantrolene used in this study is sufficient to directly reduce the L-type Ca2+ current is still unclear, further research is needed to assess the changes in the L-type Ca2+ current by the dose of dantrolene used in this study. Third, we determined only the changes in RyR2 and SERCA2a protein expression and [Ca2+]i levels to indirectly assess the SR calcium cycling, but the calcium transient recording, which can directly assess the SR calcium cycling, was not assessed in this study. Fourth, it is well known that ventricular repolarization characteristics of rat ventricular cells differ from that of human ventricular cells. Unlike human hearts, rat hearts exhibit a short duration of ventricular repolarization and a triangular shape of action potential and lack a well-defined plateau phase during repolarization, which is attributed to the differently predominated outward K+ current contributing to ventricular repolarization. The major ionic current greatly mediating the ventricular repolarization of rat hearts is transient outward K+ current (Ito), whereas the fast and the slow components of delayed rectifier K+ currents (IKr and IKs) play an important role in ventricular repolarization of human hearts. 3,55,56 Therefore, the electrophysiological findings obtained from rat hearts in this study can’t directly be extrapolated to human hearts.
Conclusions
The present study demonstrates the effects of dantrolene treatment on ventricular electrophysiological properties and arrhythmogenesis in rats with chronic β-AR activation. These data suggest that dantrolene treatment stabilizes ventricular electrophysiological characteristics and increases the expression of key SR calcium cycling proteins to reduce vulnerability to VA in rats with chronic β-AR activation.
Footnotes
Author Contributions
T. Liu contributed to conception and design, analysis, drafted the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. S.-B. Shi contributed to conception, acquisition and interpretation, drafted the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. M. Qin contributed to design, acquisition and interpretation, critically revised the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. C. X. Huang contributed to conception and design, acquisition, analysis, and interpretation, critically revised the article, gave final approval, and agreed to be accountable for all aspects of work ensuring integrity and accuracy. T. Liu and S.-B. Shi contributed equally to this work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Key Basic Research Development Program of China (The “973” Program; no. 2012CB518604), Natural Science Foundation of Hubei Province China (no. 2013CFA117), and the Fundamental Research Funds for the Central Universities of China (no. 2014302020201). This study also supported by the China Scholarship Council.