
Abstract
The discovery of the water specific channel, aquaporin, and abundant expression of its isoform, aquaporin-4 (AQP-4), on astrocyte endfeet brought about significant advancements in the understanding of brain fluid dynamics. The brain is protected by barriers preventing free access of systemic fluid. The same barrier system, however, also isolates brain interstitial fluid from the hydro-dynamic effect of the systemic circulation. The systolic force of the heart, an essential factor for proper systemic interstitial fluid circulation, cannot be propagated to the interstitial fluid compartment of the brain. Without a proper alternative mechanism, brain interstitial fluid would stay stagnant. Water influx into the peri-capillary Virchow-Robin space (VRS) through the astrocyte AQP-4 system compensates for this hydrodynamic shortage essential for interstitial flow, introducing the condition virtually identical to systemic circulation, which by virtue of its fenestrated capillaries creates appropriate interstitial fluid motion. Interstitial flow in peri-arterial VRS constitutes an essential part of the clearance system for β-amyloid, whereas interstitial flow in peri-venous VRS creates bulk interstitial fluid flow, which, together with the choroid plexus, creates the necessary ventricular cerebrospinal fluid (CSF) volume for proper CSF circulation.
Introduction
Advances in modern technology have transformed biomedical investigation. Molecular biological techniques have made it possible to analyze target protein functionality, while non-invasive molecular imaging made possible in vivo quantitative dynamic analysis of target physiological systems. As a result, widely held concepts have been challenged and old, often overlooked ideas have undergone dramatic revival (Nakada 2014; Orešković and Klarica 2010). The field of biological water dynamics has greatly advanced owing to the discovery of the water specific channel, aquaporin (AQP) (Benga and others 1986; Denker and others 1988). Paradoxically, however, this new discovery has also introduced significant confusion among investigators, resulting in misinterpretation of the findings. Even leading authorities in the field have quoted misleading information, such as AQP’s involvement in water movement across the barriers (Papadopoulos and Verkman 2013) or bulk flow in the paravascular pathway of gray matter (Iliff and others 2012). These misconceptions probably reflect the highly complex nature of brain water dynamics, which require a highly multidisciplinary approach. This review provides a succinct summary of the historical progression and modern understanding of barrier systems and interstitial fluid motion in an attempt to present a logical, systematic model of water dynamics in the brain.
The Brain Barrier
Main Barrier Systems
The interstitial fluid of the brain is effectively isolated from the systemic circulation. This fact was originally discovered by Paul Ehrlich, Nobel Laureate, in 1885. He observed that intraperitoneal dye injection rapidly stains all organs but the brain. Max Lewandowsky subsequently hypothesized the existence of a physical barrier at the level of cerebral blood vessels underlying this phenomenon in 1900 and coined the term blood-brain barrier (BBB) (Lippoldt and others 2000). Modern science has since delineated the architectural details of this protective system of the brain, which we refer to here as the Brain Barrier (Brøchner and others 2015; Günzel and Yu 2013; Liddelow and others 2011; Rosenthal and others 2010; Saunders and others 2008; Weiss and others 2009; Yasuda and others 2013).
There are three main barrier systems comprising the Brain Barrier, that is, BBB, blood–cerebrospinal fluid (CSF) barrier (BCSFB), and outer brain barrier (OBB). Circumventricular organs (CVOs) located around the third and fourth ventricles are sealed off by special ependymal cells such as tanycyte
The main player in barrier function is the tight junction, a structure virtually impermeable to fluids (Günzel and Yu 2013; Lippoldt and others 2000; Rosenthal and others 2010; Weiss and others 2009; Yasuda and others 2013). Although it has been shown ependymal cells have a so-called strap junction made initially by neuroepithelial cells and later by radial glial cells during early brain development that restricts free diffusion through the ependyma, in adult brain, the majority of ependymal cells have only gap junctions without diffusional restraint (Johanson and others 2011; Spector and Johanson 2014; Whish and others 2015).
The primary structure responsible for the BBB is the tight endothelium of brain capillaries (Weiss and others 2009). Common capillaries have a leaky endothelium due to the presence of fenestrations. The water channel aquaporin-1 (AQP-1), abundantly expressed in common capillaries, is actively suppressed in brain capillaries. As a result, water dynamics in the intracapillary space and interstitial fluid of the space surrounding brain capillaries are effectively isolated from each other (Fig. 1).
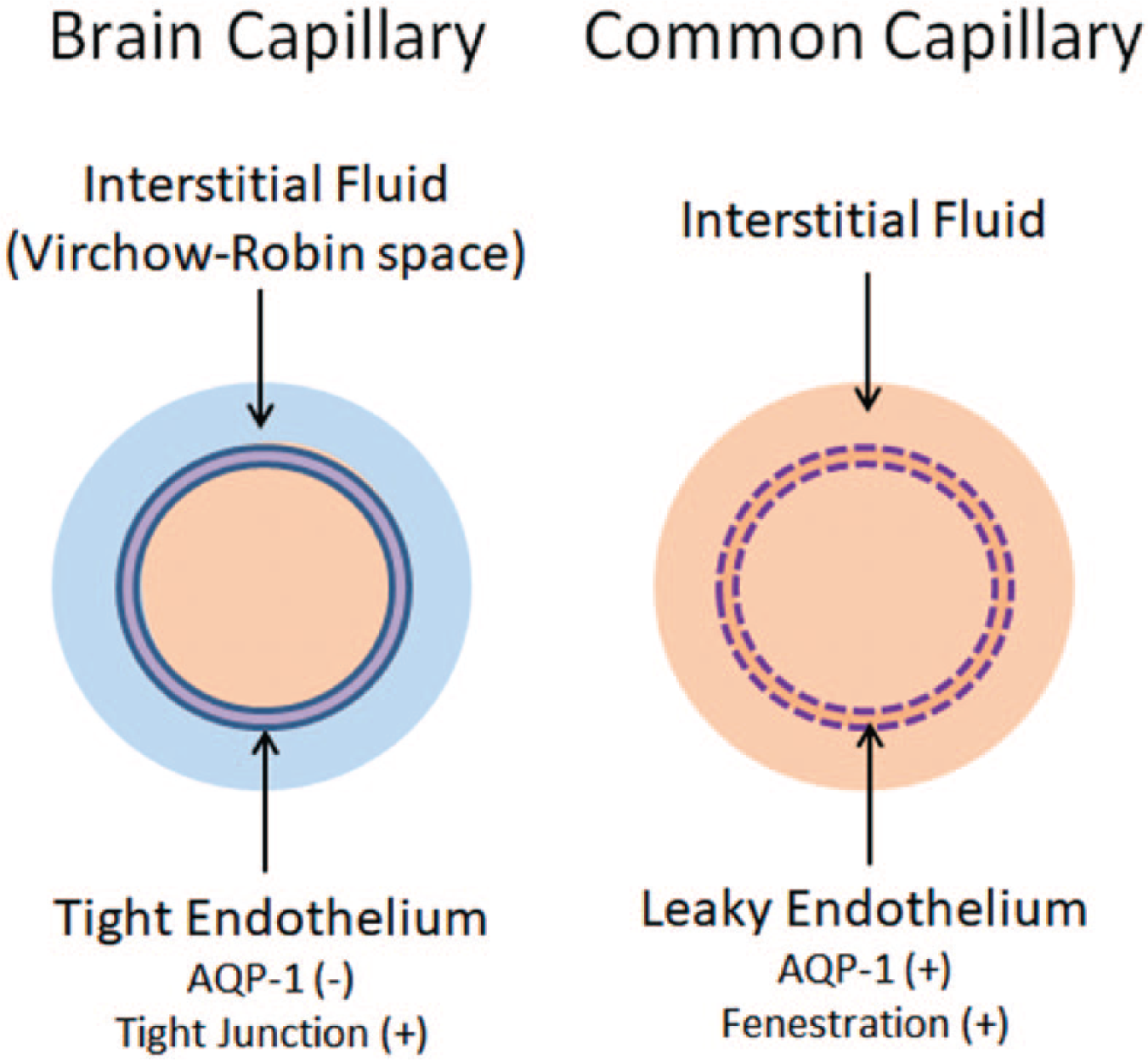
Blood-brain barrier. Common capillaries have a leaky endothelium due to the presence of fenestrations. The water channel aquaporin-1 (AQP-1) is also abundantly expressed. Accordingly, fluid from the intracapillary space moves freely into the interstitial fluid space (right). In contrast, brain capillaries lack fenestrations and have tight junctions. Expression of AQP-1 is actively suppressed. This makes the endothelium virtually non-permeable to water, constituting the highly selective blood-brain barrier (BBB) for substrates to enter the interstitial fluid space. Blue fluid indicates the fluid inside the Brain Barrier. Modified from Kitaura and others (2009).
The primary structure responsible for the BCSFB is the choroid plexus epithelium (Lippoldt and others 2000). Like common capillaries, capillaries of the choroid plexus are also fenestrated. The apical ends of choroid plexus epithelium are, however, tightly sealed by tight junctions, preventing free diffusion into the CSF through the choroid plexus (Fig. 2).
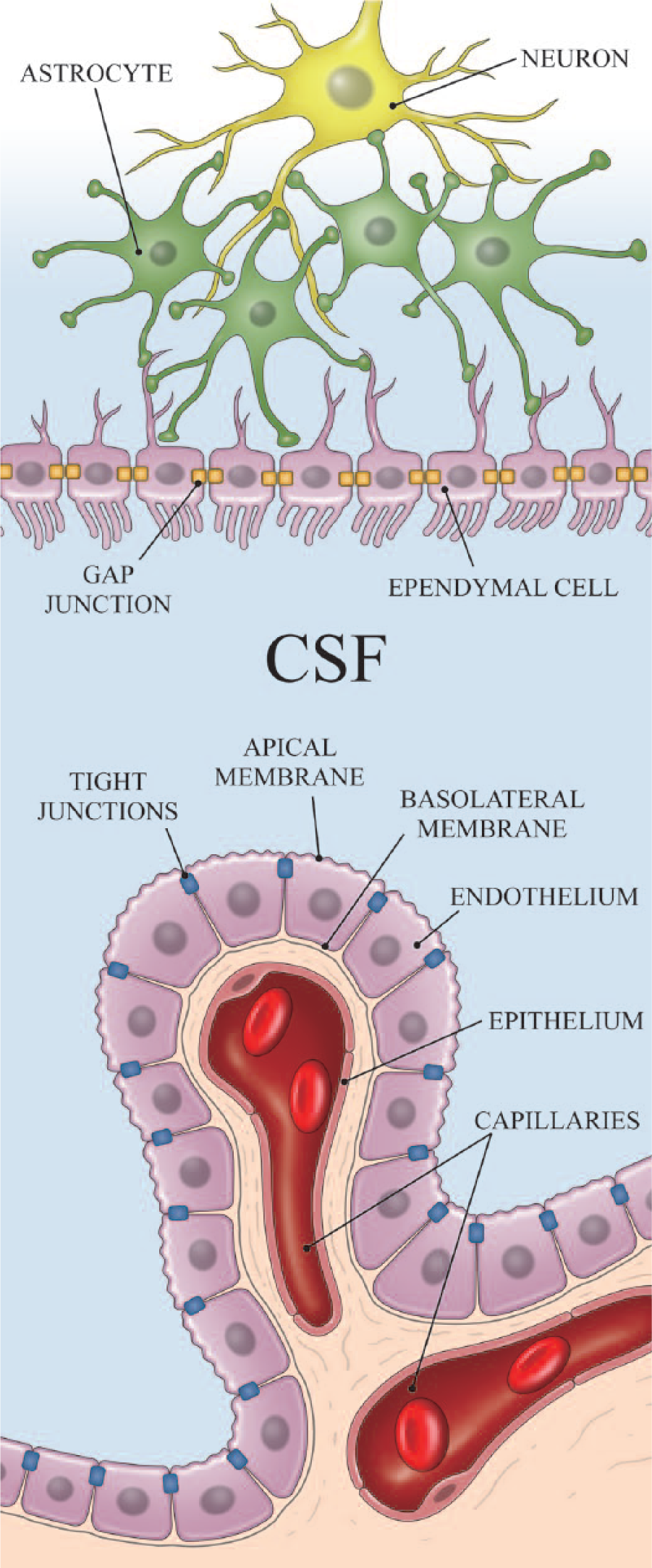
blood–cerebrospinal fluid (CSF) barrier. The choroid plexus has fenestrated capillaries. Nevertheless, the apical side of choroid plexus epithelium is tightly connected with tight junctions, making the blood-CSF barrier (BCSFB). Ependymal cells are joined with gap junctions, and CSF communicates relatively freely with interstitial fluid. Blue fluid indicates the fluid inside the Brain Barrier.
The primary structure responsible for the OBB is the arachnoid barrier (AB) cells (Brøchner and others 2015). The brain meninges consist of an outermost dura matter and the two inner layers (leptomeninges), comprising the arachnoid mater and the innermost pia mater. Arachnoid matter contains epithelial cell lines which are joined by tight junctions, effectively preventing interstitial fluid diffusion from the dura and subdural area, structures which have fenestrated capillaries. By contrast, arteries and veins in the subarachnoid space are well sealed by the tunica and capillaries inside the AB cells having tight endothelium (Fig. 3).
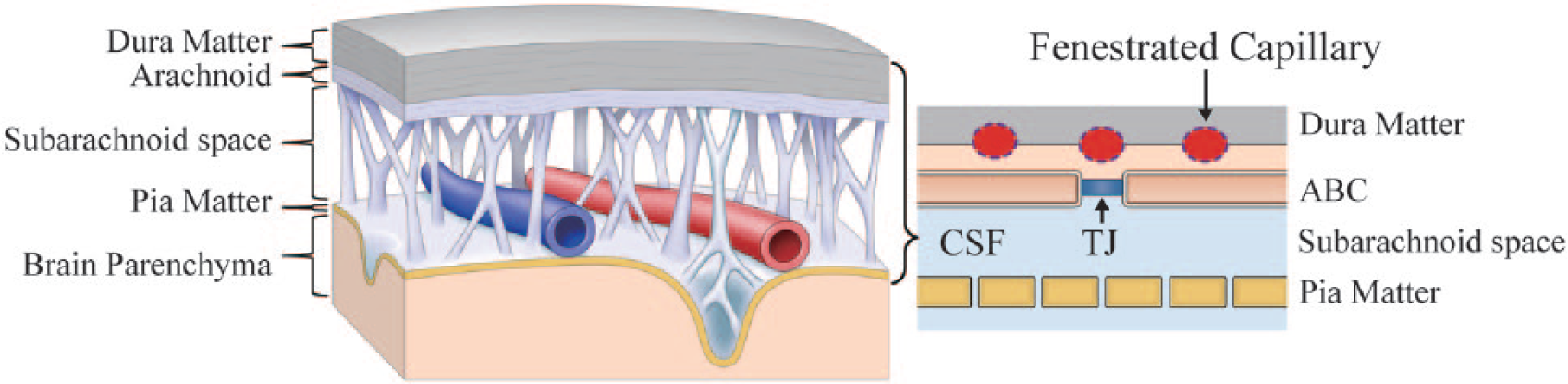
Outer brain barrier. Capillaries within the dura matter and subdural space are fenestrated. Arachnoid barrier cells are tightly connected with tight junctions, making the outer brain barrier (OBB). Arteries and veins within the subarachnoid space are well sealed by the tunica, and capillaries inside the arachnoid barrier cells (ABCs) have tight endothelium. Cerebrospinal fluid (CSF) of the subarachnoid space communicates relatively freely with interstitial fluid across the pia matter. Blue fluid indicates the fluid inside the Brain Barrier. TJ, tight junction.
Tight Junction
The tight junctions responsible for the Brain Barrier are those of endothelial cells of brain capillaries and epithelial cells of choroid plexus and arachnoid matter. Tight junctions of brain endothelial cells are structurally similar to epithelial tight junctions (Günzel and Yu 2013). The major transmembrane proteins in tight junctions include occludins and claudins. Some accessory proteins specific to brain endothelium, such as cingulin, AF-6 and 7H6, have been identified. Epithelial tight junctions, including that of choroid plexus and arachnoid epithelial cells, mostly contain claudin-1, -2, and -11. For choroid plexus, claudin-5 is also expressed (Günzel and Yu 2013; Lippoldt and others 2000). Brain capillary endothelial tight junctions primarily express claudin-3 and 5. Thus far, only claudin-2 has been shown to be permeable to water (Rosenthal and others 2010). The heterogeneity of claudins expression in tight junctions suggests differential functionality. While BCSFB and OBB may have some permeability to water due to the presence of claudin-2, the BBB is essentially impermeable to water.
Aquaporin (AQP)
Over a span of just a few short years, the consensus regarding physiological water trafficking has shifted from being dependent on non-specific permeability across the plasma membrane to efficient water movement via specialized channels. This shift in perspective began in earnest during the late-1980s with the pioneering work of the Gheorghe Benga and Peter Agre groups on the membrane protein now recognized as AQP-1 (Benga and others 1986; Denker and others 1988). The subsequent explosion of AQP-related research demonstrated the existence of these proteins in virtually every organism.
Thus far, seven AQP isoforms have been identified in the mammalian central nervous system (CNS) by reverse transcription polymerase chain reaction (RT-PCR). However, only three isoforms, namely, AQP-1, AQP-4, and AQP-9, have been identified in the mammalian CNS in vivo (Badaut and others 2003). AQP-4 represents the most abundant isoform, the distribution of which is uniquely polarized to endfeet of astrocytes at the subpial glia limitans externa (GLE) and peri-capillary space, and glial side of ependymal cell membranes (Huber and others 2012; Rash and others 1998). AQP-1, abundantly expressed on the endothelium of common capillaries, is absent in the endothelium of brain capillaries (Nakada 2014). AQP-1, however, is expressed on membranes of the apical (CSF) side of choroid plexus epithelium (Fig. 4). AQP-9 is only scarcely expressed in the CNS and is believed to have a highly specialized role in local structures, as does AQP-4 in Muller cells of the retina or Hensen and inner sulcus cells of the cochlea (Huber and others 2012).
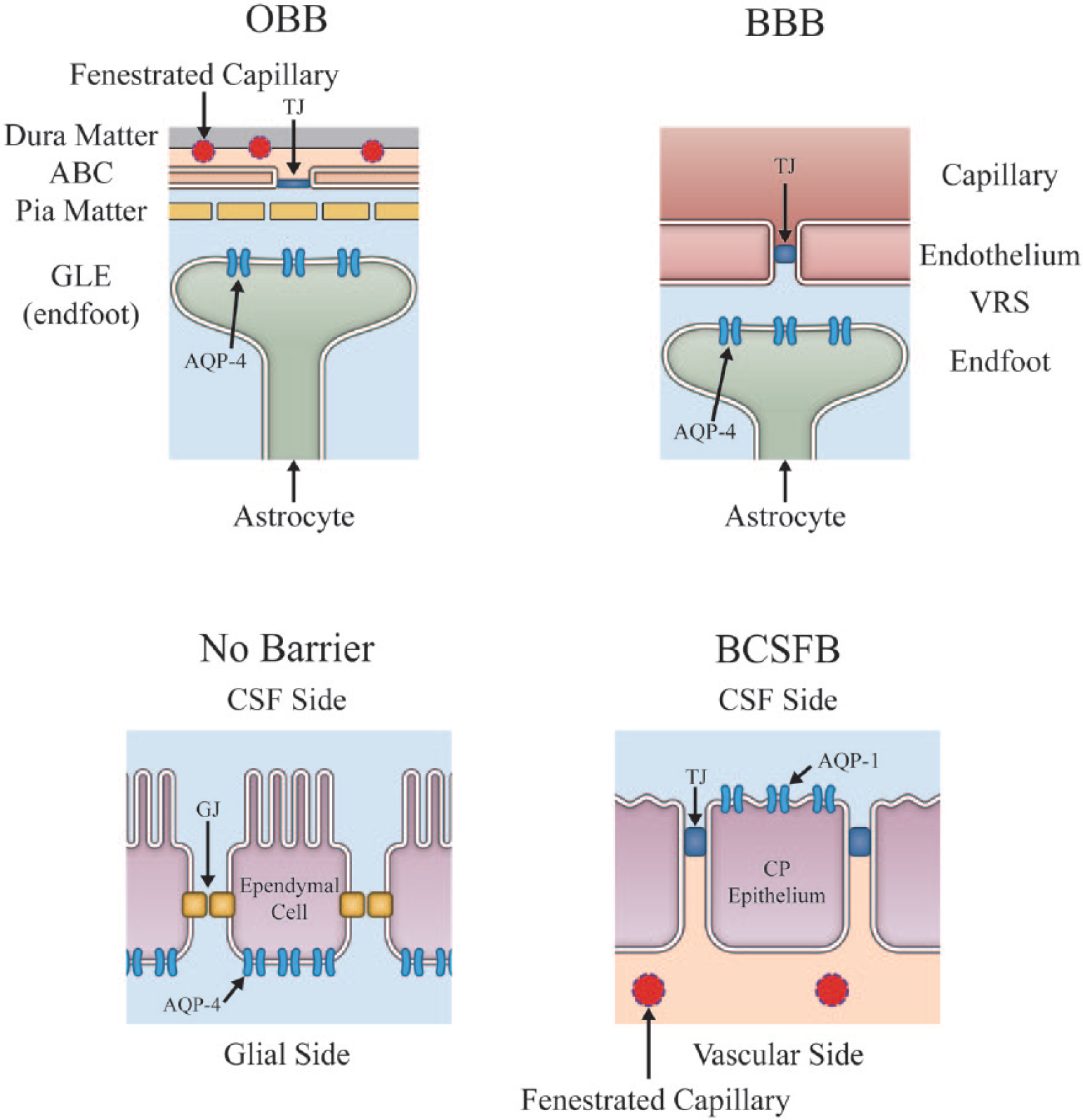
Main locations for aquaporin (AQP). Expression of AQP is uniquely polarized. The primary distribution of AQP-4 is confined to astrocyte endfeet at the subpial glia limitans externa (GLE) and peri-capillary space, and glial side of ependymal cell membrane. AQP-1 is found on the apical (cerebrospinal fluid [CSF]) side of choroid plexus epithelium. Virtually all AQP-4 in the brain are expressed on membrane localized inside the Brain Barrier, confirming that AQP-4 does not play a direct role for water communication between the outside and inside of the Brain Barrier. Blue fluid indicates fluid inside the Brain Barrier. VRS, Virchow-Robin space; TJ, tight junction; GJ, gap junction, ABC, arachnoid barrier cell.
Virtually all brain AQP-4 is expressed on membrane located inside the Brain Barrier. Therefore, AQP-4 cannot be directly involved in water movement across the Brain Barrier. This fact is often overlooked even by members of the leading AQP research teams who have at times made enthusiastic claims for this role of aquaporin in prestigious journals (Papadopoulos and Verkman 2013). It is clear that the abundance of AQP-4 within the Brain Barrier should be interpreted to mean that AQP-4 is involved in active dynamics of water movement inside the Brain Barrier (Nakada 2014; Nakada and others 2017).
The presence of AQP-1 on the apical membrane of choroid plexus epithelium suggests a role for AQP-1 in CSF production. Although net water movement through the choroid plexus epithelium is believed to be favor production of CSF and water efflux from the intracellular space of choroid plexus epithelium to CSF is likely facilitated by AQP-1, the molecular mechanism of water influx into the intracellular space of the choroid plexus epithelium through the basal (vascular side) membrane is unclear (Boassa and Yool 2005; Brown and others 2004; Praetorius and Nielsen 2006). Some studies indicated that there might be some AQP-1 expressed on the basal (vascular side) membrane as well. The general consensus among investigators, however, is that there is no specific water channel on the basal side of the choroid plexus epithelium. Therefore, it is reasonable to conclude that, similar to AQP-4 expression on the glial side of ependymal membrane, AQP-1 expression on choroid plexus epithelium has a rather passive role in maintaining CSF homeostasis, rather than having an active role in water transfer across the Brain Barrier (Lippoldt and others 2000; Rosenthal and others 2010). Indeed, in vivo non-invasive water dynamic studies on AQP-1 knock-out mice unequivocally demonstrated that there is no significant differences in water influx into CSF from the systemic circulation compared to the wild animals (Igarashi and others 2014b).
Water Movement across the Brain Barrier
Interstitial fluid of the brain is effectively isolated from the systemic circulation by the Brain Barrier consisting of the BBB, BCSFB, and OBB. There is no specific passage for water to move across the Brain Barrier. There are only two routes through which water can move across the Brain Barrier, namely (1) non-specific permeability across the plasma membrane and (2) claudin-2 at epithelial tight junction. Plasma membrane does have some water permeability, the degree of which is dependent on its lipid composition (Mathai and others 2008). Claudin-2, which has some water permeability, is shown to exist at tight junctions of epithelium including choroid plexus epithelium and AB cells. AQP-4, which is abundantly expressed inside the Brain Barrier, should play the specific role of maintaining proper water dynamics inside the Brain Barrier independent of the systemic circulation (Nakada 2014; Nakada and others 2017).
Fluid Dynamics inside the Brain Barrier
Interstitial Flow
Bulk flow of interstitial fluid in the brain has been the subject of investigation by many neuroscientists over many decades. Initially, the nature of the investigations was primarily in the context of identifying the source of CSF (Cushing 1914). The report by Weed in 1914 set the clear tone that, in addition to choroid plexus, there is another source of CSF based on interstitial fluid of the brain (Weed 1914). Bulk flow of such interstitial fluid is believed to be present in the peri-vascular space.
Fluid-filled canals surrounding perforating arteries, capillaries, and veins in the brain parenchyma were recognized early in modern medicine and were referred to as the Virchow-Robin space (VRS) after the first two scientists who described the structure in detail, namely, Rudolph Virchow in 1851 (Virchow 1851) and Charles Philippe Robin in 1859 (Robin 1859), respectively. Accordingly, VRS was postulated to be the responsible structure for bulk flow of interstitial fluid.
Although the concept has generally been accepted by the scientific community, it remained in the periphery of CSF physiology. Perhaps the simplicity of the “Choroid Plexus Hypothesis,” which states that CSF is made by the choroid plexus and circulates through the ventricles and subarachnoid spaces, made it appealing as the standard concept until recent serious challenges were made by various investigators (Brown and others 2004; Igarashi and others 2014b; Orešković and Klarica 2010; Orešković and others 2017).
Several key features regarding interstitial flow have become apparent. There are three basic general agreements regarding the physiology of interstitial flow, namely (1) interstitial fluid motion is primarily dependent on diffusional processes in gray matter, (2) white matter is the primary site for bulk flow in the VRS, and (3) interstitial flow is another source of CSF (Abbott 2004; Cserr and Ostrach 1974; Rosenberg and others 1980).
Recently, interstitial flow has undergone dramatic revival with regard to β-amyloid clearance (Iliff and others 2012; Suzuki and others 2015; Tarasoff-Conway and others 2015; Weller 1998). Since the brain lacks a conventional lymphatic system, the concept that interstitial flow in the VRS plays a role similar to systemic lymphatics for the brain was put forward almost immediately after discovery of the VRS. However, similar to interstitial flow itself, the idea was buried long ago. The discovery that β-amyloid clearance is dependent on interstitial flow in the VRS provided substantial excitement in the scientific community, and the term glymphatic, denoting glial lymphatics, was coined (Iliff and others 2012).
Interstitial Fluid Dynamics and Astrocyte AQP-4 System
From a hydrodynamic standpoint, the tight endothelium of brain capillaries responsible for the BBB creates a serious structural problem for maintaining proper interstitial fluid dynamics. In the systemic circulation where common, leaky capillaries exist, hydrostatic pressure generated by the systolic force of the heart effectively extrudes water out of capillaries without much resistance into the interstitial fluid space and helps create interstitial fluid movement necessary for interstitial circulation. The hydrostatic pressure created by the systolic force of the heart is also sufficient to return excess interstitial fluid to the systemic circulation through lymphatic capillaries under physiological conditions (Levick 2010; Margaris and Black 2012). Because of the tight junctions of brain capillary endothelium, the interstitial fluid system of the brain cannot benefit from the hydrodynamic force of the systolic pulse of the heart, and interstitial fluid may become “stagnant” without a proper hydrodynamic alternative.
The lack of fenestrated capillaries results in a lack of fluid influx into the peri-capillary interstitial space (Nakada and others 2017). This implies that, in order to have proper interstitial fluid dynamics inside the Brain Barrier akin to systemic conditions, the brain must have an alternate functional system that provides appropriate fluid influx into the peri-capillary interstitial space. Accumulating evidence now indicates that the astrocyte AQP-4 system provides essential fluid influx into the peri-capillary interstitial space in brain (Fig. 5) (Amiry-Moghaddam and others 2003; Kitaura and others 2009; Haj-Yasein and others 2012; Igarashi and others 2013; Nakada and others 2017).
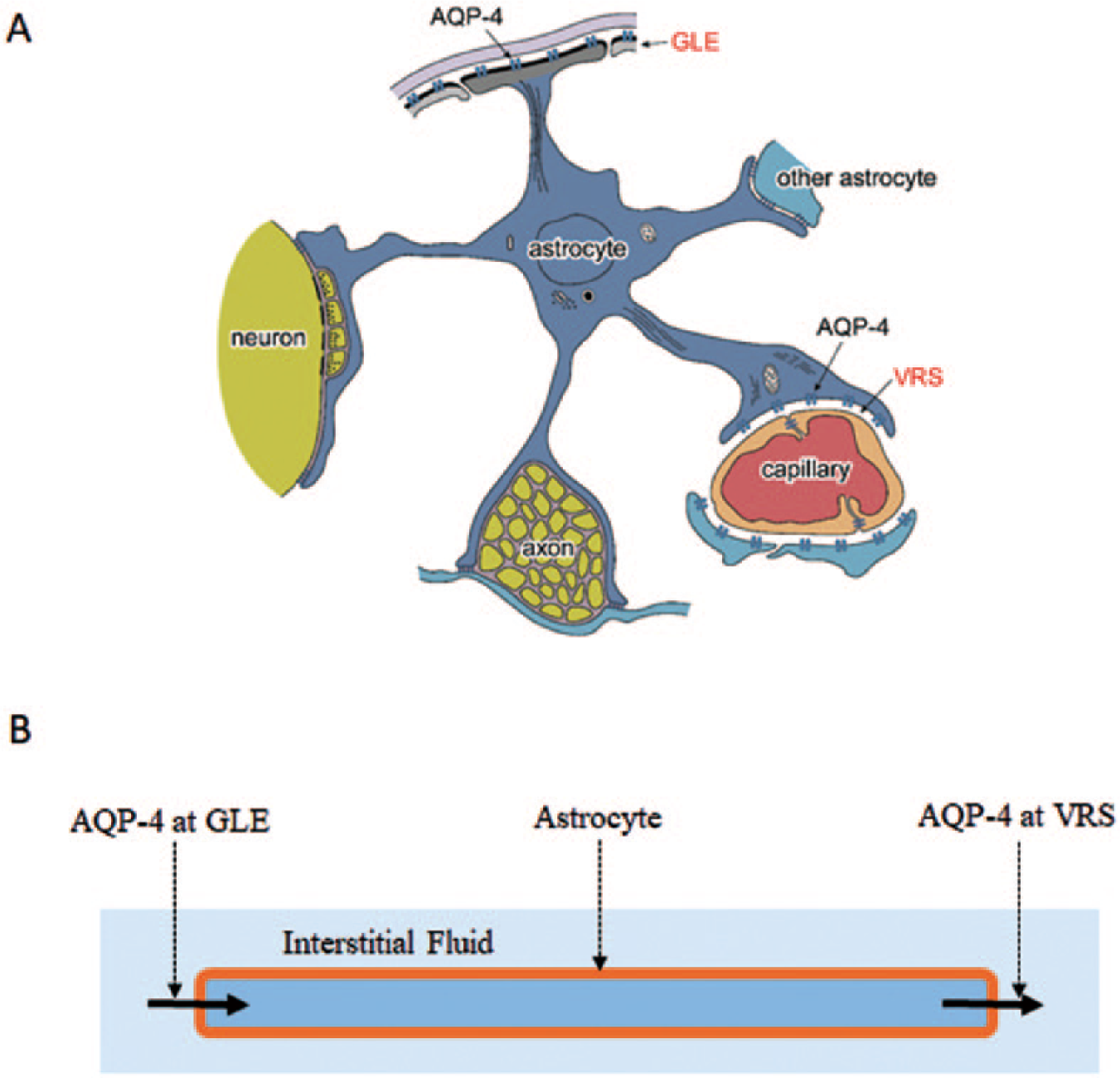
Schematic presentation of astrocyte aquaporin-4 (AQP-4) system. The AQP-4 system provides water influx into the peri-capillary Virchow-Robin space (VRS). Necessary water enters astrocytes through AQP-4 at the glia limitans externa (GLE). This system promotes appropriate interstitial fluid circulation, including bulk flow through the VRS (interstitial flow). Modified from Suzuki and others (2017).
AQP-4 of astrocyte endfeet at the peri-capillary space control peri-capillary fluid dynamics by providing water influx into the space. Since AQP-4 connects the intracellular and extracellular (interstitial) spaces of astrocytes, water efflux from astrocytes into the interstitial space must be balanced by an equivalent amount of water influx in order to maintain water equilibrium. AQP-4 localized to endfeet at the GLE is believed to play the role of ensuring proper water influx into the intracellular space of astrocytes (Fig. 5). A 7-T magnetic resonance imaging (MRI) microscopic investigation of the GLE in humans unequivocally supported this concept (Suzuki and others 2017). Furthermore, in vivo water dynamic studies have shown that this interstitial fluid circulation can be effectively facilitated by the AQP-4 facilitator, TGN-073 (Huber and others 2018).
The astrocyte AQP-4 system compensates for the critical shortcoming of the tight endothelium necessary for forming the BBB, and functions as circulator of interstitial fluid within the Brain Barrier. Its main role is to provide appropriate water influx into the peri-capillary interstitial space. The system not only ensures proper circulation of interstitial fluid but also provides necessary hydrodynamic conditions for interstitial flow (Figs. 6 and 7) as detailed below.
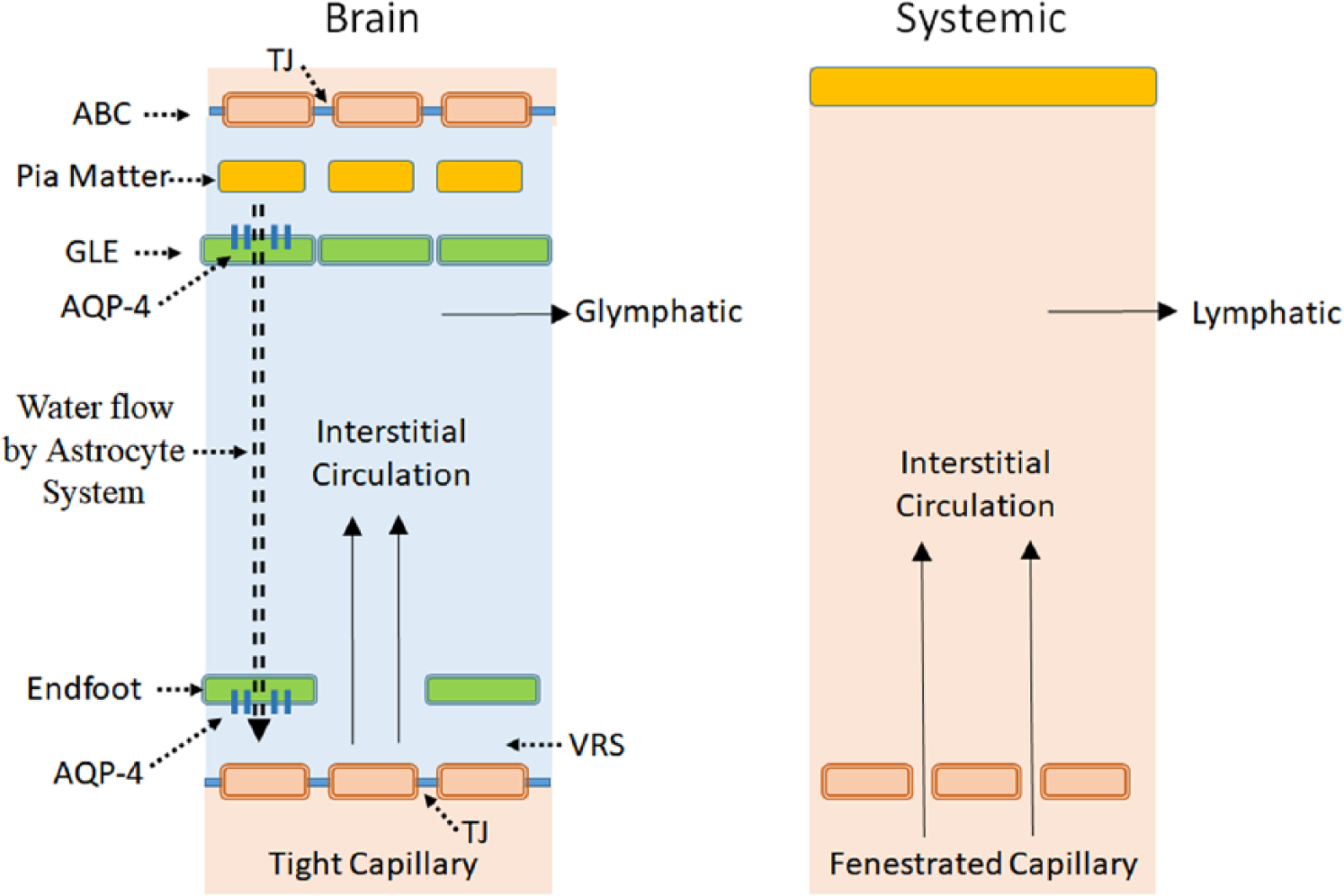
Schematic presentation of interstitial fluid dynamics. In order to have a hydrodynamic condition similar to that of the systemic environment with its fenestrated capillaries, water influx has to be provided into the peri-capillary VRS. The astrocyte aquaporin-4 (AQP-4) system effects this by removing water out of the subpial space and infusing water into peri-capillary VRS (double dotted line). The system creates the proper hydrodynamic environment for interstitial circulation as well as glymphatics akin to systemic lymphatics. Blue fluid indicates the fluid inside the Brain Barrier. ABC, arachnoid barrier cell; GLE, glia limitans externa; VRS, Virchow-Robin space; TJ, tight junction.
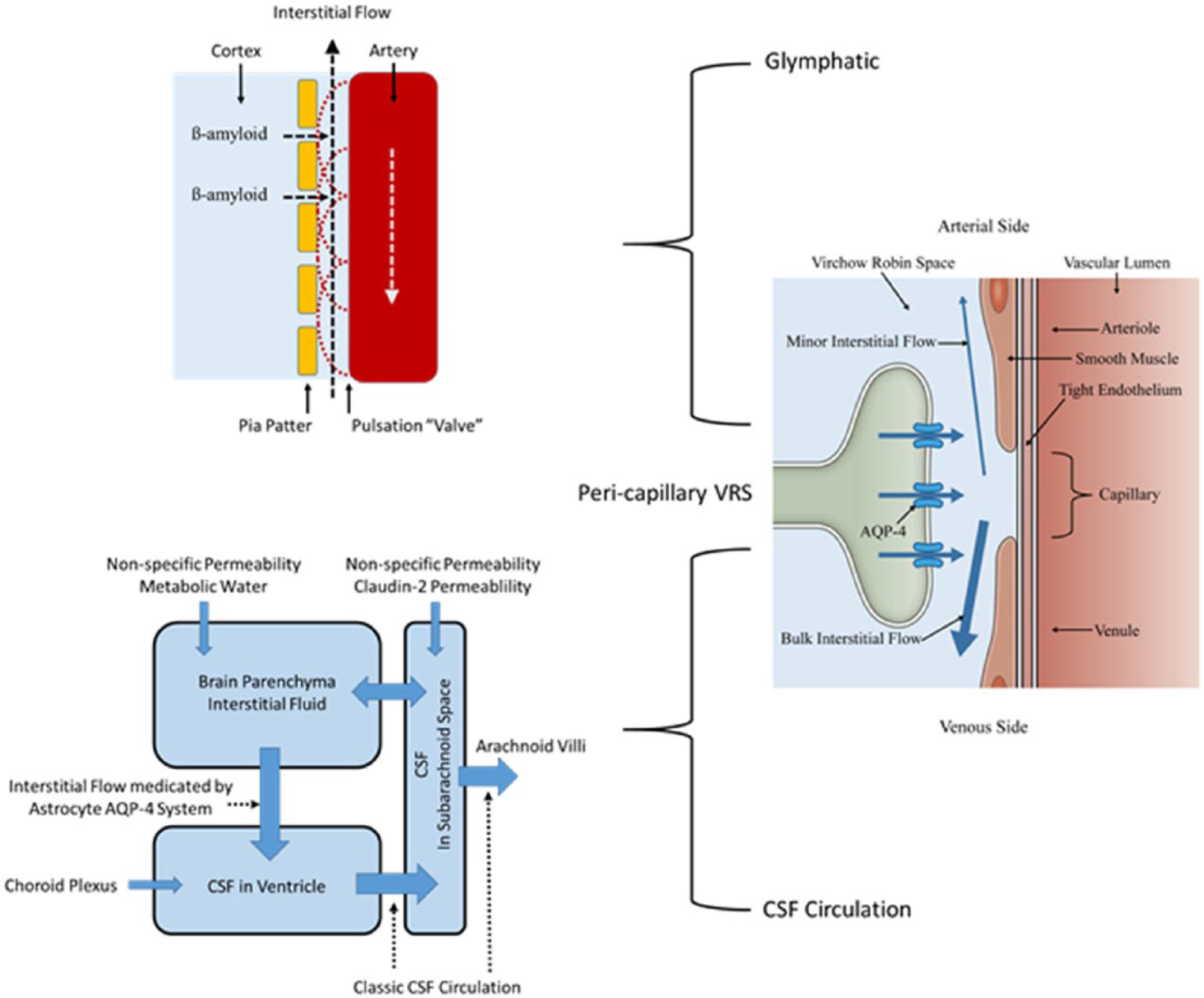
Unified view. The only driving force for the entire brain fluid dynamics system is the astrocyte aquaporin-4 (AQP-4) system. Water influx through AQP-4 at the peri-capillary VRS produces interstitial fluid flow within the VRS in both arterial and venous directions, in addition to providing proper interstitial circulation. Interstitial flow in the peri-arterial VRS is retrograde and is sufficient to create bulk flow. Nevertheless, together with intermittent closure of the flow passage by arterial pulsation, this minor flow helps prevent backflow of interstitial fluid to clear β-amyloid into the subarachnoid space (glymphatic). In contrast, interstitial flow in the peri-venous VRS is orthograde and constitutes bulk flow that has long been known to brain scientists. This bulk interstitial flow in the peri-venous VRS together with CSF produced by choroid plexus continuously provide water influx into the ventricles, creating CSF circulation. Water can enter inside of the Brain Barrier by non-specific water permeability through the plasma membrane. Metabolic water is another source of parenchymal interstitial fluid. Claudin-2 of the tight junctions in the outer brain barrier (OBB) may provide an additional water source to the subarachnoid space.
Glymphatic Flow
The basic function of the lymphatic system is drainage of cellular debris subjected to molecular scrutiny before returning to the venous system. Therefore, systemic lymphatics have two principal functions, namely, clearance of debris from tissues and returning excess water from tissues to the venous system. In this schema, CSF circulation should play the role of the lymphatic system equivalent as a whole and arachnoid villi plays the role of the thoracic duct. Accordingly, the concept of glymphatics as the lymphatic equivalent of the brain, should also be applied to the functional architecture for clearing debris such as β-amyloid from brain parenchyma into the CSF. As discussed below, brain glymphatics are not synonymous with interstitial fluid motion and bulk flow (interstitial flow), just as tissue perfusion with interstitial fluid motion in the systemic system are not all lymphatics.
β-Amyloid is a gray matter protein. Its presence is postulated to be necessary for proper synaptic formation (Parihar and Brewer 2010). Therefore, the route for β-amyloid clearance into CSF should be within gray matter peri-arterial VRS. Indeed, this is the shortest distance to the CSF-filled subarachnoid space. Nevertheless, as has been repeatedly shown, interstitial fluid motion within gray matter is primarily diffusional in nature (Orešković and others 2017; Oshio and others 2005; Papadopoulos and Verkman 2013) Therefore, β-amyloid clearance has to occur at this slow rate, not with the more rapid bulk interstitial flow. Indeed, a recent study demonstrated that glymphatic solute transport does not require bulk flow (Asgari and others 2016). It seems prudent to conclude that the glymphatic system is not synonymous with bulk interstitial flow as has been proposed previously (Iliff and others 2012).
For systemic lymphatic
Interstitial flow in the peri-arterial VRS generated by water influx into the peri-capillary VRS is retrograde, namely, flow directs toward the surface of the brain against the direction of arterial flow. This flow is not strong enough to create bulk flow. Nevertheless, together with intermittent closure of the passage by arterial pulsation, the flow is strong enough to prevent backflow, thereby effecting one-way glymphatic flow toward the subarachnoid space (Fig. 7). Indeed, the study which demonstrated that glymphatic solute transport does not require bulk flow, indicated that arterial pulsation facilitated transport (Asgari and others 2016).
The debris clearance system in the brain equivalent to the systemic lymphatic system has been shown to be responsible for β-amyloid clearance and, perhaps, the pathophysiology of Alzheimer’s disease (Suzuki and others 2015; Igarashi and others 2014a). We propose the continued use of the term glymphatic to describe this system (Fig. 7). Nevertheless, we strongly believe that use of the term glymphatic should be limited to the system involved in such a function and should not be extended to describe interstitial fluid motion, especially bulk interstitial flow, which has a different functionality.
CSF Circulation
It is apparent that a continuous fresh supply of fluid into the ventricular system is the main force for CSF circulation. It was once thought that choroid plexus was the sole provider of ventricular CSF. This classic “Choroid Plexus Hypothesis” has been replaced by a newer concept that choroid plexus plays only a minor role in CSF production. Nevertheless, continuous fluid influx into the ventricular system remains a necessary condition for proper CSF circulation. This condition can be fulfilled by influx of interstitial fluid through bulk flow in the peri-venous VRS (Fig. 7).
AQP-1 knock-out mice showed a maximum of 20% reduction in CSF volume (Oshio and others 2005). Furthermore, in vivo non-invasive water dynamics studies on AQP-1 knock-out mice showed virtually no significant change in water influx into the ventricular CSF from the systemic circulation compared to wild-type animals (Igarashi and others 2014b). In contrast, water influx into ventricular CSF from the systemic circulation is significantly reduced in AQP-4 knock-out mice which exhibit interstitial flow dysfunction (Igarashi and others 2014b).
The direction of net water movement through the choroid plexus epithelium favors CSF production. Nevertheless, modern investigations of the molecular basis for the functionality of choroid plexus epithelium suggest the process of water movement may not be dynamic or fast (Boassa and Yool 2005; Brown and others 2004; Praetorius and Nielsen 2006). CSF production by the choroid plexus may rather be to secure basic CSF homeostasis. AQP-1 is further suggested to work as a gated cation channel, not simply a water channel (Boassa and Yool 2005). The presence of “dark cells” in choroid plexus epithelium further indicates bi-directionality of water movement through AQP-1 (Boassa and Yool 2005).
In 1958, Wislocki and Ladman first described that there are two different cell types in choroid plexus epithelium, i.e., “light” and “dark” cells, based on their anatomical appearance on microscopic examination (Wislocki and Ladman 1958). The dark cells comprise 5% to 10% of the epithelium and are distinguishable in having a dark, electron-dense cytoplasm visible on both light and electron microscopy. The widely accepted hypothesis is that dark cells are associated with a state of enhanced reabsorption of excess CSF (Dohrmann 1970; Wislocki and Ladman 1958).
Although speculative, the same argument can be made for the functionality of AQP-4 in ependymal cells which also show “dark” cells (Mitro and others 2013). The function of ependymal cells may be to prevent large “leakage” of CSF into brain parenchyma. Indeed, certain AQP-4 gene polymorphism and dysfunction of AQP-4 in ependymal cells are suggested to be a pathophysiological factor for leukoaraiosis, a condition characterized by excess water in peri-ventricular white matter (Yadav and others 2014).
Conclusion
The three barrier systems, namely BBB, BCSFB, and OBB, with tight junction
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by grants from the Ministry of Education, Culture, Sports, Science, and Technology (Japan).