
Abstract
In this article, I demonstrate the value of the pathological complexity thesis for comparative consciousness research. I discuss how Ristau’s piping plover research illuminates links between life-history complexity and intentionality. How Sinha’s bonnet macaque studies demonstrate how social drivers of pathological complexity predict self-awareness and mindreading capacities. And how Ross’s discussion of elephant life histories and consciousness allows us to compare the phenomenology if humans and elephants. Finally, I address Yilmaz’s discussion of the case for plant consciousness.
Introduction
This article replies to four essay responses to A Philosophy for the Science of Animal Consciousness (Veit, 2023a, 2024) that applied the pathological complexity thesis across diverse taxa.
Because the special issue on my book has been spread across two issues, Yilmaz’s (2024) commentary can be found in Special Issue: Pathological Complexity and the Function of Consciousness in Nature: Part I (Volume 33, Issue 3, June 2025 of Adaptive Behavior), whereas the commentaries by Ross, Ristau, and Sinha can be found in Part 2, that is, the present Volume.
I begin with Carolyn Ristau’s discussion of piping plovers and how the pathological complexity framework can help us to understand their intentionality. In the subequent section, I discuss Anindya (Rana) Sinha’s case study on bonnet macaques, which provides further evidence for the usefulness of the pathological complexity thesis in linking complex social life histories to capacities for self-awareness and mindreading. I then move to offering a detailed analysis of Don Ross’s African elephant case study of the pathological complexity thesis and how we can link their life histories to their dimensions of consciousness, offering a comparative analysis of elephants and humans. Finally, I address Özlem Yilmaz’s discussion of plants and why I do not attribute consciousness to them. This response essay offers an illustration of how comparative consciousness research in ecological context can show the utility of the pathological complexity thesis for understanding the minds of other animals. 1
Intentionality, Pathological Complexity, and the Piping Plover
The first case study of the pathological complexity framework will bring us to plovers, and in particular the piping plover (Charadrius melodus), which Carolyn Ristau has spent many years studying. I have invited Ristau because of her close connection to Donald Redfield Griffin. She worked with him at his lab at Rockefeller University, when he was trying to create a field of cognitive ethology, later writing a biography of him that was published in three volumes (Ristau, 2024). My book was in more ways than one inspired by Griffin and his attempt to study animal consciousness as a natural phenomenon in the wild (Veit, 2025c). If it wasn’t my first book I would have dedicated the book to Griffin, rather than my parents. It is unfortunate that Griffin died long before I started to develop my framework, but inviting Ristau as a former disciple of him was the closest I could come to receiving an insight into how Griffin himself would have responded to my framework. Thus, I am very pleased to hear that “Griffin would agree and I agree with Veit’s insistence that we regard an animal’s consciousness from the animal’s experience” (Ristau, 2025, p. 1). Ristau endorses the gradualist step-by-step bottom-up approach appreciating my dedications to Griffin and likewise endorses my theory for practicing cognitive ethologists, noting that it “is a significant contribution to the study of animal mind and consciousness. Applying his ideas to animal life histories could inspire creative experiments to further explore animals’ mental experiences” (Ristau, 2025, p. 8). While she is sceptical that this solves the hard problem of consciousness, she admits that we have at least made the problem significantly smaller.
While Ristau finds value in my link between the pathological complexity of life history strategies and consciousness, she worries that we will struggle to determine when a species has no consciousness whatsoever. I agree with Ristau that this is a significant problem, but am optimistic that an examination of the life history complexity of Cambrian species compared to modern animals will give us a way to assess the plausibility of consciousness in boundary cases like gastropods and insects.
Ristau also notes that an evaluation of the complexity of life-history trade-offs faced by animals includes “hypotheses about the likely conscious decisions an organism is making at each of these stages and the cognitive processes involved” highlighting that this is also apparent in Ross’s discussion of elephant life histories (Ristau, 2025, p. 1). I agree that this makes the problem harder. But it should not be surprising. Once consciousness exists, it creates a new domain in which things can go right or wrong, in which things can be healthy or pathological. Consciousness is a means to reduce life-history complexity, but it also increases it in other ways, providing the opportunity for further adaptations that shape the subjective experience of animals.
Ristau’s own case study will illustrate this nicely since her response is ultimately focused on applying the “Pathological Complexity Thesis to stages in the life history of piping plovers (Charadrius melodus), suggesting the various decisions they are likely to face at each challenge” (Ristau, 2025, p. 1). Indeed, her approach beautifully shows the usefulness of cognitive ethology, since it allows us to use careful observations of their lives in order to identify the problems animals face in their natural lives. Yet, Ristau also emphasizes a critical gap in current ornithological knowledge when compared to the elephant research discussed by Ross (2025). Unlike the robust longitudinal data available for elephants, the available data for the social dynamics, life histories, and cognitive capacities of plovers is limited. But I will note that this also makes them a better model for most species in the wild that we may apply the pathological complexity thesis to, while emphasizing the need of naturalistic observational studies.
A Simplified Piping Plover Life History
Synthesizing ornithological data from Wilcox (1959), Cairns (1982), and several conservation bodies, Ristau describes the piping plover (Charadrius melodus) as a “small, sand-colored birds, about seven inches long, with white underparts” (Ristau, 2025, p. 2). Their colouration allows them to blend in seamlessly while probing the sand for invertebrates, often appearing as little more than “blobs of sand moving against the sand” (Ristau, 2025, p. 2). This is important since the piping plover is an endangered shorebird species laying their eggs not in a nest in trees, instead relying on beach dunes. One might think that they have low survival rates, but Ristau notes that while their average longevity is approximately 5 years, they exhibit a notably high adult survival rate of 80%. Within the pathological complexity framework, the importance of decisions later in life is relativized to the probability of survival until that point. To illustrate this: If a bird only has a 1% chance of making it to the age of five, then natural selection will have little ‘incentive’ to invest into costly neurocognitive architecture. Even if decisions at such a life stage might be highly complex, they matter comparatively less from a life history perspective.
Much of her data is drawn from her own observations of colonies located on Long Island, and Metompkin Island, a barrier island off the Virginian coast. Ristau characterizes piping plovers as migratory birds of intermediate range between the Caribbean and the Mexican Gulf Coast up through the southeastern United States between Texas and Georgia. Citing Wilcox (1959), Ristau further notes that individuals frequently reclaim their prior breeding territories in the Spring providing them with several important decision points during this life cycle. Very usefully, Ristau offers an analysis of these decision points and their life-history significance for the pathological complexity thesis.
Ristau describes how male piping plovers reach breeding locations ahead of females, initiating territorial claims around early April. She notes that Cairns (1982) documented these territories as consisting of connected zones: nesting sites situated within or adjacent to dune areas, with foraging areas below and closer to the ocean. Ristau explains that “feeding area is used exclusively by the mated pair and offspring” though noting that shared foraging areas located behind the dunes provide crucial alternatives during high tide or when human activity disrupts access (Ristau, 2025, p. 2).
Ristau notes that piping plovers maintain distances between territories. Where space is limited, neighbour aggression is increased, creating stress that compromises parental vigilance during confrontations. Territorial establishment involves charging threats and aerial pursuits to demarcate boundaries. Territory size and defence intensity vary across individuals, which is also impacted by nearby least tern (Sterna antillarum) colonies. While plovers benefit from their loud vocalizations to predators, they are more combative than plovers, and sometimes initiate aggressive interactions that consume energy and divert attention from offspring protection.
Ristau suggests plovers select nest sites more carefully than terns, placing nests on elevated sand rises that withstand high tides better than tern nests. But she also cites Guild et al. (2024) who report that predation and flooding are still major threats to plovers. Territory and nest site selection thus requires significant pathological complexity in their decision-making. She explains that males create multiple nests. While both evaluate potential locations, females make the final selection. Ristau reports that on Long Island beaches, most nests were positioned in dune areas with some grass, allowing effective visual monitoring while reducing visibility. She also emphasizes that delayed nesting attempts correlate with reduced hatching success.
During courtship, males use “calls and aerial and ground displays” to attract females while advertising territorial ownership to rivals (Ristau, 2025, p. 3). She notes that established pairs remain monogamous throughout the breeding season and sometimes continue in the future. Ristau observes that both sexes assess partner characteristics, though females hold final mate selection authority. She emphasizes that female reproductive investment requires considerably greater energy than male contributions, making female choice critical for reproductive success.
Ristau explains that territorial boundaries may shift through the season. She identifies parallel-run displays at the border as the predominant boundary maintenance behaviour, performed by both sexes though males do so more frequently. Though she notes that sometimes more aggressive interactions occur. She poses critical questions, such as: when does territorial defence warrant the investment, given energy costs and increased egg vulnerability when parents focus on defence? These are exactly the kind of problems that increase the pathological complexity of the plover’s life histories.
When it comes to the ultimate goal of life, Ristau reports that females normally produce four eggs with roughly synchronous hatching occurring 4 weeks later. She explains that nest loss in the season’s first half may prompt re-nesting, though these often fail. She cites Wilcox (1959) documenting that mice or rats take individual eggs, while crows, foxes, raccoons and other mammals consume entire clutches, and off-road vehicles or dogs also destroy nests. Ristau adds that gulls frequently predate both eggs and chicks. This is a very precarious situation that highlights the pathological complexity plovers are facing.
Accordingly, Ristau describes several parental defences to protect eggs and offspring. She notes that “parents, nest, eggs, and young, are all very well camouflaged against the sand” (Ristau, 2025, p. 3). Some behaviours exploit camouflage, including silent immobility or low-profile running. Others include a so-called rodent run where plovers wiggle their rumps to appear rodent-like and pretending to incubate away from the actual nest. A similarly misleading behaviour includes so-called Broken Wing Display, though Ristau notes that it is as considerably more energetically costly. It typically occurs when predators approach both nest and parent closely, creating danger for all. Ristau notes that plovers perform this display more frequently during the final pre-hatching week, which makes sense within the pathological complexity framework, since birds have already invested more into their offspring here.
Moving to the beginnings of a new life history, Ristau describes how embryonic vocalizations become audible several days before hatching, heightening parental arousal. Ristau notes that the offspring are “precocial, leaving the nest after 2–3 hours, and beginning to feed, but they generally remain within about 400–500 feet of the nest until able to fly” (Ristau, 2025, p. 4). Ristau observes that plovers possess multiple innate protective strategies but appear to “learn how to use them most effectively” (Ristau, 2025, p. 4). She cites Wilcox (1959) documenting parents knocking down fleeing chicks, who then remained immobile. Ristau notes that immobility proves more effective than movement given their camouflage, which chicks might have to learn. Ristau also notes how parents constantly monitor young, often huddling over them for protection and warmth. Chicks and parents vocalize with each other, which is marked by an increase in the previously described protective behaviours. Rather than just having fixed response patterns, Ristau notes young must learn optimal feeding times and locations, predator detection, and adaptive behaviours. During this period, Ristau explains both young and parents accumulate sufficient weight for later migration. She describes how as young gain independence and associate with others, they also range farther and investigate additional feeding locations.
Southward migration at the end of summer is characterized by Ristau as hazardous. She notes that like all migratory birds, plovers experience substantial mortality “through predation, severe weather, and challenges to their energy resources” (Ristau, 2025, p. 4). Adults return on their own to regions where they previously overwintered, whereas first-year birds congregate in flocks to migrate together. Ristau also notes that juveniles face critical migration decisions: when to depart, whom to fly with, and how to navigate. Ristau emphasizes that navigation relies on “the location of the sun and stars and cues from the earth’s magnetic field guide the juvenile’s migration” (Ristau, 2025, p. 4). Within flocks, positioning matters: air currents from nearby birds can aid or impede flight. Birds regularly reposition themselves, Ristau suggests, possibly preferring to fly near favoured companions. Again, this suggests high pathological complexity, with strategic choices that will have impacts on their survival chances.
Ristau explains that plovers establish seasonal home ranges during winter, showing site fidelity. However, data revealed larger winter ranges and smaller fall ranges, driven by high tides and flooding. Flights typically stayed under 10 km, less than most overwintering shorebirds, and diverse adjacent habitats enabled survival without costly flights, she observes. Raptor predation presents a second challenge. Ristau notes that while some shorebirds flee approaching falcons, plovers crouch: an effective strategy since falcons hunt airborne prey. Warm southern temperatures also aid survival, Ristau adds. She cites Drake et al. (2001), who conclude that population declines stem not from winter mortality but from climate change and human encroachment.
Northward migration to breeding sites around March poses similar hazards, Ristau notes. She cites Wilcox (1959), who reports fewer than 5% of first-year plovers return to their hatching sites. Again, Ristau emphasizes the abundance of trade-offs in plover decision-making: destination, habitat quality, human disturbance, predation risk, departure timing. How do young birds balance optimal habitat and timely arrival against competition for nest sites and the need for spacing from neighbours? At merely 1 year old, the cycle repeats: young plovers begin their own reproductive cycle: acquiring territory, courting, and finding mates.
Much more detail on plovers can be found in Ristau’s article and in the sources on which she draws, but even this brief summary of their life histories underscores the pathological complexity they confront. As I have emphasized repeatedly, the decision problems Ristau describes involve multi-dimensional trade-offs that have plausibly shaped the birds’ phenomenological complexity. Because much of this pathological complexity is driven by the need to assess risks posed by nearby conspecifics and predators, the pathological complexity thesis predicts the evolution of significant mind-reading capacities. This is precisely the direction Ristau pursues, following her life-history analysis with a discussion of field experiments designed to probe the mental lives of plovers that we shall now examine closer.
Field Experiments
Discrimination and Risk Assessment
Because plovers are impacted by predators and human interference, Ristau wanted to investigate whether they can learn to differentiate threatening from benign intruders. To test this, Ristau conducted field experiments using differently dressed human intruders who walked past nests at a significant distance. Afterwards, trials were conducted in which some intruders paused before reaching the nest, approached directly, hovered over it, then departed. Citing her earlier experimental work (see Ristau, 1991b), she notes that plovers showed greater arousal to these threatening intruders even when both walked at a safe distance, demonstrating rapid discriminatory learning. Ristau cites Haig (1983) who reported that at Lake Manitoba sites, plovers would avoid courtship and mating behaviours, when humans were around. Ristau also notes that plovers vary in their responses based on the study sites. Plovers on remote Metompkin Island showed much greater arousal than those at popular Long Island beaches with regular pedestrian traffic, which were only reacted to if they got too close. In an interesting anecdote, Ristau reports that: [O]ne plover who had experienced partial nest predation from an unknown cause behaved in a highly aroused fashion to a person walking a dog down by the shoreline. Upon seeing the dog, she immediately flew from the nest, flying just in front of the dog. The dog avidly followed until the plover led it into the water; the dog got completely wet and the plover flew in a great circle back to her nest. The same human and dog returned another day, and the plover did the same thing, though the dog didn’t go all the way into the water the next time. (Ristau, 2025, p. 6)
Not only does this demonstrate that plovers learn from their previous experiences, but it also suggests that they have some capacity for intentionality, which the next experiments will make even more apparent.
Injury Feigning
Ristau notes that Broken Wing Displays (BWD) occur widely among ground-nesting birds, appearing across “52 different families, encompassing 285 species”, citing de Framond et al. (2022) who suggests that it evolved several times independently in these lineages (Ristau, 2025, p. 6). Ristau argues that predation pressure appears as the fundamental determinant for BWD presence, since it is for instance correlated with higher latitudes, which provide extended daylight and increased predator activity. However, Ristau also emphasizes that BWD represents evolved behaviour, not individually invented by each plover. When she initiated research, many scientists regarded BWD as merely an innate fixed action patters in response to predators, which remains a common view even today. Indeed, she notes that her interest in this question emerged from a workshop where Dennett asserted that these birds simply perform innate responses, not purposeful deceptive behaviour.
In order to investigate this question, Ristau employed the following criteria for a First Order intentional stance: A. The direction of BWDs should usually be adequate to lead an intruder away from the nest. B. The displaying bird should monitor the intruder’s behaviour. C. If the intruder does not follow the displaying bird, the bird should then modify its own behaviour in a variety of ways in response to the intruder’s behaviour so as to keep the intruder away from nest/young. D. The bird should exhibit appropriate flexibility of behaviour in other circumstances. (Ristau, 2025, p. 6)
In the fashion of a true cognitive ethologist, Ristau notes that researchers should ideally observe naturally occurring interactions between parent plovers and local predators like raccoons, “but such occasions are rare and highly unlikely to be observed when they do occur” (Ristau, 2025, p. 6). Instead, they relied on human intruders who would communicate with observers via walkie-talkies, and providing video recordings. She notes that they would sometimes move near offspring, sometimes following plovers during BWDs and sometimes not. Ristau posed several questions: Did plovers display strategic positioning? Plovers repositioned themselves through “walking, running, or flying when it detected the approaching intruder, before it began a BWD” (Ristau, 2025, p. 7). Contrary to the conflict hypothesis, Ristau notes, that plovers did not attempt escape; in all 13 cases involving flying, they flew closer to intruders and usually closer to the intruder’s visual field. Did plovers modify behaviour when intruders didn’t follow? Yes. Ristau points out that plovers stopped displays and reapproached intruders in over half the cases, increased display intensity in almost a third of cases. When intruders followed, Ristau notes, plovers never reapproached. Are BWD directions appropriate for leading intruders away? Yes. Ristau argues that if intruders had followed to final display locations, they would have been farther from nests or young. Finally, did plovers monitor intruders? Despite laterally positioned eyes, which makes the assessment of this question difficult, Ristau argues that photos and videos show birds looking back at intruders during displays.
Ristau interprets these behaviours as supporting First Level Intentionality: plovers want to lead intruders away. She notes plovers exhibit flexibility beyond BWDs, possessing other protective behaviours and performing BWDs in only 40% of approaches during the discrimination experiments discussed in the previous section. She also cites field observations showing that killdeers (Charadrius vociferus), a close relative, rarely perform BWDs when grazing animals approach: “Instead, the killdeer may lunge at the cow’s face, startling it and causing it to veer from its path” (Ristau, 2025, p. 7). This research thus provides strong evidence that the pathological complexity plovers face has led them to evolve “purposeful, specifically, conscious, control of its actions” (Ristau, 2025, p. 7).
Perspective Taking
Ristau suggests that this research also suggests that plovers have the ability to engage in perspective taking, which she argued in earlier research is central for a theory of mind (Ristau, 1991a). She describes how two differently dressed intruders would walk identical paths parallel to nests averaging 20 m. One looked towards dunes where nests were situated; the other looked to the ocean. Each sat before reaching positions opposite nests, maintained their gaze, then resumed walking, and Ristau notes that plovers showed greater arousal to dune-gazing persons, measured by extended nest absence. While she acknowledges that plovers have cues beyond gaze direction, including head orientation and body position, she argues that plovers have shown to be sensitive to intruder attention.
I will note that this lines up with recent research on experience projection, which has shown birds to be acutely aware of other’s mental states (Lurz & Krachun, 2019), and further research would likely show plovers to have much more significant theory of mind capacities than we might have assumed without the aid of the pathological complexity thesis.
Chick Distress Calls
Finally, Ristau describes experiments in which pre-recorded chick distress calls were played to parents during egg or chick stages. Ristau notes that parents responded far more during the chick phase than incubation. She also points out that responses differed widely demonstrating why researchers must document detailed observations, particularly of infrequent behaviours that reveal complex mental capacities that might otherwise be missed. For instance, she highlights one such striking observation:
One parent made repeated, searching flights for over 15 minutes in the area over the sound source, but then abruptly stopped the search flight and flew directly to another location and began searching there. That location was the site of a sound source (SS) in a playback experiment conducted a few days earlier (Ristau, 2025, p. 8).
She also notes that plovers frequently continued searching after sound sources fell silent between trials and searched longer when sources were partially concealed in grasses versus open sand, and that responses also varied depending on whether parents accompanied chicks when calls commenced.
Ristau cites philosopher Stieg approvingly, who has whose offered a detailed analysis of her field work that concluded that “the complexity, adaptability and flexibility of the plovers’ behavior, along with its ability to utilize the content of its representations and to satisfy the conditions of concept attribution” justifies the attribution of intentionality (Stieg, 2008, p. 31). I couldn’t agree more. As Ristau’s discussion makes clear, the pathological complexity of plover life histories is driven by social dynamics: such as assessing conspecific territorial threats, evaluating predator attention and intentions, coordinating with mates and offspring, positioning within migrating flocks. It is under such conditions that we can predict intentionality to evolve.
This case study also illustrates an important methodological point Griffin championed and that I have emphasized throughout my book: consciousness must be studied in natural ecological contexts, not reduced to laboratory tests designed for humans. Ristau’s field experiments reveal cognitive capacities, such as strategic deception, rapid discrimination learning, context-sensitive behavioural adjustment that would be easy to miss in standardized laboratory paradigms that try to maximize internal validity of experiments at the cost of ecological validity. While such research is useful, it should not be our only way of investigating animal minds. The pathological complexity framework guides research by directing attention towards the specific adaptive problems animals face in their life histories, generating testable predictions about which phenomenological capacities evolution should favour.
Plovers exemplify how the pathological complexity thesis transforms animal consciousness research from a top-down enterprise (applying human consciousness criteria to animals) into a bottom-up evolutionary investigation (asking what consciousness does for organisms given their ecological challenges). This is true, even if we have much less data on the life histories of plovers than elephants, showing that my bottom-up approach is useful even under sparsity of data. Ristau’s work demonstrates that we can significantly progress our understanding of animal minds by taking seriously the ethological question Griffin insisted we must ask, that is, what role does mind play for animals in the natural environments they evolved in? Indeed, I suspect that something like an implicit version of the pathological complexity thesis has always been operating in the back of the work of cognitive ethologists like Griffin and Ristau, and my work has tried to bring it to the forefront and develop into a rigorous methodology to show that the behaviourist tradition is mistaken to discard this kind of ecological research. Indeed, the following discussion of another cognitive ethologist’s analysis of the pathological complexity thesis will support this view. But before we do so, let me reiterate that I very much recommend people to read up on the history of cognitive ethology in Ristau’s biography of Griffin which highlights Griffin’s early philosophical arguments that inspired much of my own desire to bring him back to the forefront of animal minds research (see Ristau, 2024; Ristau, & Griffin, 2024).
Bonnett Macaques, Social Drivers of Pathological Complexity, and Self-Awareness
I will now turn to the essay by Anindya (Rana) Sinha (2025) on how the pathological complexity framework can help us to understand the phenomenological complexity of macaques. Like Ristau, Sinha is a cognitive ethologist known for his research on animal minds in the wild, especially bonnet macaques (Macaca radiata). This is why I invited him and was especially interested to see whether he would find my framework useful, since it has been deliberately designed for cognitive ethologists. To my relief, I am glad that he did: My primary question, apparent to me when I began my purely observational cognitive studies in the wild more than three decades ago and one that remains within me even today, is, simply asked, what is consciousness? How do I define this phenomenon in the more than two thousand individual macaques, each of whom I have come to know over the passing years, often very closely, but who are unable to tell me what they are feeling, what they are thinking of, and how they are taking the complex ecological and social decisions that they are? And it is here that Veit’s ideas of phenomenological and pathological complexity strike resounding chords in me. (Sinha, 2025, p. 2)
As Sinha emphasizes, his own work on trying to understand the complex minds of macaques was already focused on assessing the pathological complexity challenges they are facing, “through deep naturalistic observations of their behavioural decision-making, social cognition, and communicative capacities” which allows to catch “glimpses of the pathological complexity that individual macaques may have faced over evolutionary timescales and continue to do so across their strikingly independent life histories” (Sinha, 2025, p. 2). As he emphasizes at the end of his essay: Pathological complexity is best conceptualised and measured using long-term life-history traits and that, more specifically, we can measure the complexity of a macaque’s adaptive traits – manifest through their possibly unique forms of consciousness, as Veit suggests – by actively identifying their life-history strategies, and their temporally and spatially characteristic behavioural response patterns, with their underlying cognitive complexity, all of which may have essentially evolved to promote their long-term survival and fitness. (Sinha, 2025, p. 7)
Like Ristau, Sinha begins by outlining the surprising pathological complexity of their societies. He notes that the demands of forming and sustaining complex social bonds in nonhuman species may have driven the evolution of sophisticated cognitive capacities. He suggests that understanding where other group members stand in social networks could help individuals assess their standing “in the prevailing affiliative and dominance networks within a primate society” (Sinha, 2025, p. 3). Through systematic field observations, Sinha identified a specific competition pattern among female bonnet macaques involving grooming access: in allogrooming supplants, a higher-ranking female approaches and replaces one individual from a grooming pair, both of whom rank below her (Sinha, 1998). He notes that in most three-way encounters, the lower-ranking grooming partner withdraws before the dominant female arrives. Occasionally, however, the intermediate-ranked female leaves instead. When Sinha analyzed subordinate identities across interactions, he discovered that females who were more socially desirable as grooming partners showed greater tendency to hold their ground when approached.
Sinha measured this social attractiveness by calculating the “reciprocal of the coefficient of variation for the duration of allogrooming received by a female from all the other adult females in the group” (Sinha, 2025, p. 3). Sinha notes that this social desirability index captures not just the total amount of grooming attention a female receives from her group, but also how reliably she is sought out as a preferred grooming companion. He later found that individual females appeared to recognize which group members were more socially desirable, could accurately assess the relationships between others, and “appeared to possess egotistical knowledge of their own rank positions, relative to those of others, in the socially constructed dominance hierarchy prevailing within their group” (Sinha, 2025, p. 3).
These findings demonstrate how macaques navigate social pathological complexity: simultaneously tracking dominance ranks, attractiveness levels, and relationship patterns to make adaptive decisions shifting social dynamics. The cognitive sophistication required to evaluate social variables in real-time suggests high evaluative complexity. Like Sinha, I believe that it is this pathological complexity in their social life histories that suggests the need for self-awareness. Like the previous discussion on intentionality in plovers, I hope to make use of Sinha’s insights here to progress our understanding of the evolution of self-awareness.
Sinha situates his findings within broader research traditions. He cites, Bachmann and Kummer’s (1980) pioneering work with hamadryas baboons, which showed that primates might recognize others’ social relationships. However, Sinha notes that their experimental design left ambiguous whether observers truly tracked relationships between others or merely responded to behavioural cues. Sinha argues that his naturalistic observations provided clearer evidence: bonnet macaque females demonstrated awareness of grooming interactions between other group members. Sinha observed that dominant females approaching grooming pairs employed two strategies, either aggressively displacing one partner or peacefully joining and affiliating with one. Their aggressive targeting showed context-dependent patterns. In three-way scenarios, they nearly always attacked the lowest-ranking individual while rarely targeting the intermediate-ranked female, thus contrasting sharply with two-way encounters where they preferentially targeted higher-ranking subordinates (Sinha, 1998).
Sinha argues that this pattern reveals bonnet macaque awareness of relative rank positions which feeds into their ability to flexibly adjust social strategies based on context. The social environment presents macaques with variable, context-dependent challenges resisting simple stimulus-response solutions. As Sinha emphasizes, these decisions require real-time evaluation of who is present, their relative positions in the hierarchy, and what strategy is appropriate for that particular situation, thus providing “striking evidence of Veit’s pathological complexity, rooted spatially and temporally, and requiring an immediate, but conditional, possibly cognitively ideated decision” (Sinha, 2025, p. 3).
Furthermore, Sinha found that when subordinate females decided whether to flee or hold their ground as a dominant approached, two pieces of information proved critical, that is, their awareness of where they themselves stood in the group’s social ladder, and the hierarchical gap between themselves and the incoming individual (Sinha, 1998). From the pathological complexity perspective, this social awareness aids in adaptive challenges. Retreating or remaining carries significant fitness consequences since challenging much higher-ranked individuals risks injury, while unnecessarily yielding forfeits valuable grooming opportunities. This increases paradigmatic pathological complexity due to multi-dimensional trade-offs, thus suggesting explicit conscious evaluation. The social hierarchy proves neither completely predictable nor entirely random. Successfully navigating it requires more than associative learning; it demands evaluative capacity to assess one’s current state, compare it to situational features, and select actions accordingly.
However, Sinha suggests a limitation to this capacity since females could grasp rank disparities between themselves and others without necessarily tracking where those individuals fall within the overall group structure. This is why Sinha earlier proposed that primate social cognition operates through fundamentally ‘self-centred’ or ‘egoistical’ knowledge of their own position (Sinha, 1998). This makes computational sense. Calculating absolute positions across entire hierarchies demands considerably more mental resources than tracking relational differences, particularly when learning stems from personal experience. Self-referenced knowledge can be acquired through direct dyadic encounters, potentially bypassing the need to observe third-party interactions. This may constitute experience projection, like in the case of plovers, where one’s own experience is used to model that of others.
Elsewhere, Sinha has argued that macaques can integrate information about multiple companions simultaneously when navigating intricate social scenarios and that this provides us with evidence for their sophisticated forms of self-awareness, perhaps an “intermediate stage in the evolution of self-awareness” before human language-based self-awareness (Sinha, 2017, p. 567). Sinha notes that strategic calculations grew even more nuanced in cases where middle-ranking females withdrew as the gap between themselves and approaching dominants narrowed, even though their grooming partners held lower positions (Sinha, 1998). Pointing to his earlier work (in Sinha, 1996), Sinha identifies a distinctive pattern in one group of macaques where lower-ranked females became more desirable social partners, which is undocumented in other macaque communities. He notes that this explains why intermediate-ranked females sometimes retreated despite outranking their companions, since enhanced social desirability altered the strategic calculus. Sinha argues that high hierarchical standing can offset low social desirability, and vice versa. He interprets this as evidence that macaques weigh multiple, sometimes competing social dimensions when making split-second decisions “while confronting the pathological complexity that emerges from the social attributes of individual macaques, enmeshed as they are in complex societies of their own making” (Sinha, 2025, p. 4).
Sinha also emphasizes that macaque cognition extends beyond self-centred knowledge frameworks since evidence beyond this has been gathered in his later work (Sinha, 2003). He argues that we now have evidence for tactical deception which reveals that individual bonnet macaques can strategically “manipulating other individuals’ actions in various social situations that present them with significant pathological complexity” including deliberately sharing or concealing information they’ve personally obtained (Sinha, 2025, p. 4). Sinha even argues that certain individuals demonstrate recognition that companions hold divergent beliefs, subsequently executing deceptive manoeuvres to alter those beliefs and later exploiting the false beliefs they’ve deliberately cultivated. These sophisticated deceptive strategies represent responses to pathological complexity at its most cognitively demanding involving sophisticated theory of mind capacities. Sinha also suggests that the connections between this empirical work and the way their subjective experience is organized “could be mapped to the phenomenological complexity” profiles of these species in “functionally informative ways” (Sinha, 2025, p. 5). While I won’t attempt this here, let us now turn to how Sinha’s work could help us to understand the evolution of selfhood.
Social Life and the Evolution of Selfhood
Sinha asks what does this experientially derived self-knowledge reveal about self-awareness and selfhood concepts (see also Sinha, 2017)? While researchers once viewed selfhood as uniquely human, Sinha notes that recent scholarship has increasingly recognized biological continuity of self across species and traced its evolutionary history, citing work by Gilbert et al. (1995), Leary and Buttermore (2003), and Sedikides and Skowronski (2003). Drawing on Sedikides and Skowronski’s (1997) framework for understanding the symbolic self, that is, the “ability to both consider oneself as an object of one’s own reflection and to store the products of such” (Sinha, 2025, p. 5), Sinha focuses on their evolutionarily prior forms of self-awareness, that is, subjective self-awareness and objective self-awareness. While I only discussed the evolution of minimal self-awareness in my book, this gives me a good opportunity to relate the pathological complexity thesis to more complex forms of it.
Drawing on von Bertalanffy (1967), Sinha explains that subjective self-awareness involves an organism’s ability to distinguish itself from its “socioecological environment”, enabling self-regulation alongside the ability to “perceive, interpret, and, if necessary, change its environment through self-initiated action” (Sinha, 2025, p. 5). Sinha also notes that Sedikides and Skowronski (1997) emphasize that this level does not require stored mental representations of one’s own characteristics; it involves only rudimentary self-environment differentiation, which can be understood as equivalent to my discussion of minimal selfhood emerging out of sensory distinctions between internal and external information. While essential for survival, Sinha is right to note that this basic differentiation along with self-regulatory responses has typically been characterized as nonconscious (Damasio, 1994; Gallagher, 2011; Veit, 2023a). Sinha (2017) catalogues various capacities falling under subjective self-awareness: processing complex sensory information, mentally representing physical and social surroundings, numerical cognition, memory, categorization, communication with conspecifics and predators, tool use and creation, and problem-solving strategies, citing support from Gallistel (1989), Snowdon (1990), and Ristau (1991a). Yet Sinha emphasizes that Sedikides and Skowronski (1997) maintain these abilities need not involve self-representation and may operate effectively without it.
Sinha contrasts this with objective self-awareness, for which he cites several definitions: the capacity to “become the object of its own attention” (Gallup Jr., 1992, p. 117), awareness of one’s “own states of mind” (Cheney & Seyfarth, 1992, p. 144), and “to know it knows, to remember it remembers” (Lewis, 1992, p. 124). Sinha notes that Sedikides and Skowronski argue that this form of objective self-awareness indicates the existence of rudimentary self-representation, what they term the “objectified self” (Sedikides & Skowronski, 1997, p. 80). Sinha explains that objective self-awareness enables self-referential behaviour, where one uses one’s own knowledge as a template for modelling others’ knowledge, predicts companions’ actions by attributing motives, and shapes others’ behaviour through strategic intervention. Sedikides and Skowronski (1997) argue that species lacking objective self-awareness should not display these capacities and that among primates, only great apes and humans possess it. Citing Bargh (1984), Damasio (1994), Lewis (1992), and Sedikides and Skowronski (1997), Sinha argues that capacities objective self-awareness provides operate at a conscious level. Sinha notes that both he and I would tend to agree with this argument, though I will not that I do not think of consciousness as restricted to objective self-awareness. Many of the things falling under subjective self-awareness will also be experienced by conscious creatures, so the objective vs subjective terminology is unhelpful here, especially since ‘subjective’ evokes subjective experience.
While prevailing theorizing limits objective self-awareness to great apes, Sinha presents two arguments suggesting macaques developed an objectified self (Sedikides & Skowronski, 1997) earlier in evolutionary history. First, Sinha notes that macaques integrate hierarchical information about multiple interacting groupmates simultaneously, deploying these mental models during competitive scenarios (Sinha, 1998, 2017). This sophisticated ability, rare among nonhuman primates, suggests objective rather than merely subjective self-awareness.
Second, Sinha notes that when higher-ranking females withdrew despite dominance advantage because their partners were more socially desirable, they acted as though believing approaching females sought their companions rather than themselves. He argues that this retreat strategy rested on the principle that socially desirable individuals attract affiliative attention regardless of rank.
Furthermore, the tactical deception Sinha documented, sometimes involving novel strategies, potentially demonstrates second-order intentionality beyond what we have seen in the plover (Sinha, 2003, 2017), with Sinha suggesting that the deception macaques attempted to manipulate targets’ belief states, fits Dennett’s (1987) intentional stance. Sinha also notes that a motive attribution capacity suggests macaques form beliefs about others’ motivations representing what Sedikides and Skowronski (1997) identify as a defining feature of objective self-awareness, that is, allowing organisms to use self-knowledge for modelling other’s behaviour.
Sinha is right to emphasize that the pathological complexity thesis urges us to “consider both external sources of complexity and sources that arise endogenously from the individual’s subjective agency” (Sinha, 2025, p. 6). He observes that researchers are gaining clearer understanding of the evaluative and decision-making behavioural repertoires macaques deploy when confronting the social complexities of everyday life: I can thus easily see the phenomenological complexity of such decision-making as individual macaques confront the rapidly changing pathological complexities of their socioecological environments but I still need to understand how these evaluations and decisions can be potentially articulated in terms of Veit’s notions of other-than-human consciousness. (Sinha, 2025, p. 6)
Sinha finds my argument that consciousness evolved incrementally, with contemporary forms of consciousness in living species (macaques included) emerging primarily as adaptations for navigating the distinct dynamic ecological niches each species inhabits, to be compellingly self-evident. Sinha suggests this perspective aligns with arguments he and Gupta advanced earlier from a position of frustration (Gupta & Sinha, 2014), where they proposed potentially abandoning the consciousness concept altogether due to its apparent empirical impasse, advocating instead for concentration on the neurobiological executive capacities and functions that different taxonomic groups display within their respective socioecological contexts. Sinha characterizes this earlier proposal as representing “a modular manifestation of Veit’s phenomenological complexity thesis” (Sinha, 2025, p. 6). Sinha expresses particular enthusiasm for directing greater attention towards the “phenomenological complexity that different individuals within a species may harbour and express – clearly in response to the corresponding pathological complexity that they confront” (Sinha, 2025, p. 6). Sinha sees this individual variation in evaluative decision-making as potentially revealing differences in how conscious states are expressed across individuals.
Sinha notes my proposal that evaluative responses to pathological complexity may represent the fundamental function that initiated and guided consciousness evolution. He argues that socioecological life-history challenges macaques face created exploitable opportunities while demanding sophisticated evaluation, potentially driving emergence of conscious strategies for avoiding harmful outcomes. Throughout his essay, Sinha documented cognitive features qualifying as consciousness under my framework: sensory complexity, evaluative complexity, rudimentary objective selfhood, awareness of other selves, and behavioural deployment of integrated experiential knowledge. He proposes these capacities evolved to navigate external complexity characterizing macaque societies, shaped by internal phenomenological complexity, though acknowledges not previously labelling these as consciousness manifestations.
Sinha’s bonnet macaque research illuminates how pathological complexity drives not just minimal consciousness but increasingly sophisticated forms of self-awareness. The computational demands of navigating shifting social hierarchies, creates selection pressure favouring objective self-awareness and theory of mind. Unlike fixed dominance hierarchies where simple associative learning suffices, the variable and context-dependent social challenges macaques face reward capacities for mindreading and experience projection. This suggests that objective self-awareness evolved not as a cognitive luxury but as an efficient computational solution to specific forms of social drivers of pathological complexity. Just as hedonic valence provides common currency for comparing heterogeneous action values, objective self-awareness provides something similar to a common currency for social cognition, by enabling organisms to leverage experiential self-knowledge for modelling others’ minds. The pathological complexity framework thus predicts that objective self-awareness should evolve in species facing social environments where (i) successful action selection requires tracking multiple agents’ mental states simultaneously, (ii) optimal strategies vary based on one’s own position within dynamic social networks, and (iii) fitness consequences of social miscalculation are severe enough to justify cognitive investment costs. Sinha’s macaques exemplify precisely these conditions, suggesting the evolutionary threshold for objective self-awareness has been crossed in many other social species. If we find more evidence for these capacities in other socially complex species, this would provide further support for the pathological complexity thesis.
What Is It Like to Be an African Elephant?
African elephants have long fascinated humans due to their impressive size, complex social lives, and high intelligence, which suggest equally high levels of phenomenological complexity. This makes them an ideal case study for the pathological complexity framework. I am grateful to Don Ross, an economist and philosopher, who has spent a lot of time studying African bush elephants, for doing just that, offering us insights into what goes on in their minds by examining their peculiar life histories (Ross, 2025). Before I begin to examine his analysis, I will note that I am here focused on the African savanna (bush) elephant, Loxodonta africana, rather than the smaller forest elephant, Loxodonta cyclotis. Given their close phylogenetic relationship Ross could have decided to treat them together, but the ecological divergences between savanna and forest contexts counsel restraint in thinking that their subjective experiences will be the same. Unless otherwise indicated, ‘elephant’ refers to L. africana, and any extrapolation to L. cyclotis should be regarded as provisional.
Ross has shown a lot of enthusiasm for the pathological complexity thesis. His commentary does not aim to criticize it, but to draw on African elephants as a case study to demonstrate the value of my life history framework. His central claim is that my pathological complexity thesis clarifies why the most metabolically expensive brains in the metazoan lineage, that is, those of humans, cetaceans, and elephants, are relatively recent evolutionary products. Such energetically costly neural architectures become evolutionarily viable only under rising pathological complexity in the more complex life history strategies we find in us and them. This is a message we frequently encounter his recent book with Glenn W. Harrison The Gambling Animal, where they explicitly compare the life histories of humans and elephants, and use it to make inferences about the evolution of our respective minds. As they emphasize in their book, we shouldn’t treat intelligence as a default outcome of evolutionary processes: “Unusually expensive brains are, by definition, not recurrently naturally selected” (Harrison & Ross, 2025, p. 102). Unfortunately, this is something consciousness researchers frequently neglect, as if consciousness just came for free. In what follows, I will offer a detailed analysis of Ross’s exploration of the pathological and phenomenological complexity of L. africana and expand it to offer a tentative profile of their conscious experiences.
Elephant Life Histories
As Ross has argued forcefully over the last years (Harrison & Ross, 2025; Ross, 2019, 2022, 2023) an ecological life-history perspective makes elephants at least as informative a comparison class to humans as the great apes, such as orangutans, chimpanzees, and gorillas, despite their phylogenetic distance. He emphasizes that both elephants and hominins originated in Central and East Africa and were shaped by steppe environments and continuous changes in climate. Whereas orangutans, chimps, and gorillas live in forests, the environmental pressures that shaped elephants were the same as our own. These analogous ecologies, he suggests, selected for a suite of convergent traits: “long lives, intense socialisation, remarkably metabolically expensive brains, lengthy gestation and childhood, alloparenting, complex communication systems, self-awareness, tool use and modification, and advanced problem-solving capacities” (Ross, 2025, p. 2). Both lineages subsequently spread far across the globe from their African origins to occupy nearly every terrestrial biome, including “prairies, scrubland, forests, semi-deserts, wetlands, and tundra” (Ross, 2025, p. 2), even if this is now often forgotten due to the much smaller numbers of 450,000 elephants alive today that poorly reflect how dominant they once were across continents (Ritchie, 2024). Their comparable ecological flexibility is grounded in deeply social and cognitively demanding ways of life (Hedwig et al., 2021; Lee, 2014; Κ. Payne, 2003; Wittemyer et al., 2005) that makes them an excellent test case for a convergent evolution in consciousness.
However, as Ross stresses, there is a significant problem that we face in studying them. Empirical research on elephant minds, he notes, is still in its infancy and bounded by substantial methodological challenges. Based on his own experiments with African elephants over the last 5 years to assess their risk preferences (Harrison & Ross, 2025), Ross notes that the practical dangers of studying large, unpredictable, and dangerous wild elephants make it nearly impossible to conduct controlled experiments in natural habitats. Consequently, current knowledge of elephant behaviour and cognition is drawn primarily from comparatively tame individuals living under some degree of human control, such as in zoos or controlled semi-wild populations. But these conditions, as any ethologist readily recognizes, inevitably limit how far findings can be generalized to fully wild populations, which is where the pathological complexity thesis aims to understand the evolutionary pressures that shaped the minds of animals. Indeed, Ross draws on reviews of elephant cognition by Byrne and Bates (2011) to emphasize that this problem is especially pronounced in elephant populations. In their natural habitats, adult elephants must constantly make high-stakes foraging decisions, and “successful track records in such decisions are the basis for group acknowledgment of authority in leaders of matriarchal herds” (Ross, 2025, p. 3). Captive individuals, by contrast, inhabit an artificial stability and the inherent risk that shapes cognition in the wild is largely absent.
Ross suggests that such conditions inevitably distort the cognitive profile of captive elephants, much as a study of human reasoning confined to those “too wealthy to ever have to earn income or subject themselves to hierarchical control after childhood” could yield misleading conclusions about ourselves (Ross, 2025, p. 3). To address this, Ross’s group seeks to design experiments for managed elephants without human contact and could eventually be adapted for use with wild populations, though he concedes that achieving this remains a long-term goal. I will note that these methodological challenges are not just an unfortunate limitation. If consciousness evolved to deal with the pathological complexity challenges unique to each species, then captive and regulated environments eliminate much of this complexity, making it significantly more difficult to understand the functions of their mental capacities.
Luckily not all hope is lost. As Ross points out, systematic knowledge of African elephant life history has been built primarily through decades of field observation rather than experimental study. The richest source of such data is the Amboseli Elephant Research Project (AERP), which, since the early 1970s, has tracked identifiable individuals across generations within a protected and ecologically diverse park in Kenya. A 2001 report, for instance, noted that “data on life histories and association patterns have been obtained for more than 1700 individual elephants over 28 years” (McComb et al., 2001, p. 492), and much more data has been gathered since (C. J. Moss et al., 2019). As Ross points out, because elephants are large, conspicuous, and generally unthreatened by observers in vehicles, researchers have been able to document their behaviour with unusual continuity and precision. Furthermore, Ross notes that this data has even led to projection matrix models to describe their life histories (Croze & Lindsay, 2011), which is the approach favoured in my recent collaboration with biologists at Oxford to measure pathological complexity (Veit et al., 2025). But there is also much more data that hasn’t made it into scientific publications. As Ross notes, a wealth of anecdotes is curated on the website of ElephantVoices, a non-profit organization dedicated to research and advocacy for elephants. Like Ross, I maintain that: “at sufficient critical mass, given professional-quality reliability of observers, such anecdotes constitute scientific evidence” (Ross, 2025, p. 3; see also Browning, 2017). They even offer a detailed elephant ethogram cataloguing and detailing their behaviours that can serve as a reference point for anyone studying them. 2 An article in the Scientific American describes this ethogram as “Google Translate for elephants” (Nuwer, 2021). For an ethological approach to animal consciousness this puts us in an ideal position, since I describe ethograms in my book as a precursor to detailed life histories. While Ross qualifies that his analysis operates through the lens of someone who has spent many hours with elephants, this is precisely to how I have argued that the ethologists like Konrad Lorenz enabled a revolution in our understanding of animal behaviour (Veit, 2023a). We can do the same for animal consciousness.
Ross identifies the central pathological complexity challenge shaping every aspect of elephant life history as the “challenge of maintaining very large bodies, with extremely metabolically expensive brains, in relatively arid savannah and scrubland” (Ross, 2025, p. 3). Elephant digestion, he notes, is inefficient compared to that of most large herbivores, and their typical diet is nutritionally poor. Ross draws by a review Hart et al. (2008) who have argued that this forces elephants to spend wild elephants devote “60–80% of each 24-h day obtaining this low quality forage (Owen-Smith, 1988). For elephants, meeting foraging and water requirements does not involve coordinated, fine-grained cognitive responses but rather an ability to reliably locate scattered, distant foraging and water resources” (Hart et al., 2008, p. 87). Elephants have not only the largest brains, but also the largest cerebral cortex, of all terrestrial mammals (Hart et al., 2008, p. 87). Ross points out that they remain near the top of the metazoan kingdom even adjusting for encephalization quotient, which quantifies the expected brain size given their body size, and has marked a unique place for our own species in the animal kingdom (Deaner et al., 2007; Herculano-Houzel, 2016; Williams, 2002).
To understand how such energetically demanding neural machinery could have been favoured by selection, Ross reviews the distinctive features of elephant neuroanatomy. He frames this explicitly trough the pathological complexity framework, noting that the high metabolic cost of such brains must have brought about significant fitness effects in their life histories. Elephant brains, Ross explains, exhibit a structural design found in no other mammal: their cerebellum contains an extraordinary proliferation of “densely folded cerebellar neurons, ten times the number occurring in any other terrestrial mammal” (Ross, 2025, p. 3).
3
Because the cerebellum is generally associated with fine motor control and spatial coordination, Ross cites Maseko et al. (2013) as a prominent discussion of the idea that this folding evolved to manage the elephant’s trunk, an organ unmatched in its versatility. Yet, Ross argues that, given the neural plasticity and general computational potential of cerebellar tissue, it is improbable that this brain region does not do more. He suggests that the enlarged cerebellum has likely been co-opted to support elephants’ more general intelligence. This is in line with more general discussions on the connection between high intelligence and social complexity, which Ross argues has been well-established in the work of Dunbar and Sutcliffe (2012). As Ross notes, the social and ecological pressures are inseparable in elephant herds: [T]he ‘push’ and ‘pull’ aspects of the selection of their expensive brains are likely closely entangled: foraging complexity favoured expensive brains, but expensive brains increased foraging demands. The social structure of their life history is likely best explained as the mechanism for achieving equilibrium in these evolutionary dynamics. Because an elephant’s required portfolio of nutrients requires typically widely disbursed ranges of foods, elephants cannot forage opportunistically – that is, simply travelling semi-randomly within range of water sources and eating what they happen to find. (Ross, 2025, p. 4)
Ross proposes that elephants’ intricate social arrangements evolved as a stabilizing mechanism for balancing these competing pressures, involving careful planning for long-distance movements between food and water sources. Drawing on decades of evidence from the AERP, Ross points to the central role of leadership within both matriarchal family groups and bachelor herds, where experienced individuals guide collective decision-making. Ross suggests that adult sex-based grouping in elephants is an adaptive partitioning of the foraging niche: by keeping mature males separate from maternal units, the species minimizes direct competition over food. In this arrangement, bulls can range widely and move quickly across sparse resources, whereas mothers with calves can prioritize slower, selective, risk-averse feeding strategies suited to developing young. Selection, on his view, would have favoured this social segregation because it reduces interference between large males and growing offspring and thereby improving overall resource use. Again, from the perspective of the pathological complexity thesis this makes a lot of sense. As I have argued elsewhere, sex-specific life-history strategies are common to maximize reproductive output and they can even help us to understand differences in the mental states of sexes (Veit & Browning, 2022).
Ross also applies these ideas to the question of elephants’ exceptional memory. Drawing on his experimental work with six African elephants, he observes that the animals learned risky quantitative choices more quickly when they were allowed to keep walking. They also showed a strong aversion to standing still, even when doing so yielded an immediate food reward (Harrison & Ross, 2025). Ross interprets this pattern as evidence of a close link between movement and learning, similar to how humans learn skills through demonstration rather than texts. Ross even argues that their remarkable long-term memory might itself be embodied. The species’ enlarged cerebellum, he proposes, could function as a dense proprioceptive index, allowing specific experiences to be recalled through characteristic sensorimotor states, making it a large-scale analogue of human muscle memory. I have much sympathy for this hypothesis since it would suggest that bodily coordination is not incidental to cognition but an integral part of it, which is precisely what the pathological complexity thesis would predict. The fact that elephants have evolved to maintain a vast range of memories is suggestive that they deal with highly multi-dimensional trade-offs in their decision-making.
Ross also argues that modelling elephant consciousness should not treat their consciousness as an inferior version of our own because they lack external records that played such an important role in the rise of human civilization. On his view, humans rely on such scaffolds to counter two liabilities linked to our frontal-cortex-heavy cognition: (i) the natural decay of individual memory and (ii) the tendency of shared imagination to decouple collective models from reality unless anchored by common external references. But this is not a problem shared by elephants: Elephants, with highly stable individual memories rooted in their cerebellar layers, and less active imaginations, were under no similar selection pressure. Emphasis on this difference very exactly expresses Veit’s pathological complexity thesis: humans, but not elephants, are under continuous threat of collective insanity, and this should be expected to result in differences in their respective forms of consciousness. Avoidance of anthropocentrism in Veit’s sense here consists in treating the importance of constructed semantic scaffolding in human consciousness as irrelevant to modelling elephant consciousness. The evolution of elephant consciousness is not a train on a trip to human consciousness that stopped a station or two short of the final destination. On the other hand, these two evolutionary trajectories might have run along more closely parallel tracks than those followed by any other kinds of animal, and this might be of profound importance to future practical efforts to build more cooperative interspecific relationships outside zones of captivity. (Ross, 2025, p. 6)
Ross maintains that elephants may possess subjective experiences that are not only just as rich as our own, but also can help us to understand our own minds better than even our closest Great apes relatives. A convergent evolution in human and elephant consciousness would help us to make inferences about which pathological complexities shaped human consciousness. Ross uses the example of chimpanzees who show a lack of interest in sharing subjective perspectives, which might indicate this capacity was absent in the last common ancestor, but he notes that “we must allow for the possibility that chimps lost this disposition, or even capacity, if its value for their fitness declined after the point of separation from hominids” while the “shared ecological history of two distantly related species, such as humans and elephants” can provide “a stronger basis for inferences from features of pathological complexity” (Ross, 2025, p. 6). I share Ross’ hope that a recognition of this parallel evolution of phenomenological complexity in the sparse environments of the African steppe will allow us to enable better interspecies communication and relationships that recognize their rich and unique subjective experiences.
To understand the subjective experiences of elephants we should, of course, turn closer attention to their communication with each other, which Ross spends the most time in his commentary on, reflecting his previous work on elephant language (Ross, 2019). Indeed, Ross further suggests that my program could potentially revolutionize our attempts to communicate with them by providing AIs with detailed data about elephant life histories. He highlights findings by O’Connell-Rodwell et al. (2024), Mutinda et al. (2011), and K. Payne (1998) that specific acoustic patterns are used to signal and negotiate changes in direction, and speculates that this communication evolved to deal with the pathological complexity challenges in their foraging strategies. He argues that future decoding of elephants’ sub-sonic rumbles with modern machine-learning tools could test whether elephants “consciously manipulate somatic memories to an extent that humans at least generally do not try to do” such that they might be able to “describe, to themselves and others, techniques for such evident skills as testing air currents for learned indicators of distant rainfall” (Ross, 2025, p. 4). Ross emphasizes that this possibility hinges on what is ultimately discovered about the nature of elephant language, though he is suggestive that we may have underestimated its complexity: Close-quarter elephant communications through rumbling are accompanied by ear movements, head position adjustments, and trunk touching that also appear to have signalling properties. If such signals modulate rumble semantics – a possibility about which, as far as I am aware, nothing is yet known – then this may complicate efforts to decode rumbles using only acoustic recordings provided as data to deep-learning decoder systems. The AI decoders might also need access to video records. At longer distances, elephants exchange information through seismic signals detected by sense organs in their footpads. There is suggestive evidence that such communication is used to relay information about locations of water and forage, but of course ranges of other possible uses are possible, which await systematic investigation. (Ross, 2025, p. 5)
Groups of elephants, Ross notes, produce near-continuous rumbles at frequencies we cannot detect. These signals, he argues, bear community-specific structure and are sensitive to small differences. As Ross points out, in mixed groups of elephants assembled from distant populations in captivity rumbling rates drop markedly, which he interprets as consistent with limited cross-dialect comprehension.
Methodologically, Ross is sceptical that human-style codebreaking will suffice. Approaches modelled on deciphering human symbol systems, he suggests, are likely to stall because people import their own anthropocentric perspectives. This is why he thinks that AI is more promising in its search for statistical regularities without strong priors about topic or grammar. Drawing on the ground-breaking work of Pardo et al. (2024), who have discovered name-like calls between animals, as well as earlier findings of the meaning of deliberation rumbles about foraging decisions, Ross judges it credible that researchers will soon conduct two-way exchanges using “sub-sonic microphones and acoustic synthesisers” (Ross, 2025, p. 5).
I agree with Ross that if humans could engage elephants in interactive exchanges we might infer far more about the structure and content of their consciousness than passive observation allows. Ross explicitly embraces the idea that elephants may possess language, arguing that the persistent effort in linguistics and philosophy to define language as uniquely human is untenable. Evolution, he notes, rarely draws sharp boundaries, and expecting one here is likewise misguided. Yet, he maintains that humans remain distinctive in producing durable, externally stored records of communication, which enabled the rise of human civilization.
Ross also cautions that even if elephant communication is decoded, wild individuals may not be motivated to converse with humans, while captive ones may respond in artificial ways. Still, once genuine elephant-to-elephant exchanges are understood, such limitations could be empirically tested. Finally, he issues an ethical warning: interspecies dialogue will require strict safeguards, since being addressed by another species could alter elephant consciousness itself. And once humans can speak with elephants, he warns, they will also be able to deceive them which is a prospect that demands serious moral consideration.
Unfortunately, this is a problem that animal ethicists have so far given little attention and it is one I hope to address in future research. Indeed, it is an especially pressing one given the evaluative richness we expect to find in their conscious experiences. As Ross’s review of Sukumar (2003), Moss and Poole (1983), and McComb et al. (2001) shows, elephants can sometimes create large social gatherings of “hundreds of individuals who mutually recognise one another” (Ross, 2025, p. 5). Ross also notes that elephants unlike most other animals seem incredibly interested in diseased individuals, even returning to the places where they died. Drawing on Moss and Lee (2011), Ross even argues that friendships among elephants go beyond their own groups, showing special interest in introducing themselves to newly born calves of other groups, and seem to play a role in decisions of who becomes the new matriarch. Ross even points to the curious finding reported in Sheldrick (2012) that “orphan elephants rescued and fostered by people and then reintroduced into wild lives recurrently journey with their new offspring to present them to their former caregivers” (Ross, 2025, p. 5). Ross also points to the rich empirical literature of norms in elephants, who stand out among other animals (Andrews et al., 2024), which suggests a high degree of evaluative complexity. In humans, culture vastly increases the pathological complexity our species has to deal with. The same should be expected in elephants, thus increasing their mental complexity to deal with culture.
Following this detailed analysis of Ross’s discussion, I would now like to provide a comparative analysis of how we may use the pathological complexity thesis to compare human and elephant consciousness (see Figure 1). A comparative consciousness profile for the phenomenological complexity of humans and elephants [inspired by Birch et al. (2020, Figure 1, p. 791) suggestion for how to construct consciousness profiles].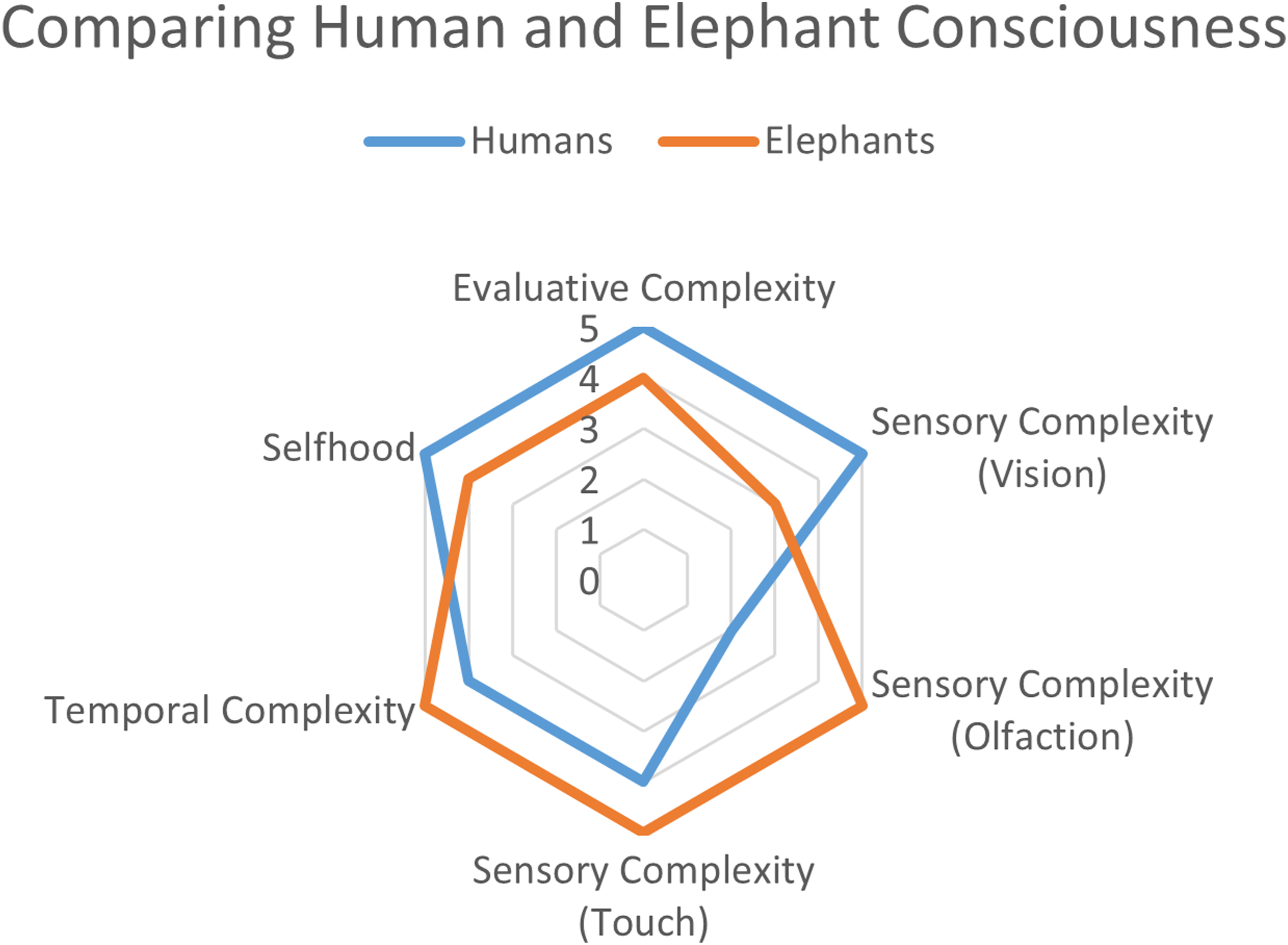
Dimensions of Elephant Consciousness
In their Dimensions of Animal Consciousness article that influenced the multi-dimensional approach in my book, Birch et al. (2020) offered a hypothetical consciousness profile comparing elephants, corvids, and cephalopods. This was a bit surprising since the paper almost exclusively discussed evidence from the latter two groups, though they admit that their profile is meant to be entirely speculative. In Figure 1, I hope to have provided our current best but nevertheless speculative comparative assessment between elephants and humans.
Evaluative Complexity
Both species face intense pathological complexity driven by expensive brains, challenging environments, cultural norms, and complex decision-making involving long time horizons. I position humans slightly higher because our evaluative capacities have become intertwined with abstract economic evaluations using monetary currencies. Elephant evaluation, while rich, lacks this economic scaffolding, which significantly increased the pathological complexity our own species has to deal with. However, I admit that there is great uncertainty here. Because humans have externalized evaluative information through records and numerical systems, we may have actually reduced our reliance on hedonic experience, potentially making our evaluative experiences less rich than that of elephants.
Sensory Complexity
Humans score substantially higher on visual sensory complexity. Human experience is dominated by vision, with our richer colour perception that is trichromatic rather than dichromatic, fine-grained spatial acuity, and visual-dominated attention, especially in regard to other’s faces. For elephants, vision seems to occupy a comparatively secondary position in their sensory hierarchy, which sounds, smells, and touch playing more important roles. In the domain of tactile experiences, elephants seem to surpass humans again. Even Asian elephants, despite their smaller size, have incredibly complex muscle and nerve arrangements that take up a large part of the brain that our smaller skulls couldn’t accommodate (Purkart et al., 2022), thus likely providing tactile information of a richness we struggle to imagine. Combined with their potential reliance on ground-borne seismic vibrations detected through footpads (Mortimer et al., 2018), elephants appear to inhabit a tactile phenomenology where mechanical and vibrational information provides a primary perceptual framework. Human touch, while important, occupies a far more limited role in our conscious experience, though we excel in the usage of our hands. Elephants also vastly exceed humans in olfactory complexity. Their highly developed olfactory capacities provide crucial information about distant water sources, individual identification, reproductive states, and environmental conditions. The empirical data suggests a chemical sensory world far richer than anything humans can experience. Human olfaction, by contrast, plays a comparatively much reduced role in our phenomenology and life histories. While I didn’t include auditory complexity in the figure to reduce visual cluttering, elephants would likely surpass us even here, detecting frequencies we can’t.
Temporal Complexity
Elephants appear to have much richer temporal experiences than our own species. While humans possess rich memories compared to most species our reliance on external records may have reduced selection pressures in our life histories for robust endogenous memory, potentially degrading our natural capacities since this cultural innovation emerged. Elephants, lacking external scaffolds, maintain highly stable memories. Elephants likely engage in mental time travel extending decades, enabling matriarchs to draw on extensive experience when making group decisions. This is a unique adaptive response to the pathological complexity of navigating territories where resource locations and migration routes must be remembered across decades to provide their expensive brains with energy.
Selfhood
Both humans and elephants possess self-awareness and engage in complex cultured societies. However, I score humans higher on selfhood not because elephant subjectivity is less robust, their identity across time is likely more stable, but because human identity involves an additional layer of constructed narrative, as well as richer language, and the need to avoid exploitation. As Ross argues, humans faced unique selection pressure to anchor shared imaginations in external references to prevent what he aptly described as insanity, that is, the tendency of our brains to decouple from reality. Elephant selfhood is differently structured: deeply embodied and socially embedded, but lacking the reflexive narrative layer that characterizes human self-consciousness.
To conclude, elephants provide a beautiful case study of the pathological complexity thesis, showing how we can move beyond a human-centric approach to consciousness that treats other animals as only having lesser versions of our capacities. In many ways, elephants can justly be described as more conscious than our own species.
Plants
Chincarini suggests that the pathological complexity thesis could potentially be criticized at the level of plants (Chincarini, 2025). This is why I invited Özlem Yilmaz, which initially surprised her, because of her unusual combination of expertise: she holds doctorates in both plant biology and philosophy. Although my book deals exclusively with animal consciousness, and I argue there against attributing consciousness to plants, or for that matter only a very small probability (Terrill & Veit, 2024), I was keen to hear how someone deeply versed in plant biology would evaluate the framework due to the rising interest in plant cognition and sentience.
Yilmaz sees considerable promise in the pathological complexity thesis, but proposes that the concept of ‘stress’ might serve as an equally illuminating construct, given its tight links to the life-history trade-offs that lie at the centre of my account. Even if a careful look at the distinctive pressures shaping plant lives does not push us towards attributing consciousness to them, she emphasizes that their forms of cognition remain scientifically and philosophically instructive.
Plant Cognition? Yes. Consciousness? No
Yilmaz is right to note that the expanding discussions of non-human consciousness have not only focused on other animals. While comparatively much smaller, there are now serious discussions of plant intelligence and the possibility of their consciousness (Calvo, 2017; Calvo et al., 2017, 2020; Garzón & Keijzer, 2009; Hansen, 2024; Mallatt et al., 2021; Segundo-Ortin & Calvo, 2023; Sims & Yilmaz, 2023; Taiz et al., 2019, 2020; Trewavas, 2003, 2004; Trewavas et al., 2020). Like in the pathological complexity framework, Yilmaz argues that cognition should be understood in the context of the organism as a whole and in its environment. Yilmaz argues that the concept of the organism is essential for understanding cognition, which is fundamental characteristic of living beings since they are self-regulating and cognitive systems that continuously engage with their surroundings. I am happy to concur with her that they actively perceive their environment to regulate and maintain internal processes, while also impacting the environment itself, as illustrated by examples like plants altering soil chemistry. This constant organism-environment interaction is fundamental to life and involves at least quasi-cognitive capacities across all organisms, down to the level of single-celled bacteria. Thus, Yilmaz suggests that internal coordination/organization as well as the external environment are fundamental components of biological cognition.
However, while Yilmaz notes that it is important to maintain a distinction between cognition and consciousness for scientific and ethical purposes, she does seem to hold two positions that are not easy together.
At first, Yilmaz argues that she avoided doing animal experiments during her education in order not to inflict them with pain, and was happy when she was doing plant work instead. She argues that plant research also involves ethics and that “we should be unflinching defenders of plants in forests, steppes, wetlands, cities, and beyond, I think that it is important not to confuse these efforts with animal welfare efforts that are connected to the conscious experiences of animals” (Yilmaz, 2024, p. 2). She emphasizes that we should avoid using zoocentric concepts in plant research which is suggestive that she wants to avoid using terms like consciousness for plants, since she also approvingly quotes Brooks Pribac who argues that “comparisons to animals used without adequate caution can have serious ethical implications, not only because they may put scientific quality at risk (Struik, 2023) but also because they can trivialize animal sentience and jeopardize efforts to expose and eliminate the ubiquitous violence inflicted upon nonhuman animals” (Brooks Pribac, 2023, p. 1). But then Yilmaz surprisingly argues that we should not think of plants as less conscious than animals: Even though plants do not have consciousness in the ways that most animals do, it does not mean that they are “less.” The ways plants experience the world are simply different than how animals experience it. […] Perhaps, plants have a specific kind of capacity, that we do not know of yet. Such a capacity would be a plant version of consciousness that would entail their unique ways of interacting with the environment. And, regardless, even if plants do not have consciousness, that does not mean that they do not have cognitive capacities and highly complex ways of interaction with their environments, and adaptive behavior. So, it should not be said that plants are similar to those animals that have the “least consciousness.” (Yilmaz, 2024, p. 2)
Similarly, Yilmaz argues that treating plants as less cognitive or intelligent is a mistake reflecting an unwillingness to grant plants the status of an organism. But I think she goes too far in the opposite direction. Yilmaz appears to be attracted to the organizational approach that is close to the autopoietic tradition. Here, agency and cognition are largely treated as a definitional matter of all life. But that is a failure to recognize that things can come in degrees. Moving away from a binary view in which only some entities have consciousness, agency, or intelligence, towards one that grants these properties to all life, involves an equal aversion to gradualist thinking. But a deeply gradualist evolutionary view does not appear to suffer from the human-centric biases that deny plant capacities. Plants have a unique evolutionary history and very different life-history strategies that allow us to understand plant agency and cognition as real and important. While our animal-centric perspective may still bias us, we shouldn’t deny that some organisms are more agentic, cognitive, and conscious than others. These are all multidimensional capacities that can come in degrees and are important for ethical decision-making (Terrill & Veit, 2024).
Yilmaz does not offer a positive case for plant consciousness. She notes that plants lack centralized nervous systems and neurons for that matter, instead relying on a distributed phloem and xylem network for organismal organization, control, and communication, with a modular design. But it is this organization that makes consciousness unnecessary even when plants are dealing with high pathological complexity. As I’ve argued in several talks over the years, plants should be understood similarly to a collective of humans in a company. While we can usefully and accurately describe businesses as agent-like, we do not think they have a unified consciousness. Plants do not deal with action-selection problems in the same way as animals where the actions have to compete against each other. Rather, each cell is engaged in independent and parallel decisions that may be influenced by information from other cells, just like a group of coordinating humans. Thus, there is no useful role here for consciousness. This becomes even more apparent when we turn to the concept of stress, which Yilmaz proposes as a supplemental concept to pathological complexity.
Stress and Pathological Complexity
She points out that the stress research “in both plant and animal science involves concepts such as avoidance, sensitivity, resistance, tolerance, and resilience” (Yilmaz, 2024, p. 3). However, she never defines the term, which is a problem since there is no real agreement across literatures here. As reviews of the concept are quick to point out, stress research “continues to be challenged with the problem of operationalizing the concept: The ways in which the term stress is used in research is almost as subjective as an individual’s experience of a stress” (Robinson, 2018). Yilmaz is attracted to the organizational tradition and so I suspect that she views stress as a state that challenge homeostasis, but such a generalized view makes it unclear of whether it captures any unified phenomenon since on such a broad view almost all biological processes may be seen as dealing with homeostasis in some way. I say almost because, as I argue in my book, the ultimate goal of organisms is of course not maintenance but reproduction. Organisms only aim to restore homeostatic balance in order to and to the extent to which this is contributing to pursuit of their life history strategy. One may thus treat it as a competitor concept to my concept of pathological complexity, but of course I don’t deny that the mechanisms we often describe as dealing with stress are real. They may well be redescribed as dealing with pathological complexity, or alternatively we may redefine stress in a Darwinian sense as pathological complexity challenges to the life history strategies of organisms. That would give us a better way of distinguishing between those states that trigger significant organismal responses and those that do not, which the interdisciplinary field of stress research desperately needs. But my goal here is not to develop this view of stress further. I will only say that Yilmaz is right that stress research is of significant relevance to the pathological complexity framework, though not because I think it is useful to consciousness research (with the exception of psychological stressors in particular), but because it has identified many of the mechanisms organisms have evolved in order to deal with their pathological complexity challenges, regardless of whether they are “genetic, epigenetic, physiological, and developmental processes” (Yilmaz, 2024, p. 3). I will aim to return to this in future work on how the pathological complexity framework can help us to Darwinize our understanding of health and pathology, but that is not my goal here.
Footnotes
Acknowledgements
I am thankful for the detailed examinations of my work by the contributors to this special issue. They have demonstrated the usefulness of my framework and helped me to expand it further. Lastly, I thank the editors of the journal Tom Froese and Lola Cañamero for enabling this special issue.
Funding
The author acknowledges funding from Open Philanthropy.
Declaration of Conflicting Interests
The author declares no conflicts of interest related to this article.