
Abstract
Small-cell lung cancer (SCLC) represents the progressive form of lung cancer. Patients with SCLC have poor prognosis, partially due to drug resistance. Therefore, understanding the underlying mechanism for drug resistance in SCLC is needed to improve clinical outcomes. The concentrations of heat shock protein 90α (HSP90α) in medium were detected by enzyme-linked immunosorbent assay. The protein levels were detected by Western blot. Cell apoptosis was detected by propidium iodide staining in cell lines or terminal deoxynucleotidyl transferase dUTP nick end labeling staining in tumor sections. Doxorubicin (DOX) was administered into cultured cell lines or intraperitoneally injected into xenograft mouse to induce apoptosis. In SCLC cell lines, either DOX or ABT-737 increased extracellular HSP90α levels, which attenuated the percentage of apoptotic cells. Extracellular HSP90α activated Ak strain transforming (AKT) and β-catenin signaling and inhibited glycogen synthase kinase 3β (GSK3β) signaling. In the xenograft mouse model, extracellular HSP90α promoted tumor development and inhibited apoptosis of tumor cells. Heat shock protein 90α attenuates the efficacy of anticancer drugs in SCLC cells through AKT/GSK3β/β-catenin signaling.
Introduction
Lung cancer is one of the most common cancers worldwide and is one of the leading causes of cancer-related death. 1 The incidence and fatality rates of lung cancer remain high even after reductions in smoking prevalence. 2 Among all lung cancers, small-cell lung cancer (SCLC) represents about 10% to 15% of all cases and strongly correlates with cigarette smoking. 3 Small-cell lung cancer attracts increasing research attention due to its rapid growth and progress, metastasis at early stage, and rapid drug resistance after primary sensitive response. 4,5 The cure rate is 15% to 20% among patients with SCLC after combination of chemotherapy and radiotherapy. In addition, patients with SCLC have poor survival period of about 10 to 12 months. 6 Therefore, improved understanding of the mechanism that regulates the development and progress of SCLC is urgently needed to develop effective strategies for SCLC therapy.
Heat shock protein 90 (HSP90) is an important chaperone protein associated with cell growth, cell proliferation, and cell differentiation. 7,8 More importantly, as an essential molecular chaperone, HSP90 plays crucial roles in stress response and stabilization of mutant proteins. 9,10 Accumulating evidences have implicated HSP90 in the development of various tumors. Heat shock protein 90 promotes prostate cancer invasion via initiating mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) kinase/ERK signaling pathway and inhibiting E-cadherin expression. 11 When treated with a HSP90 inhibitor PU-H71 in the early stage of Janus kinase-dependent acute lymphoblastic leukemia (ALL) murine models, the disease was significantly attenuated. Furthermore, HSP90 inhibitor improved the survival of ALL mice. 12 In melanoma models, HSP90 inhibition by ganetespib increased the expression levels of interferon response genes, which further enhanced T-cell-mediated killing of melanoma cells and the efficiency immunotherapies using anti-cytotoxic T-lymphocyte-associated protein 4 and anti-programmed cell death protein 1. 13 All of these studies demonstrate HSP90 could potentially serve as a therapeutic target. Therefore, in the present study, we aimed to investigate the role of HSP90 in SCLC.
Materials and Methods
Cell Lines and Drugs
The human SCLC cell lines H69 (American Type Culture Collection, ATCC No: HTB-119), H82 (ATCC No: HTB-175), and H146 (ATCC No: HTB-173) were purchased from American Type Culture Collection (Manassas, Virginia). These cells were cultured in Roswell Park Memorial Institute (RPMI) 1640 (Invitrogen, Pleasanton, California) with 10% fetal bovine serum, 100 U/mL penicillin, and 100 g/mL streptomycin in humidified 5% (vol/vol) CO2 at 37°C incubator.
Anti-heat shock protein 90α (Hsp90α; 2G5-G3, ab79849) antibody was purchased from Abcam (Cambridge, Massachusetts). Human recombinant (hr) Hsp90α (SPR-101C) and hrHsp90β (SPR-102C) were purchased from Stressmarq Bioscience (Victoria, Canada). Doxorubicin (DOX, #25316-40-9, LC Laboratories, Woburn, Massachusetts) and ABT-737 (ABT, Abcam, ab141336) were dissolved according to the manufacturer’s instructions. Ak strain transforming (AKT) inhibitor MK-2206 (Selleck, S1078) was purchased from Selleck (Houston, Texas).
Enzyme-Linked Immunosorbent Assay
To detect secreted Hsp90α levels, a total of 2 × 105 cells were plated overnight. Then, the cells were replenished with complete media 24 hours before harvest. Then, the medium was collected, followed by removing debris with centrifugation (5 minutes, 1200g). Heat shock protein 90α levels were detected using Hsp90α enzyme-linked immunosorbent assay kit (Cusabio, CSB-E13462 h, Houston, Texas) according to the manufacturer’s instructions. All detections were performed at least 3 times.
Western Blot
The SCLC cells were harvested and washed using phosphate-buffered saline (PBS). Then, radioimmunoprecipitation assay buffer was used to lysate tumor cells on ice. Equal amounts of proteins were electrophoresed in sodium dodecyl sulfate polyacrylamide gel electrophoresis, and Western blot was performed as previously described. 14 The information of antibodies was as follows: extracellular heat shock protein (eHSP)α, eHSP90β, HSP90α (CST, #8165), HSP90β (CST, #5087), phospho-Akt (Thr308; D25E6) XP rabbit mAb (CST, #13038), phospho-Akt (Ser473; D9E) XP rabbit mAb (Abcam, #4060), Akt (pan; 11E7) rabbit mAb (CST, #4685), phospho-GSK3β (Ser9; D3A4) rabbit mAb (CST, #9322), GSK3β (Abcam, ab93926), non-phospho (active) β-catenin (Ser33/37/Thr41; D13A1) rabbit mAb (CST, #8814), β-catenin (D10A8) XP rabbit mAb (CST, #8480), and β-actin (13E5) rabbit mAb (CST, #4970). Signals were assessed using GeneTools software. β-Actin was used to normalize the intensity of band signals.
Cell Apoptosis Assay
To detect cell apoptosis, a total of 2 × 106 cells were seeded in 75 cm2 flasks. After 24 hours, the cells were fixed with 4% (vol/vol) paraformaldehyde and resuspended in 1 mL PBS. Then cells were stained with 12.5 μg/mL propidium (PI). One hour later, DNA content was detected under a fluorescent microscope.
Terminal Deoxynucleotidyl Transferase dUTP Nick End Labeling
The tissues in paraffin were cut into 5-μm sections. Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay was performed using a cell apoptosis detection kit (MK1020; Boster, Wuhan, China) according to the manufacturer’s protocols, as described previously. 15 Cells with brown nuclear staining in the sections were TUNEL positive.
Xenograft Mouse Model
A total of 1 × 106 tumor cells were subcutaneously injected into the flank of nude mice. Doxorubicin was administered by intraperitoneal injection at 4 mg/kg, once every other day during the whole experiment. Recombinant protein (100 μg/kg) or antibody (1 μg/kg) was intraperitoneally injected after being dispensed in PBS alternatively. Tumor volumes were determined every 3 days and calculated by the formula: V = L × W2/2. This study was approved by Institutional Animal Care and Use Committee of First Affiliated Hospital of Anhui Medical University.
Statistics
All data were presented as mean ± standard error of mean (SEM). The significances among groups were calculated with 1-way analysis of variance followed by Tukey post-test. *P < .05, **P = .01, ***P = .001, and † P > .05, respectively, compared to control. # P < .05, ## P = .01, ### P = .001, and ‡ P > .05, respectively, compared to model or indicated.
Results
Treatment of DOX or ABT-737 Induces Extracellular HSP90α in SCLC cells
To explore the role of HSP90α in SCLC, we treated SCLC cell lines H69, H82, and H146 with 200 nM ABT-737 (ABT) or 40 nM DOX for 96 hours, both of which are known to induce cell apoptosis. The cell survival rates of H69 (Figure 1A), H82 (Figure 1B), and H146 (Figure 1C) were significantly decreased after ABT or DOX treatment. Extracellular HSP90α levels in conditional medium of H69 (Figure 1D), H82 (Figure 1E), and H146 (Figure 1F) were significantly increased after ABT or DOX treatment. Western blot assay showed that extracellular HSP90α protein levels increased 48 to 96 hours after ABT treatment, while extracellular HSP90β expression levels showed no significant difference (Figure 1G). Moreover, 72-hour treatment with ABT or DOX both significantly increased extracellular HSP90α, as compared to control, in all the 3 cell lines. While only in H146 cells, ABT treatment was more effective than DOX treatment (Figure 1H). These data demonstrated that HSP90α was increased in SCLC cell lines by ABT or DOX treatment.
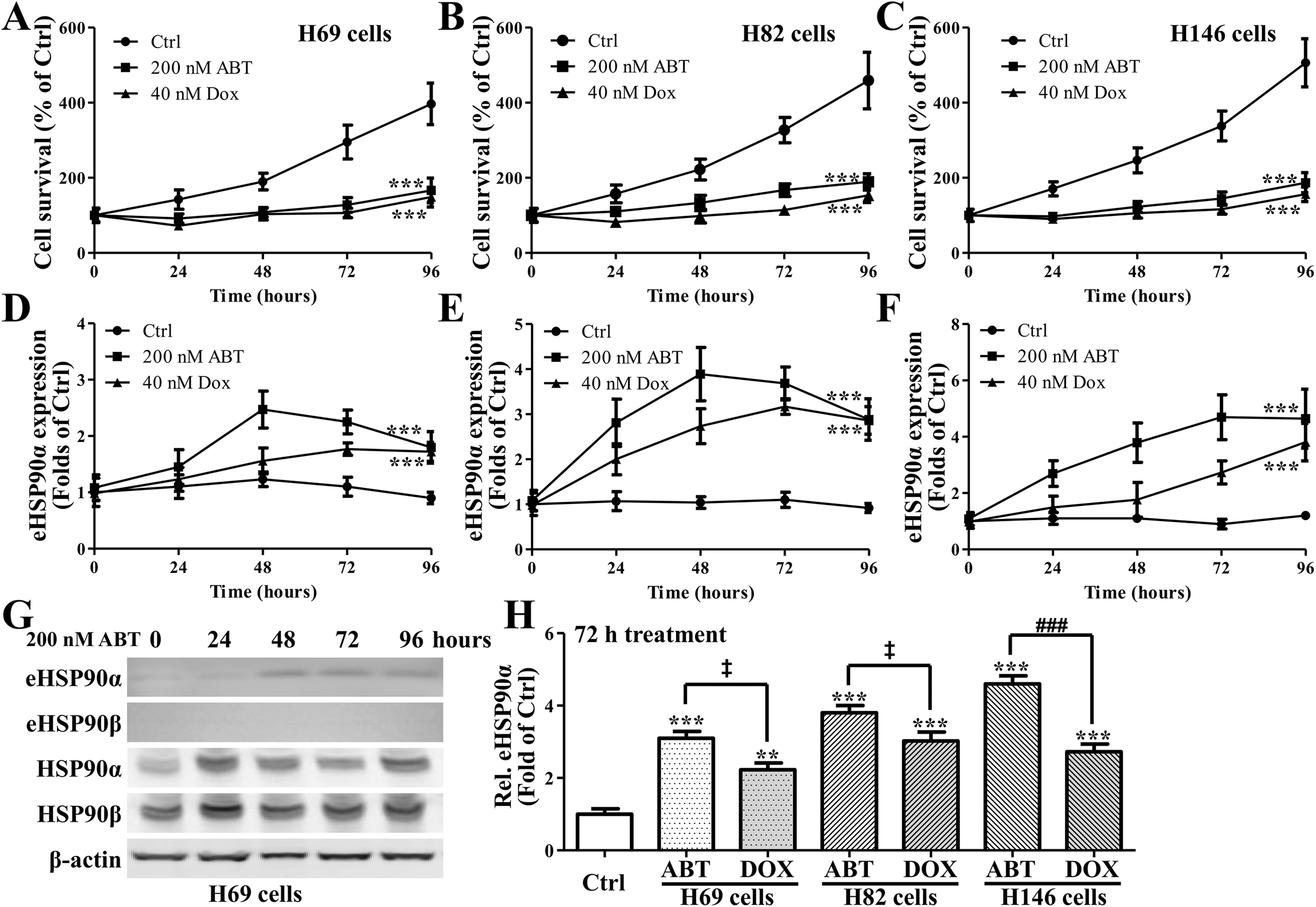
Cell viabilities of (A) H69, (B) H82, and (C) H146 cells under the treatment of ABT-737 or doxorubicin. Enzyme-linked immunosorbent assay (ELISA) assay revealed the existence and concentrations of extracellular heat shock protein 90α (HSP 90α) in the medium of (D) H69, (E) H82, and (F) H146 cells under the treatment of ABT-737 or doxorubicin; (G) representative and (H) summary of Western blot assay for the existence and quantities of HSP 90α and β in culture medium of various cell types (as in eHSP90α and eHSP90β), and the amount of intracellular HSP 90α and β. *P < .05, **P = .01, ***P = .001, and † P > .05, respectively, compared to control. # P < .05, ## P = .01, ### P = .001, and ‡ P > .05, respectively, compared to models or indicated.
Extracellular HSP90α, but not HSP90β, Attenuates DOX- or ABT-737-Induced Apoptosis in SCLC cells
As extracellular HSP90α expression levels in SCLC cells increased after ABT or DOX treatment, we next investigated the role of HSP90α in apoptosis. As shown in Figure 2A, ABT treatment induced apoptosis, whereas addition of 40 μg/mL hrHSP90α protein inhibited apoptosis. The cell survival of SCLC cells decreased after 200 nM ABT treatment. Addition of HSP90α significantly increased survival of H69 (Figure 2B), H82 (Figure 2C), and H146 (Figure 2D) cells in a dose-dependent manner, while HSP90β exhibited almost no influence on cell survival. Moreover, during DOX-induced apoptosis, extracellular HSP90α, but not HSP90β, increased cell survival in all 3 SCLC cell lines (Figure 2E, F, and G). These data demonstrated that HSP90α, rather than HSP90β, attenuated apoptosis.

Apoptosis of cells after different treatments were tested and presented as (A) representative and (B-G) summaries of cell survival rates. Concentrations of human recombinant heat shock protein 90 (hrHSP90) were 10 and 40 μg/mL. *P < .05, **P = .01, ***P = .001, and † P > .05, respectively, compared to control. # P < .05, ## P = .01, ### P = .001, and ‡ P > .05, respectively, compared to models or indicated.
Extracellular HSP90α Activates AKT Signaling to Inhibit GSK3β
We next studied the influence of HSP90α on related signaling pathways. As shown in Figure 3A, B, and C, hrHSP90α treatment increased the expression levels of p-AKT (T308, S473), p-GSK3β (S9), and GSK3β in all 3 cell lines. In addition, hrHSP90α significantly activated AKT signaling (Figure 3A and B) and expression levels of p-AKT (S473; Figure 3C), as well as inhibited GSK3β signaling (Figure 3D), while hrHSP90β treatment had no influence on AKT or GSK3β signaling.

Extracellular heat shock protein 90α (HSP90α) inhibits glycogen synthase kinase 3β (GSK3β) via the activation of Ak strain transforming (AKT) signaling. A, Western blot assay of phosphorylation of AKT (p-AKT; T308), p-AKT (S473), AKT, phosphorylation of GSK3β (p-GSK3β) and GSK3β in H69, H82, and H146 cells after 40 μg/mL human recombinant heat shock protein 90 (hrHSP90) treatment. B, Summaries of p-AKT (308) and AKT levels in (A). C, Summaries of p-AKT(S473) levels in (A). D, Summaries of p-GSK3β and GSK3β levels in (A). *P < .05, **P = .01, ***P = .001, and † P > .05, respectively, compared to control. # P < .05, ## P = .01, ### P = .001, and ‡ P > .05, respectively, compared to models or indicated.
Extracellular HSP90α Activated β-Catenin Signaling
We next investigated the role of HSP90α in β-catenin signaling. In all of the 3 SCLC cells (H69, H82, and H146), protein expression levels of active and total β-catenin significantly increased after being treated with 40 nM hrHSP90α (Figure 4A and B). However, hrHSP90β treatment had no influence on β-catenin signaling (Figure 4B). Moreover, AKT inhibition by MK-2206 repressed phosphorylation of AKT (p-AKT, T308) and GSK3β (p-GSK3β, S9), indicating that AKT inhibition could partially reverse the phosphorylation status of various proteins after hrHSP90 treatment. These data demonstrated that HSP90α, but not HSP90β, activated β-catenin signaling.

Subsequently after the inhibition of glycogen synthase kinase 3β (GSK3β), extracellular heat shock protein 90α (HSP90α) could then activate β-catenin. A, Western blot assay of active β-catenin and β-catenin in H69, H82, and H146 cells after 40 μg/mL human recombinant heat shock protein 90 (hrHSP90) treatment. (B) Summaries of active β-catenin and β-catenin levels in A. (C) Akt inhibitor MK-2206 could partly reverse the phosphorylation states of various proteins after hrHSP90 treatment. *P < .05, **P = .01, ***P = .001, and † P > .05, respectively, compared to control. # P < .05, ## P = .01, ### P = .001, and ‡ P > .05, respectively, compared to models or indicated.
Extracellular HSP90α Increases Tumor Size and Apoptosis of Tumor cells in Xenograft Mouse Model
Given the fact that HSP90α increased in SCLC cells with attenuated apoptosis and activated AKT and β-catenin signaling in vitro, we next verified the role of HSP90α in vivo. As shown in Figure 5A, in the xenograft mouse model, intraperitoneal injection of hrHSP90α increased the tumor volume, while hrHSP90β treatment had no influence on tumor growth as compared to vehicle. However, administration with 2G5-G3 (the anti-secreted HSP90αmAb) inhibited tumor growth (Figure 5A). In the tumor area, the expression levels of p-AKT (T308), active β-catenin, and p-GSK3β were significantly increased after hrHSP90α treatment, while there were no differences in p-AKT, active β-catenin, and p-GSK3β levels after hrHSP90β treatment, compared to vehicle (Figure 5B and 5C). More importantly, 2G5-G3 treatment decreased the expression levels of p-AKT, p-GSK3β, and especially active β-catenin, which showed significant decrease when compared to vehicle (Figure 5B and C).

Extracellular heat shock protein 90α (HSP90α) but not HSP90β increased the phosphorylation levels of Ak strain transforming (AKT) and glycogen synthase kinase 3β (GSK3β) while enhanced the expression of active β-catenin. (A) Tumor volumes of xenograft mouse after various treatments. (B) Representative and (C) summary of the phosphorylation and expression levels in tumor areas. (D) Representative and (E) summary of the transferase dUTP nick end labeling (TUNEL) staining indicating the apoptosis of tumor cells under different treatment. *P < .05, **P = .01, ***P = .001, and † P > .05, respectively, compared to control. # P < .05, ## P = .01, ### P = .001, and ‡ P > .05, respectively, compared to models or indicated.
Treatment with hrHSP90α significantly inhibited apoptosis in tumor area, while hrHSP90β treatment had no influence on apoptosis. In addition, 2G5-G3 treatment significantly promoted apoptosis as compared to vehicle or hrHSP90α treatment (Figure 5D and E). These data demonstrated that extracellular HSP90α, but not HSP90β, promoted SCLC tumor growth, AKT, and β-catenin signaling, as well as inhibited GSK3β signaling and apoptosis.
Discussion
In this study, we explored the crucial role of HSP90α in SCLC. We showed that the expression levels of HSP90α in SCLC cell lines were significantly increased after DOX or ABT treatment, which led to apoptosis in SCLC cells. Addition of hrHSP90α to SCLC cells attenuated the percentage of apoptotic cells. Mechanically, HSP90α inhibited SCLC cell apoptosis via activating AKT and β-catenin signaling and inhibiting GSK3β signaling. In the xenograft mouse model, extracellular HSP90α promoted tumor development and inhibited apoptosis of tumor cells. Moreover, administration with anti-secreted HSP90α mAb 2G5-G3 inhibited tumor development and promoted apoptosis in the tumor area. However, HSP90β treatment exhibited no influence on tumor cell apoptosis or the related signaling pathways. Together, these data suggest that HSP90α attenuates the efficacy of anticancer drugs in SCLC cells through AKT/GSK3β/β-catenin signaling.
Small-cell lung cancer shows good response to initial chemotherapy or radiotherapy. However, SCLC progresses or relapses quickly after primary treatment. Moreover, SCLC metastasis to blood or lymph system is the main cause of SCLC-related death. 16,17 Therefore, the principal problem for SCLC treatment is drug resistance. Previous studies have demonstrated that drug resistance could result from single or multiple factors. At the primary tumor tissues or metastatic sites, there are abundant extracellular matrix around SCLC, which is partially responsible for the enhanced tumorigenesis and resistance to chemotherapy. The activation of tyrosine kinase stimulated by β1 integrin inhibits chemotherapy-induced apoptosis. Therefore, the strategies to block β1 integrin-regulated apoptosis of SCLC cells may be a potential therapeutic approach to drug resistance. 18 After 4 hours of pretreatment with transferrin at 220 to 880 nM, the multiple drug-resistant SCLC cells H69VP showed good response to low dose of artemisinin (IC50 = 5.4 nM), which is close to drug-sensitive H69 cells (IC50 = 2.3 nM). In addition, apoptosis, but not necrosis, increased in artemisinin-treated cells, indicating that combination of transferrin and artemisinin may be the potential strategy against drug resistance SCLC. 19 In our study, HSP90α, but not HSP90β, attenuated apoptosis in SCLC cell lines, which could be the potential mechanism underlying drug resistance in SCLC. It is likely that, in SCLC tumor microenvironment, HSP90α expression level increases and inhibits the apoptosis of SCLC cells, which further promotes SCLC tumor growth. Therefore, it is important to find out the mechanism responsible for the increased HSP90α expression in SCLC tumor microenvironment to further develop effective SCLC therapy.
In antitumor therapies, HSP90 inhibition emerges as a novel choice. The HSP90 inhibitor ganetespib suppressed growth of SCLC cells through inducing G2/M arrest and caspase-3-dependent apoptosis. Ganetespib treatment also downregulated the expression of RIP1, which promoted apoptosis in recombinant human tumor necrosis factor receptor type 1-activated cells. Moreover, the combination of ganetespib and DOX significantly inhibited SCLC tumor growth. 20 Another HSP90 inhibitor NVP-AUY922 showed antitumor effects in SCLC when used together with ABT-737, a B-cell lymphoma 2 inhibitor. The combination treatment promoted apoptosis of SCLC cells via downregulating AKT and ERK signaling and blocking nuclear factor-k-gene binding activation. 21 In our study, we tested the effect of anti-secreted HSP90α mAb (2G5-G3) in SCLC and found that 2G5-G3 treatment promoted cell apoptosis, inhibited AKT and β-catenin signaling, and finally suppressed tumor growth, indicating that 2G5-G3 may serve as a potential candidate in combination with DOX to treat SCLC. However, the fact that 2G5-G3 could not completely block the drug resistance of SCLC hints the existence of other potential pathways for drug resistance in SCLC, which warrants further studies in drug resistance of SCLC.
Ak strain transforming and β-catenin signaling has important roles in tumor development. 22 -26 The AKT kinase inhibitor MK-2206 promoted apoptosis of colorectal cancer cells and significantly inhibited tumor growth via induction of factors that associated with caspase-independent cell death. 27 In liver cancer models, downregulation of β-catenin expression by dicer-substrate small interfering RNAs inhibited tumor burden and reduced the expression levels of genes related to β-catenin. 28 In our study, HSP90α activated AKT and β-catenin signaling and inhibited GSK3β signaling in SCLC cell lines. Moreover, HSP90α treatment also activated AKT and β-catenin signaling and finally promoted tumor growth, indicating that AKT/GSK3β/β-catenin signaling could be very important in SCLC cells.
In summary, we hereby report that DOX or ABT induces extracellular HSP90α levels in SCLC cell lines, which attenuates the percentage of apoptotic cells. Extracellular HSP90α activates AKT and β-catenin signaling and inhibits GSK3β signaling. In the xenograft mouse model, extracellular HSP90α inhibits apoptosis of tumor cells and finally promotes tumor development. These data demonstrate that HSP90α attenuates the efficacy of anticancer drugs in SCLC through AKT/GSK3β/β-catenin signaling, suggesting HSP90α inhibitor 2G5-G3 could be a potential candidate in combination with DOX to treat SCLC.
Footnotes
Authors’ Note
This study was approved by Institutional Animal Care and Use Committee of First Affiliated Hospital of Anhui Medical University (#AJ671).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Anhui Province Key Scientific and Technological Project (1704a0802163).