
Abstract
Opa-interacting protein 5 (OIP5) is a member of the cancer-testis antigen (CTA) family that elicits a spontaneous antitumor immune response. The failure of current immunotherapies for glioma has prompted the search for novel biomarkers that may be utilized as therapeutic targets. This study aimed to investigate whether OIP5 serves as a target for malignant glioma immunotherapy. Glioma specimens from 53 adult patients were evaluated for OIP5 expression by immunohistochemical (IHC) staining, and the correlation of OIP5 expression with World Health Organization (WHO) tumor grade was analyzed. Endogenous expression of OIP5 in glioma cell lines was determined via real-time polymerase chain reaction (RT-PCR). Using lentiviral siOIP5, the effect of OIP5 gene knockdown on proliferation, cell cycle, and apoptosis in U251 glioma cells was studied. The results show that OIP5 is overexpressed in glioma tissues and is correlated with WHO tumor grade (P < 0.001). However, OIP5 protein expression is barely detectable in normal adult brain tissues. MTT assays and analysis using the Celigo Imaging Cytometry System reveal that the silencing of OIP5 inhibits U251 cell growth. Cell cycle assays and Annexin V staining show that OIP5 silencing disrupts the balance of the cell cycle and increases U251 cell death. These results indicate that OIP5 is upregulated in malignant glioma specimens but barely detected in normal brain tissues. OIP5 knockdown inhibits the biological function of glioma cells, reinforcing that OIP5 may serve as an immunotherapeutic target for malignant glioma.
Introduction
Despite improvements in surgical interventions, postoperative radiotherapy, and adjuvant chemotherapy, the median survival of individuals with malignant gliomas remains to be less than 2 years. 1 Moreover, conventional oncologic treatment has been associated with significant side effects. 2,3 Current treatments for malignant gliomas have generally failed, thus prompting investigators to develop novel and targeted therapeutic options. Cancer immunotherapy approaches have become a research interest because of their ability to circumvent drug cross-resistance, fewer adverse effects, and persistent antitumor activity. 4 However, several immune-based treatments for glioblastoma have failed despite decades of investigations. 5 Based on past clinical trials, the most promising immunotherapy for malignant gliomas is an approach driven by biomarkers for appropriate patients selected by cerebral spinal fluid DNA analyzing. 6 Cancer-testis antigens (CTAs) are potential tumor immunotherapy target because these are present in various cancers and are barely detectable in normal adult tissues. 7 Opa-interacting protein 5 (OIP5), which was discovered using the yeast 2-hybrid approach, is a member of CTA family. 8 However, studies on OIP5 expression in malignant glioma are limited and often with conflicting findings. 9,10 This study investigated the expression of OIP5 in specimens from adult glioma patients and analyzed its correlation to glioma clinicopathological grade. Moreover, the effect of OIP5 knockdown on the biological function of glioma cells was examined to assess whether OIP5 may be utilized as a potential immunotherapeutic target for malignant glioma.
Materials and Methods
Ethics Statement
All study protocols for the handling of human tissues in this research were approved by the Ethics Committee of the Second Affiliated Hospital of Xi’an Jiaotong University (Xi’an, Shannxi, China). This study was conducted in full accordance with ethical principles, including the World Medical Association Declaration of Helsinki and its later amendments, and local legislation. Written informed consent to use excess pathological specimens for research purposes was obtained.
Patients and Tissue Samples
Fifty-three patients with glioma were included in this study. All patients had initial surgery performed at the Second Affiliated Hospital of Xi’an Jiaotong University, and none of the patients had received therapy prior to surgery. The gliomas were classified according to the World Health Organization (WHO) criteria for tumors of the central nervous system. The demographic and clinicopathological information of all participants is summarized in Table 1. Normal brain tissues were obtained from 5 patients with hypertensive intracerebral hemorrhage (ICH).
Demographic and Clinical Characteristics of Patients.

Immunohistochemical (IHC) Evaluation
IHC staining was performed as previously described. 10 Paraffin-embedded tissues were cut into 4-μm-thick sections, deparaffinized in xylene, and rehydrated across an alcohol gradient. Antigen retrieval was performed, and hydrogen peroxide was used to eliminate endogenous peroxidase activity. Sections were then incubated at 4°C overnight with a primary antibody to OIP5 (mouse antihuman, dilution 1:50, Catalog No. TA810617, OriGene, Rockville, MD, USA), and negative controls were incubated with PBS alone. After washing with PBS, sections were incubated with a biotinylated secondary antibody (rabbit serum polyclonal antibody, Zhong Shan, Beijing, China) at room temperature for 30 min. Finally, diaminobenzidine (DAB) was used to develop the color reaction.
The most representative areas were selected to assess the percentage of immunoreactive cells and staining intensity. All immunostained slides were reviewed and recorded independently by 3 researchers who were blinded to the clinical findings of the patients. Based on OIP5 staining intensity, OIP5 positivity was classified as positive, weakly positive, or negative. For statistical analysis, positive was defined as “high OIP5 expression,” while weak positive and negative were defined as “low OIP5 expression.”
Culture of Cell Lines
Four human glioma cell lines (glioblastoma of unknown origin, U373, A172, and U251) purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA) were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (Gibco) in a 5% CO2 humidified atmosphere incubator. The identity of the glioblastoma of unknown origin cell line was confirmed through comparison with a short tandem repeat profile (Amelogenin, CSF1PO, D13S317, D7S820, D5S818, D16S539, vWA, THO1, and TPOX).
RNA Isolation and Quantitative Real-Time PCR (RT-PCR)
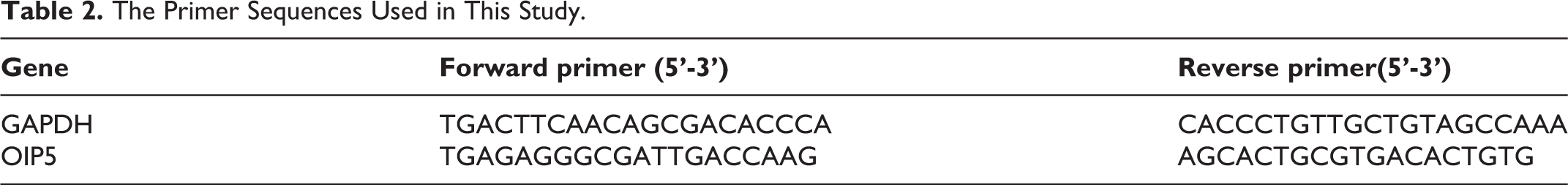
Total RNA from was extracted from the glioblastoma of unknown origin, U373, A172, and U251 cell lines using TRIzol® (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The primer sequences (Genechem, Shanghai, China) used in the study are summarized in Table 2. RT-PCR was conducted using the SYBR PrimeScript RT-PCR kit (Takara) in an Applied Biosystems 7500 Fluorescent Quantitative PCR System (ABI, Foster City, CA, USA). GAPDH was used as internal control for data normalization. All experiments were performed in triplicate. The 2-ΔΔCT method was used to calculate the relative expression of OIP5.
The Primer Sequences Used in This Study.
Lentiviral shRNA Vector Construction and Infection
For the silencing experiments, a lentiviral siRNA vector targeting the human OIP5 gene was produced by Genechem. The designed OIP5 sequences were synthesized and cloned into the pGVX115-green fluorescent protein (GFP) lentiviral vector using the pHelper system and named OIP5-siRNA. The pGVX115-GFP was used as negative control.
Western Blot Analysis
Cells were washed twice with PBS and lysed on ice in ice-cold lysis buffer. Proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The protein samples were transferred onto polyvinylidene difluoride membranes, and bands were visualized using the ECL+plusTM western blotting system kit (Amersham, Arlington Heights, IL, USA) and Kodak X-ray film (Eastman Kodak, Rochester, NY, USA). Primary antibodies (mouse anti-Flag from Sigma and mouse anti-GAPDH from Santa Cruz) were diluted to 1:2,000; GAPDH was used as internal control. The data were analyzed using a gel imaging system.
MTT Assay
Cell proliferation was analyzed using the 3-(4, 5-methylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) colorimetric growth assay from days 1 to 5. Optical density (OD) values were measured at a wavelength of 490 nm to determine the growth rate of cells at different time points. All experiments were repeated thrice to assess cell viability.
Cell Growth Curve
The U251 cells infected with lentivirus were seeded into a 96-well cell plate. The fluorescence intensity of cells was scanned using the Cellomics® ArrayScan® VTI high-content image analysis platform (Thermo, Waltham, MA, USA) for 5 days. The number of cells was automatically calculated.
Cell Cycle Assay
U251 cells infected by lentivirus were seeded into a cell plate and cultured for 72 h. Then, the cells were stained with propidium iodide P4170 (Sigma-Aldrich, St. Louis, MO, USA). Cell cycle distribution was analyzed by flow cytometry (FACSCalibur, BD, San Jose, CA, USA).
Apoptosis Assay
An Annexin V APC apoptosis detection kit (Ebioscience, San Diego, CA, USA) was used to determine apoptosis. Apoptosis was quantified and analyzed using flow cytometry (Guava easyCyte HT, Merck Millipore, Billerica, MA, USA).
Statistical Analysis
All statistical analyses were conducted using SPSS 19.0 software (SPSS, Chicago, IL, USA). The relationship between OIP5 expression and clinicopathological grade was analyzed using the Chi-square test. The student’s t test was used to analyze differences between 2 groups. The data were expressed as
Results
Patient Characteristics
Histology assessment indicated that grade I, II, III, and IV gliomas were detected in 8, 15, 15, and 15 patients, respectively (Table 1). The average age of glioma patients was 51.15 ± 19.32 years (range: 19–78). The average age of the 5 hypertensive ICH patients was 63.8 ± 8.23 years (range: 51–72).
Expression of OIP5 in Glioma Tissues and Cell Lines
OIP5 expression in each tissue section of glioma and normal brain tissues was determined by IHC staining. Dark brown OIP5 staining was mainly observed in the cytoplasm of the glioma tissues (Figures 1B-E), whereas light brown OIP5 staining was only lightly detected in normal adult brain tissues (Figure 1A). Upregulated OIP5 expression levels were correlated with increased pathological grade (Table 3). In grade III/IV gliomas, high OIP5 expression was observed in 27 cases (27/30). However, only 10 cases (10/23) in grade I/II glioma showed high OIP5 expression (P < 0.001). These results show that OIP5 is mainly expressed in gliomas of higher histological grade. RT-PCR experiments revealed that OIP5 mRNA was abundantly or moderately expressed in the U251, U373, A172, and glioblastoma of unknown origin cell lines (Figure 2A-C).

Representative immunohistochemical staining of OIP5 expression in normal brain and glioma tissues. Immunohistochemical staining of OIP5 protein using an anti-OIP5 antibody in (A) normal human brain tissue (×200), (B) grade I glioma (×200), (C) grade II glioma (×200), (D) grade III glioma (×200), and (E) grade IV glioma (×200).
Association Between OIP5 Expression and Clinicopathological Features in Glioma.
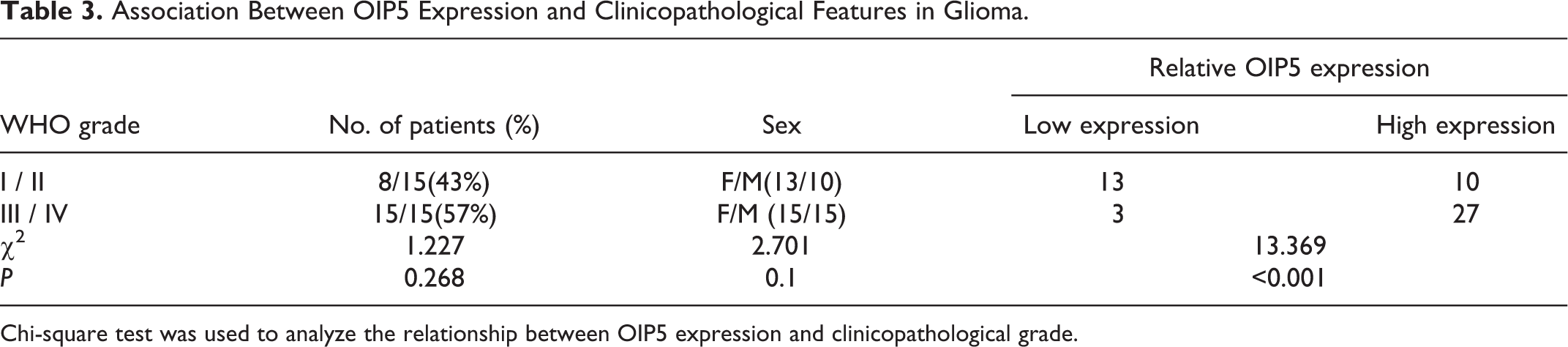
Chi-square test was used to analyze the relationship between OIP5 expression and clinicopathological grade.
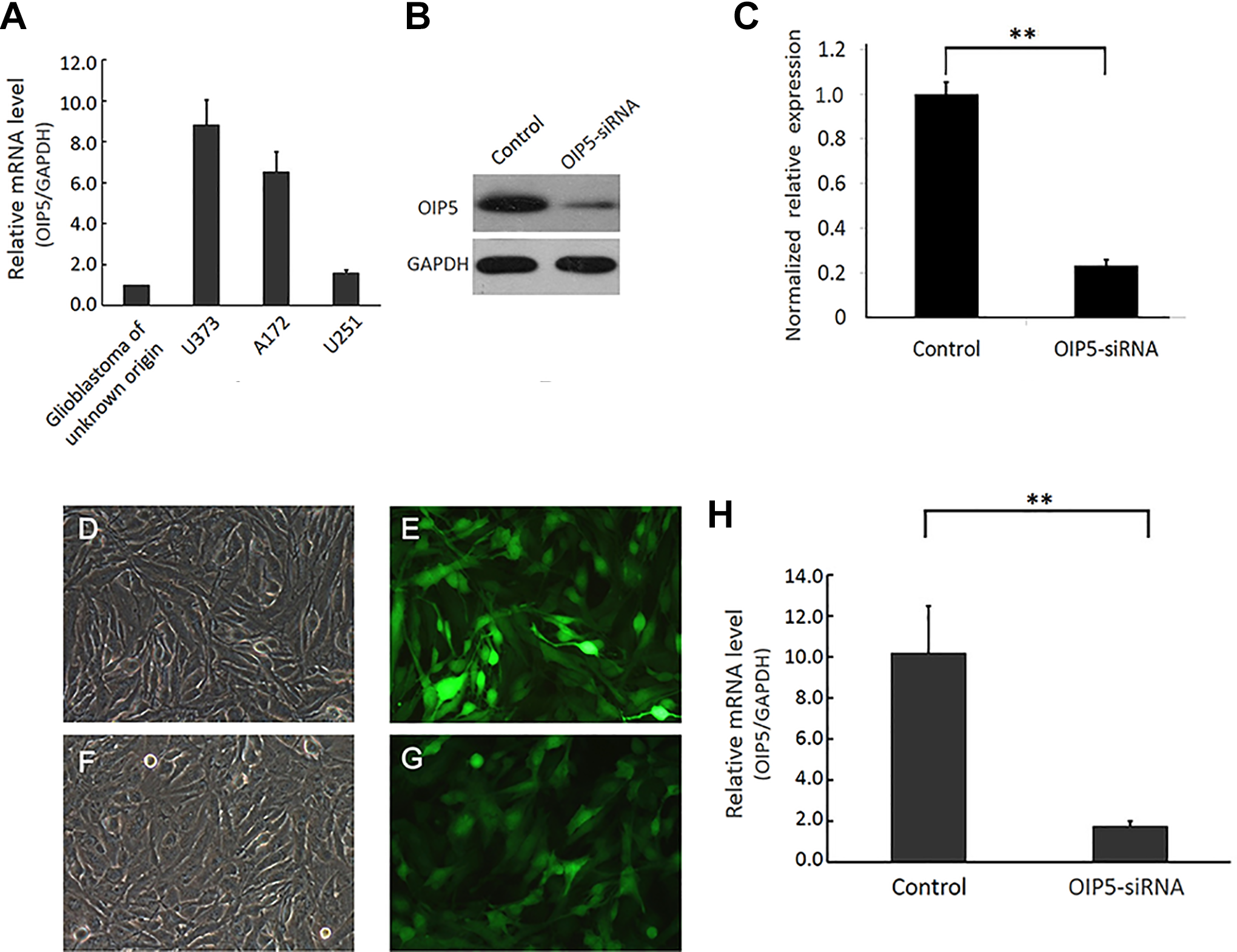
OIP5 expression in glioma cell lines and OIP5–siRNA inhibition of OIP5 expression in cells. (A): RT-PCR revealed that the OIP5 gene is abundantly or moderately expressed in U251, U373, A172, and glioblastoma of unknown origin cell lines. (B) and (C): Effect of OIP5 knockdown on the levels of OIP5 protein in 293 T cells as detected by western blot analysis. D-G: Analysis of GFP expression in U251 cells after infection with a lentiviral vector expressing GFP. (D) and (E): U251 cells in the control group (×200). (F) and (G): GFP expression in the OIP5-siRNA group (×200), reflecting infection efficiency in U251 cells. (H) Infection with Lentivirus-siOIP5 inhibited OIP5 mRNA expression in U251 cells as determined by RT-PCR. (**P < 0.05).
Lentivirus-siOIP5 Infection Suppresses OIP5 Expression in U251 Cells
PGV115-siOIP5/pGV115-siCtrl was used to infect U251cells (Figure 2D-G). To examine the knockdown effect of lentivirus-siOIP5, OIP5 mRNA levels in U251 cells were assessed by RT-PCR. Figure 2H shows that the relative mRNA levels of OIP5 significantly decreased in U251 cells (P = 0.015) after lentivirus-siOIP5 infection compared to the control group.
OIP5 Knockdown Suppresses U251 Cell Growth
To investigate whether OIP5 silencing inhibits U251 cell survival and proliferation, MTT assays and the Celigo Imaging Cytometry System were utilized to determine cell growth curves. Celigo detection revealed that the fluorescence intensity of the OIP5-siRNA group (for 5 days) markedly decreased compared to the control U251 cells (Figure 3). The MTT assay showed that cell growth in the OIP5-siRNA group was significantly suppressed from days 3 to 5 compared to the control U251 cells (Figure 4).

Celigo Image Cytometer analysis shows that knocking down OIP5 significantly inhibits U251 cell proliferation. (A) and (B) The number of cells in each colony was counted over 5 days. (C) and (D) Cell number in the OIP5-siRNA group significantly decreased from days 3 to 5 compared to the control group (P < 0.05).
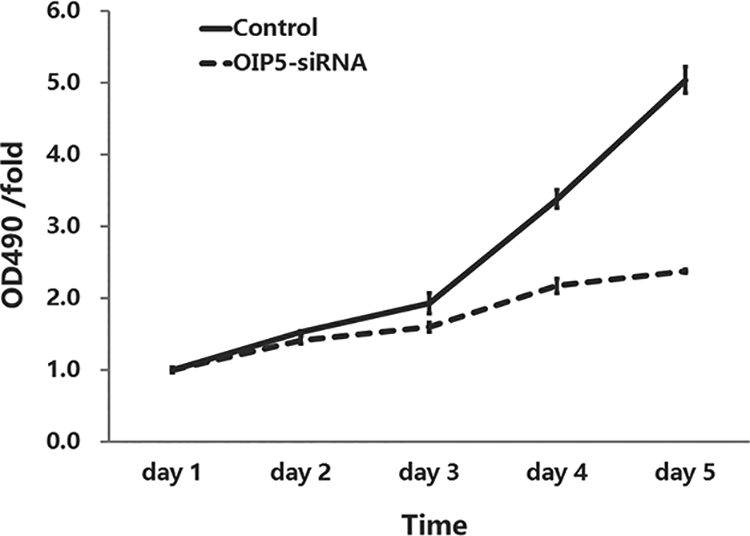
MTT assay showed that OIP5 knockdown inhibits U251 cell proliferation. Infection with lentivirus-siOIP5 significantly inhibited cell proliferation in U251 cells from day 3 to day 5. Cell counts were determined by MTT assay over 5 days (P < 0.05).
OIP5 Silencing Disrupts Cell Cycle Balance in U251 Cells
The effect of OIP5 knockdown on the cell cycle was determined by flow cytometric analysis. The data showed a remarkable increase in the number of cells in the G1 phase (P = 0.017, 53.07 vs. 57.75 ± 0.18%) and G2 phase (P = 0.009, 5.25 ± 1.26% vs. 12.13 ± 0.27%) and a dramatic decrease of cells in the S phase (P = 0.0002, 41.68 ± 0.12% vs. 30.12 ± 0.44%) in the OIP5-siRNA group compared with the control U251 cells (Figure 5).

Knockdown of OIP5 induces cell cycle arrest in U251 cells. The effect of silencing of OIP5 on the cell cycle was evaluated by flow cytometric assay. A significant increase in the cell population in the G1 and G2 phase and a significant decrease of the cell population in the S phase were detected in the OIP5-siRNA group compared with control U251 cells (*P < 0.05, **P < 0.01).
Knockdown of OIP5 Increases Cell Death in U251 Cells
Annexin V staining was used to evaluate the effect of OIP5 knockdown on apoptosis. Figure 6 shows that the apoptosis rate of U251 cells in the OIP5-siRNA group was remarkably higher than the control group (P = 0.000, 3.09 ± 0.44% vs. 17.18 ± 0.30%).

Knockdown of OIP5 promotes apoptosis in U251 cells. FCM shows that knocking out OIP5 expression increases U251 cell apoptosis compared with the control group (**P < 0.05).
Discussion
The current poor prognosis of individuals with malignant glioma creates an extreme need for the development of novel therapies. Cancer immunotherapy is clearly a revolution in cancer care due to their low toxicity and high specificity. CTAs are a unique group of cancer antigens that could potentially be important antigen targets for cancer immunotherapy. 7 As a member of the CTA family, OIP5 is highly cancer-specific and frequently expressed in various types of cancer. 8 This study reports OIP5 expression in glioma tissues using IHC staining. We revealed that OIP5 fulfills the criterion of CTAs and could potentially be utilized as a prognostic biomarker for malignant glioma.
Using IHC staining, the current study examined OIP5 expression in 53 human glioma tissue samples and observed OIP5 protein overexpression in malignant glioma tissues, whereas trace levels were detected in normal brain tissues. This result indicates that OIP5 fulfills the definition of CTAs in malignant glioma. Freitas et al. identified that the OIP5 gene is uniquely expressed in glioblastoma multiforme (GBM) tumors when compared to normal adjacent brain tissues. 9 It has been suggested that the presence of OIP5 in GBM tissues might elicit a spontaneous immune response, and this could possibly result in an antitumor effect. Stratification of glioma tissues based on clinical grade showed that OIP5 protein expression levels in III/IV grade gliomas are significantly higher compared to I/II grade gliomas. This is in line with the previous findings, which show that OIP5 is mainly expressed in tumors of higher histological grade. 11 -14 In lung, renal, AML, esophageal carcinomas, and gastric colorectal cancer, increased OIP5 mRNA abundance is correlated with poor prognosis. 12 -17
OIP5 encodes Opa (Neisseria gonorrhoeae opacity-associated)-interacting protein 5 (also known as CT86, MIS18B, LINT-25, MIS18beta, hMIS18beta, and 5730547N13Rik), which plays an important role in the recruitment of CENP-A through the mediator Holiday junction recognition protein. 13 Using siRNA-mediated knockdown studies, we found that OIP5 silencing significantly inhibits glioma cell survival. Previous studies have shown that the JAK2/STAT3 pathway in nasopharyngeal carcinoma, 18 BMPR2/JUN/CHEK1/RAC1 in liver cancer, 19 miR-139-5p/NOTCH1 in breast cancer, 12 and mTORC2 and p38/PTEN in hepatocellular carcinoma, 20 and E2F1 signaling in glioblastoma 21 are involved in OIP5-regulated tumor progression. These pathways are primarily associated with tumor cell proliferation and migration. The current results also revealed that OIP5 knockdown causes significant growth inhibition, apoptosis, and cell cycle arrest in U251 glioma cells. These findings suggest that OIP5 downregulation contributes to the inhibition of glioma.
Our data show that OIP5 is upregulated in malignant glioma but barely expressed in normal brain tissues. Further evidence suggests that OIP5 could be potentially utilized as an immunotherapeutic target in malignant glioma.
This study is currently at the preliminary stage and thus requires further studies to investigate the mechanisms of OIP5 in terms of glioma biological function. Moreover, the strategy of immunotherapy targeting on OIP5 needs to be developed and verified in vitro and in vivo.
Supplemental Material
Supplemental Material, LetPub_-_Certificate_2020(29) - Opa-Interacting Protein 5 Expression in Human Glioma Tissues Is Essential to the Biological Function of U251 Human Malignant Glioma Cells
Supplemental Material, LetPub_-_Certificate_2020(29) for Opa-Interacting Protein 5 Expression in Human Glioma Tissues Is Essential to the Biological Function of U251 Human Malignant Glioma Cells by Libo Chen, Ruizhi Wang, Ligui Gao and Wei Shi in Cancer Control
Footnotes
Acknowledgments
Authors’ Contributions
LC and RW conceived and designed the study. LC, LG and WS performed the experiments. RW and WS wrote the paper. LG and LC reviewed and edited the manuscript. All authors read and approved the manuscript.
Availability of Data and Materials
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics Approval and Consent to Participate
All study protocols for the handling of human tissues in this research were approved by the Ethics Committee of the Second Affiliated Hospital of Xi’an Jiaotong University.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Shaanxi Provincial Scientific and Technological Research Projects under Grant <2012SF2-03-05>.
Patient Consent for Publication
Written informed consent to use excess pathological specimens for research purposes was obtained from all participants.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.