
Abstract
The contents of consciousness are complex and dynamic and are embedded in perception and cognition. The study of consciousness and subjective experience has been central to philosophy for centuries. However, despite its relevance for understanding cognition and behaviour, the empirical study of consciousness is relatively new, embroiled by the seemingly opposing subjective and objective sources of data. Francisco Varela (1946–2001) pioneered the empirical study of consciousness by developing novel, naturalised and rich approaches in a non-reductive and comprehensive manner. In this article, we review the main conceptual distinctions and philosophical challenges of consciousness research and highlight the main contributions of Varela and his associates: the development of neurophenomenology as a methodological framework that builds a bridge between subjective and objective sources of data and the discovery of gamma-band phase synchronisation as a neural marker of perceptual awareness. Finally, we describe the work of Varela on time consciousness, his philosophical approach and the implementation of his neurophenomenological framework for its study by integrating subjective reports with neural measures.
1. Introduction
Humans are unique in the momentary and lifelong collection of their contents of consciousness, or subjective experiences, which include – but are not limited to – sensations, feelings, and thoughts. Typically, the contents of consciousness are complex and dynamic, with an extremely wide range of possible experiences that continuously adapt to internal and external stimuli as well as to other experiences. While in standard social situations, we are relatively successful at guessing subjective experiences of other individuals by drawing inferences from their behaviour and affective display (Ames, 2005), it proves to be very difficult to measure the contents of consciousness with scientific methods.
The study of consciousness and its contents typically distinguishes between sensory processing and subjective experience – for example, correctly recognising a colour and having an experience of that colour are not the same 1 (Block, 2011; Fazekas & Overgaard, 2018; Overgaard, 2018). The problem of the detection of consciousness can be addressed with objective methods (i.e. third-person approaches) such as electroencephalography (EEG) and psychophysics (e.g. by measuring performance with a visual detection task alongside participants’ neural correlates), with subjective methods (i.e. first- and second-person approaches), such as analysis of verbal reports and questionnaires (e.g. by asking participants about different aspects of their subjective experience), or with a combination of both types of methods. For instance, analysis of neurophysiological data can distinguish which of two competing images participants perceive when each of them is presented to different eyes (binocular rivalry paradigm; Haynes & Rees, 2005), or to evaluate if an unresponsive vegetative-state patient follows mental imagery-related instructions (Cruse et al., 2011). More recently, neural information analyses of ambiguous images and sounds (i.e. bistable perception) can distinguish the specific contents experienced by the participants not only when contents are explicitly reported (Canales-Johnson et al., 2020b) but also when they are inferred without explicit report (Canales-Johnson et al., 2021a). Machine-learning-based analysis of functional magnetic resonance imaging (fMRI) data can even enable prediction above chance level of the visual contents of non-rapid eye movement (NREM) sleep dreaming (Horikawa et al., 2013). Yet, despite the invaluable progress, studies using objective methods are limited by the predictability of the contents of consciousness, that is, they rely on the recognition of neural activity patterns that emerge in association with the standardised contents of consciousness. For instance, in the same study mentioned above, dream content detection relied on the neural similarity between relatively simple perceptual experiences in waking and sleep onset, which may not hold for more intense and unpredictable rapid eye movement (REM) sleep dreaming or other unresponsive states of consciousness with complex spontaneously developing subjective experiences. Furthermore, neuroimaging-driven predictions are not explanations, while the ultimate goal of the science of consciousness is the explanation of how subjective experiences arise from brain activity.
Compared to objective methods, content analysis of subjective reports enables a very detailed assessment of naturally and spontaneously occurring subjective experiences, including – but not limited to – such diverse contents of consciousness as self and body, visual experiences, perception of sound, temporal and spatial distortions, etc. Arguably, a detailed mapping of human phenomenology in different states of consciousness will eventually provide a solid ground for research into the neural basis of consciousness. While subjective methods alone are not able to provide sufficient insight into the neural mechanisms of consciousness, a combination of both objective and subjective methods will probably become one of the most promising avenues of consciousness research in the near future. Nevertheless, it is not straightforward to combine these methods, and they are still being developed largely separately.
Francisco Varela (1946–2001) was a Chilean polymath whose work spanned from biology, immunology and neuroscience to experimental psychology, philosophy of mind and phenomenology; he addressed these matters with scientific rigour while holding great regard for the philosophical concerns about human experience. Among his many contributions, we find neurophenomenology, a research approach that attempts to build a bridge between objective and subjective information, that is, an integrative method to study the hard problem of consciousness (see section 4.1); the study of neural oscillations such as phase synchronisation (see section 4.2) as a testable mechanism of perceptual awareness, and the experience of time in the flow of consciousness (see section 4.3). Here, we review the main concepts of consciousness research, the main philosophical challenges involved in the study of consciousness, and the contribution of Francisco Varela to the field.
2. Consciousness and its hard problem
Glossary. Definitions of consciousness

James (1890) argued that there is continuity in the diversity of phenomenal experiences, which can be conceptualised as the stream of consciousness, pointing to the constantly changing, yet coherent and continuous, contents of phenomenal consciousness. Continuity of the stream of consciousness depends on the temporal and spatial unity or binding of individual subjective experiences (Dainton, 2005; Revonsuo, 2009). Even though each distinct experience may occur at different points in the phenomenal space and time, they are joined into a coherent flow of a single unified stream of consciousness. That is, we do not have an experience of unrelated sensations that jump in time and space, although this may take place in certain psychiatric or neurological conditions such as schizophrenia (Vogeley & Kupke, 2007) or akinetopsia (Zeki, 1991), but an experience of spatially and temporally bound qualities that form a single stream of consciousness.
Phenomenal contents of consciousness are subjective and directly available only to who experiences them. Therefore, explaining the ontology of consciousness poses a unique problem for science: it violates the universal assumption that the phenomena studied by science should be equally available to all observers, and no observer should enjoy privileged access to those phenomena. Even though representational contents of consciousness can be successfully mapped using neuroimaging techniques, specific blood-flow or electrophysiological activation patterns do not seem to provide access to subjective experiential qualities. Consequently, there seems to be an unbridgeable explanatory gap between the objective neurophysiological data and the subjective contents of consciousness; that is, even if we are certain that the existence of consciousness is dependent on the brain, we cannot understand how brain activity causes or constitutes consciousness (Levine, 1983; McGinn, 1989). Reductive analyses of consciousness, that is, suggestions that consciousness is identical with some types of brain processes or functional descriptions, always seem to be logically consistent with the absence of phenomenal subjective experience despite the presence of these processes (Chalmers, 1997; Nagel, 1974). These intuitions have been illustrated by several famous thought-experiments (Jackson, 1986; Nagel, 1974). Nagel (1974) argued that even if objective science would acquire all physical facts about humans or other mammals, we would still miss information about their subjective point of view. For instance, even if we would know all neurophysiological and behavioural facts about a bat, including its sensory echolocation system, we would still have no idea of ‘what is it like to be a bat’, in other words, what are its subjective experiences of flying in complete darkness among trees. Likewise, the same argument can extend to other conscious organisms, including humans: their subjective point of view is always left out in an objective scientific description of behaviour and neurophysiology (Nagel, 1974). Jackson (1986) argued that a neuroscientist who is congenitally colour blind but knows all objective facts about the psychophysics and physiology of colour perception would still learn something new if one day her colour vision was cured, which seems to indicate that there might be more to consciousness than physical facts about its mechanisms.
Many of the consciousness-related research problems, such as the ability of a system to access its own internal states or the difference between wakefulness and sleep, seem to be relatively easy, that is, they seem to be directly susceptible to the standard methods of cognitive neuroscience (Chalmers, 1995). Even though some of the easy problems of consciousness are not yet solved, we have a clear idea of what would be their scientific explanation – for example, the discovery and description of the physiological correlates of consciousness should shed light on their solutions. Contrary to this, the problem of the material basis of conscious experience is a problem of a different nature: it is a hard problem, which seems to resist any of the empirical methods available to contemporary science (Chalmers, 1995). We do not have any idea of how and why subjective sensations or emotions arise from a physical basis and yet they do. Even worse, we cannot even imagine how this problem could be solved in terms of neurophysiological or neurochemical processes.
The hard problem can be tackled by a number of different philosophical theories of the mind–body relationship, and some of these theories do not seem to have the hard problem of consciousness at all. For instance, panpsychism – the assumption that phenomenal contents are basic to all matter – does not (Dennett, 2018; Goff, 2009; Seth, 2021). Arguably, the hard problem applies mainly to materialistic theories that aim to explain subjectivity in terms of objective physical or biological processes.
3. The mind–body problem
3.1 Classical solutions to the mind–body problem
Throughout centuries of theoretical research, controversies between dualism versus monism, idealism versus realism, and rationalism versus empiricism, placed the mind and consciousness at the top of philosophical investigations (Crane & Patterson, 2000; Heinämaa et al., 2007). Dualistic approaches to the mind–body problem assume that the mind (or certain aspects of it) and the physical world are ontologically separate and cannot be reduced to each other. Proponents of substance dualism, most notably Descartes (1641), suggested that consciousness and the brain are different substances, which can exist independently of each other. Property dualists supposed the existence of one substance with two fundamentally different aspects, that is, mentality and materiality, where mental properties do not exist in the absence of physical properties (e.g. Mill, 1974). Contrary to dualistic positions, monism assumed the existence of only one independent substance. For instance, La Mettrie (1748) defended a materialistic position (also called physical monism), arguing that consciousness has purely physical causes and there is no necessity to assume the existence of a separate ‘soul-substance’. On the opposite extreme of monistic positions lied idealism (also called mental monism), which states that consciousness is the only ontologically real substance, whereas all the other material substrates are derivable from it (Berkeley, 1710), that is, brain and other physical phenomena are real, but only as the subjective contents of consciousness. Further developments of classical solutions to the mind–body problem showed that there is no ‘pure dualism’ or ‘pure idealism’, but instead there are many different versions of them (Morton, 2010), some of which continue being considered and developed.
Not all ontologies of mind that take the physical world seriously seek to reduce or eliminate phenomenal consciousness. For instance, embodiment theories (one of 4E theories; for a general introduction, see Newen et al., 2018) assume that experiences depend on the physical world, but processes and entities which are necessary for consciousness are not limited to the brain: the contents of consciousness are ontologically dependant on causal relations with peripheral body as well as with external physical objects (Noë & Thompson, 2004b; Thompson & Varela, 2001). Another relevant example are enactivist theories, first introduced by Varela et al. (1991), which see cognition as the result of dynamic sensorimotor interactions between an acting organism and its environment – a relationship of codetermination (Di Paolo & Thompson, 2014; Varela et al., 1991; Ward et al., 2017). Unlike cognitivist approaches, in which organisms are seen as passive receivers of information from their environment, the enactivist view conceives organisms as actors that shape their experience by how they act (Hutchins, 1996). As argued by Thompson (2010), the enactivist view addresses the explanatory gap in the hard problem of consciousness by building a bridge between experience and the brain. While the formulation of the hard problem often takes a dualistic form, the enactivist view embraces experience by asserting that science is also enacted and as such it arises from humankind’s interactivity with the world, through the interaction between sensorimotor patterns of perception and action. Crucially, enactivist theories conceive cognition not as the mere processing of information in a computationalist sense, but rather, as an exercise of skilful know-how in situated and embodied action. Therefore, while traditional cognitive views conceive cognition as information processing based on a prespecified external realm represented internally, the enactivist view conceives cognition as a relational domain enacted.
4E theories such as embodiment and enaction theories seem plausible in the case of visual perception, as one of the prerequisites of vision is external physical stimulation of the retina, yet embodiment and enactive theories do not provide a satisfactory explanation of how internal subjective experiences are generated in the absence of behavioural embodied interactions with the world, such as during mental imagery or dreaming.
3.2 Biological approaches to consciousness
The majority of the empirical researchers of consciousness tend to follow assumptions of biological naturalism, which is a scientific common-sense position that consciousness is a qualitative, subjective, unified and (usually) representational high-level brain function (Searle, 1992, 2007, 2015). Even though consciousness is thought to be caused by lower-level brain processes, it also seems to be irreducible to them and having its own causal efficacy to influence cognition and behaviour. Consistently with these intuitions, biological realism assumes that phenomenal consciousness is a real biological phenomenon that resides within the confines of the brain and cannot be reduced to the fundamental laws of physics (Revonsuo, 2009). Given that being real implies having its own causal powers (Kim, 1992), consciousness is assumed to be subordinated to causal interactions with other neural processes in the brain and, through them, with the rest of the body and the physical world. Methodologically, biological realism proposes that subjective experiences as biologically real phenomena should be studied by biology. In particular, the material basis of consciousness should be studied within the spatial and temporal scales that are common for neurosciences. Further, biological realism assumes that we are not cognitively closed to the understanding of consciousness, but instead biological sciences, especially cognitive neuroscience, are (or will be) able to explain it (Revonsuo, 2009). Thus, metaphysically, biological realism represents the weak emergent materialism, which predicts that the complete understanding of the brain will explain how consciousness emerges from brain processes (Revonsuo, 2010). Contrary to this, strong emergent materialism assumes that the neural basis of consciousness will remain unresolved even when all facts about the human brain will become known to the scientific community.
While explanations in physics typically involve descriptions of universal natural laws through which an explanandum, that is, the target of explanation, can be reduced to physical processes at the smaller spatial and temporal scales, explanations in biological sciences involve descriptions of multilevel mechanisms whose causal interactions are too complex to be described by universal laws (Bechtel & Richardson, 1993; Craver, 2007; Craver & Darden, 2001). Instead, biologists, including cognitive neuroscientists, develop constitutive, contextual and aetiological explanations of their explanatory target (Revonsuo, 2009). Constitutive explanations reveal the lower levels of a phenomenon under investigation, for example, neuronal long-term potentiation can be described as an emergent outcome of the lower-level NMDA receptor activation (Craver & Darden, 2001). Contextual explanations point to the functional roles of explanandum in interaction with the higher levels of biological organisation. Aetiological explanations describe biological processes that are capable of modulating the explanandum, but cannot constitute it, such as abnormal developmental pathways. Consciousness is assumed to reside in the largely unknown phenomenal level of organisation in the brain (Revonsuo, 2009), yet it is expected that descriptions of interactions between the cognitive, phenomenal and neural level of organisation will eventually solve the mind–body problem (Bechtel & Mundale, 1999).
However, despite continuous development of a biological research programme during the previous decades and its relative success in identifying some neural correlates of consciousness, the hard problem of phenomenal consciousness remains unsolved. Thus, even though cognitive neuroscience and neuropsychology have already yielded a number of fascinating discoveries and neurocognitive theories, it remains possible that some other approaches to consciousness than the one defended by biological realism might prove to be more accurate. For instance, it might turn out that phenomenal consciousness emerges as a product of multilevel functional interactions between biological entities and processes, and that these interactions can be replicated in artificially designed systems. This would contradict the assumptions of biological naturalism and realism, which are regarded as an alternative to functionalism (Revonsuo, 2009; Searle, 1984). Likewise, information integration and differentiation theory of consciousness (Tononi, 2012; Tononi et al., 2016) may loosen its ties with neuroscience if consciousness-generating informational complexity would be detected or designed in non-biological systems.
Until science and philosophy provide conclusive evidence and arguments, it may be too early and premature to commit to a single ontology of consciousness, as this may hinder other more accurate, yet currently neglected possibilities. 2 Importantly, none of the discussed philosophical theories of the mind–body relationship neglects the importance of biological research, as even in substance dualism non-material entities are thought to be able to cause changes in the brain (Descartes, 1641), which subsequently can be studied empirically. Thus, successful implementation of the methodology of biological realism may lead to, but currently does not require, the acceptance of emergent materialism and is compatible with metaphysical indeterminacy. While it is feasible to focus on the neurobiology of consciousness, we do not know yet where we will end up in trying to explain the hard problem of consciousness.
4. The contribution of Francisco Varela to the study of consciousness
Varela was deeply concerned about the nature of consciousness; particularly, with developing methods that could address the hard problem of consciousness by building a bridge between objective and subjective sources of data, or in his own words: first-, second, and third-person perspectives. In addition, Varela investigated the neural patterns underlying experience and perceptual awareness; his work on these matters led to the idea that consciousness does not happen in specific brain regions, but rather, it is a product of different neural systems communicating with each other.
Varela made three major contributions to the field of cognitive science: the enactivist theory, which proposes that cognition arises from the dynamic interaction between an organism and its environment; the neurophenomenological framework, a methodological programme that proposes an approach to study experience in a pragmatic way; and the study of neural phase synchronisation, an index of neural integration of cognitive acts involved in the emergence of consciousness. A lesser-known contribution of Varela was the conceptualisation and study of time consciousness by putting together contributions of phenomenology of time, neurophenomenology, and neural connectivity tools.
Given the scope of this article, in the following sections we focus on neurophenomenology, neural phase synchronisation, and time consciousness as they were the contributions of Varela that impacted consciousness research in a more explicit, direct, and specific manner.
4.1 Neurophenomenology
How can the hard problem of subjectivity be addressed? Varela proposed a pragmatic approach called neurophenomenology (Varela, 1996) that seeks to narrow the gap between the qualitative and quantitative aspects of consciousness contents. To achieve this, Varela proposed that rigorous first-person methods must be developed to examine qualitative experience rigorously, thus giving phenomenal data the same level of importance enjoyed by objective data in cognitive science.
4.1.1 Bridging the gap between qualitative and quantitative information
The neurophenomenology programme entails training researchers in the skill of phenomenological description since subjective experience is not completely private as it can be shared (for a recent practical account, see Berkovich-Ohana et al., 2020). In deep, Varela sought to revolutionise how cognitive science addressed consciousness by cultivating a systematic capacity for reflexiveness and phenomenological examination (Varela & Shear, 1999). Varela was inspired by the works of western phenomenologists such as Edmund Husserl (1859–1938) and Maurice Merleau-Ponty (1908–1961); he argued that Husserl’s phenomenological reduction technique could also be used to develop an empirical science of consciousness (Varela, 1997).
Varela (1996) defined four intertwined moments in the neurophenomenological framework as part of phenomenological reduction: 1. Attitude: reduction. A self-induced suspension of reference to contents such as beliefs, expectations, and fast elaborations, followed by a redirection of thought towards the process through which contents are constituted. This way, neurophenomenology stresses the importance of tolerating the suspension of conclusions that allow new aspects or insights of a phenomenon to unfold, ideally making this self-observing act as discrete and automatic as possible, in other words, to help the participant redirect their attention towards their implicit know-how processes. 2. Intimacy: intuition. To gain intimacy with the phenomenon of study so the way it is experienced can appear less encumbered and more vividly present. This gain in intimacy with the phenomenon becomes the basis of the criteria of truth in phenomenological analysis. According to Varela, is this intimacy with our experience what it is traditionally referred to as intuition. 3. Descriptions: invariants. The descriptive process must determine phenomenological invariants, that is, controlled variations of the subjective experience or ‘eidetic variations’ as defined by Husserl (1901). These invariants should be intersubjectively shareable or communicable. As explained by Varela, this is comparable to what mathematicians have been doing for centuries, but neurophenomenological invariants refer to contents (or modes) of consciousness. 4. Training: stability. Stabilising and deepening one’s capacity for attentive bracketing and intuition to also illuminate descriptions. This can be achieved by investigating repeated experiences by the researchers themselves.
One crucial aspect here is that a process of description must take place in order to define phenomenological invariants. Such an extraction of invariants supposes a work on controlled ‘variations’ of the experience – what Husserl called ‘eidetic variations’ – in order to specify ‘the appropriate dimensions of mental states’ (Husserl, 1901; Varela, 1996). Therefore, the phenomenological description mentioned above should aim at a dynamic picture that can represent the naturalistic embodied nature of experience.
Another important aspect of neurophenomenology is its constrained nature. In order to reduce the distance between subjective and objective (or qualitative and quantitative) information, Varela proposed aiming at finding a mutual determination between lived experience and biological mechanisms. By searching for a circulation between both while constraining each source of information with the other, neurobiological data could help illuminating the domain of subjective experience and vice versa (Varela, 1976, 1996). It was this dynamical nature of the phenomenology that made Varela look into different neural dynamical processes in his search for candidates for neural mechanisms of consciousness (see below).
Importantly, neurophenomenology requires subjects to act both as a subject of a particular task but also as a subject who can report their own experience. Therefore, neurophenomenology requires awareness to provide accurate phenomenological descriptions.
4.1.2 Mutual circulation: bridging the gap through second-person interviews
Mutual circulation is a neurophenomenological implementation proposed by Varela and Shear (1999) that intends to bridge first- and third-person sources of information through second-person interviewing methods. First-person research assumes that the subject and object of study are equivalent (as we study our own acts of knowing; second-person research defines an interviewer that collects data from an interviewee (e.g. their ‘lived experience’), and third-person research collects objective data from another subject (i.e. another person’s acts of knowing). Thus, mutual circulation allows for communication between phenomenological and neurobiological data. However, studying one’s own experience to provide accurate and meaningful reports requires training – it requires subjects to act both as subject of a particular task and as subject who can accurately report their own experience. The mediation between second- and third-person approaches may enhance this mutual communication, for which different interview methods have been developed (Olivares et al., 2015).
Intersubjectivity is a key concept for the implementation of interviewing methods. Since interviews may be prone to bias – for example, an interviewer may inadvertently influence the interviewee’s answers – Varela and Shear (1999) proposed an objectivity notion. By identifying invariant structures across participants (i.e. interviewer and interviewee), interviewing methods may achieve intersubjective validation, thus allowing for replicability. Intersubjectivity, therefore, does not call for eliminativist or reductionist views of consciousness (Olivares et al., 2015; Thompson et al., 2005), but rather, for a more fluid and accurate communication between participants.
An important task in neurophenomenology is to distinguish between the ‘content’ of a moment in consciousness or of a mental act and the ‘process’ through which such contents appear (Varela, 1999b). In Varela’s view, a moment of consciousness involves the emergence of a content in the flow of consciousness that has uncompressible duration. Because the constitution of a conscious moment involves high temporal integration of contents in a transitory manner, Varela thought that consciousness might be the result of transient organising synergy of mass action between distant brain areas; such a process would bring unity to a conscious moment while providing it with lack of stability, thus explaining or resembling its transitional nature (Rudrauf et al., 2003; Varela, 1995).
Multiple studies have taken a neurophenomenological approach to study consciousness. For example, Lutz et al. (2002) trained participants to verbally describe and categorise their experience right before being presented with three-dimensional illusions on a screen. Their brain activity was recorded using EEG. Then, by conversing with the experimenter, phenomenological clusters were defined as an attempt to determine invariant aspects of their experience. Later on, EEG data were classified according to these clusters for dynamical analyses. They found patterns of endogenous synchronisation in frontal electrodes before visual stimulation, which were related to the first-person descriptions given by the participants, thus demonstrating the existence of a relationship between behavioural, neurophysiological, and first-person data. In a more recent study, Martial et al. (2019) used a neurophenomenological approach to study near-death experiences (NDEs). They induced NDEs by using hypnotic suggestions during hypnosis. Hypnotic suggestions are verbal suggestions that can induce very compelling changes in people’s perception and cognition (Canales-Johnson et al., 2012; Lanfranco et al., 2014; 2021; Oakley & Halligan, 2013). The researchers recruited participants who had NDE in the past and used hypnotic suggestions to help them re-experience their NDEs while measuring their brain activity using EEG. By measuring EEG and collecting phenomenological reports, the researchers were able to isolate aspects of the EEG signal that may be related to phenomenological invariants in the hypnotically induced NDEs.
4.2 Neural synchronisation and perceptual awareness
Consciousness can be seen as an integrative feature of cognition that arguably always has a unified cognitive content. Because consciousness is believed to involve integration among different perceptual and cognitive functions, it was proposed that a neural correlate of consciousness should entail large-scale integration across different neural systems (Engel & Singer, 2001). Based on past studies on nonlinear dynamic systems, chaos, and time–frequency analyses, Varela began conceptualising the brain as a dynamical system (Letelier, 2001); he proposed that neural synchronisation between different neuronal assemblies could enable the dynamic integration necessary for such a unified perceptual experience in a given cognitive instance, thus binding together sensory, emotional, mnemonic, and motor information (Thompson & Varela, 2001; Varela, 1995).
Varela stressed the idea of consciousness as dense moments of synthesis in the flow of experiences that could not be temporally compressible (Engel & Singer, 2001; Thompson & Varela, 2001). This phenomenal integration contained in conscious experience could involve high – yet transient – temporal integration of multiple contents between distant brain regions. The brain, as a dynamical biophysical system constrained by multiple non-stable attractors, should contain such integrative mechanisms. Based on these ideas and others, Varela proposed that transient phase-locking between brain ensembles could be the mechanism of large-scale integration in the brain which, in turn, could be a condition of possibility for the emergence of consciousness (Thompson & Varela, 2001; Varela, 1995). Synchronisation between neural assemblies would be only limited by the time needed to establish stable states between them, thus perhaps accounting for the transitoriness and temporal flow of experience (Uhlhaas et al., 2009; Varela, 1999b). In addition, mere variations in the spatial scale of the synchronised neural assemblies could provide a switch between conscious and unconscious information processing (Dehaene et al., 2006; Thompson & Varela, 2001). This modular perspective agrees with two processing modes found in small world networks (of which the brain seems to be a case): local modularity and long-range connectivity (Buzsáki, 2007; Yu et al., 2008). Thus, dynamic shifts in coupling between the same anatomical structures in a network could support both local (unconscious) and global (conscious) processes.
The most plausible mechanism for large-scale integration, and therefore for the emergence of consciousness, is phase synchronisation across multiple frequency bands, according to Varela (Thompson & Varela, 2001; Varela et al., 2001). Neuronal assemblies can exhibit a wide range of oscillations in the theta to gamma frequencies (6–80 Hz) as measured with EEG, with precise phase-locking or synchrony (Lachaux et al., 1999). Varela’s team developed a measure of synchrony called phase-locking value (PLV) to detect synchrony in a given frequency band between two recording sites or electrodes, irrespective of signal amplitude. PLV can take any value between zero and 1, where 1 denotes no phase difference between recording sites (See Lachaux et al. (1999) for a more detailed description). There are several newer phase-based connectivity measures today, such as imaginary coherence (Nolte et al., 2004), phase-slope index (Nolte et al., 2008), phase-lag index (Stam et al., 2007), and weighted phase-lag index (Vinck et al., 2011), all of which provide more reliable phase synchronisation indices. For a detailed technical explanation of the different inter-trial phase synchronisation indices, see Cohen (2014, chapter 26).
Multiple studies have supported the role of neural phase synchronisation in conscious processing. More specifically, that long-range synchronisation in the beta and gamma frequency bands may play a role in the emergence of the unified perception of a given stimulus. For example, Rodriguez et al. (1999) presented participants with Mooney faces for 200 ms on a screen. Mooney faces are binary images that can be easily recognised as faces when presented upright but are seen as meaningless shapes when presented upside down. After each stimulus presentation, participants had to press a key indicating whether they saw a face. When participants reported seeing a face, a transient episode of large-scale phase-locking between electrodes was found around 250 ms after stimulus presentation (Figure 1(a)). This episode of phase synchronisation was found mainly in the gamma frequency band (30–80 Hz). However, no significant synchronous ensemble was found when participants reported not having seen a face. These findings were interpreted as evidence in favour of a relationship between perceptual awareness and large-scale neural synchronisation. Consciousness studies measuring phase synchronisation. (a) Main results of Rodriguez et al. (1999). By presenting Mooney faces in upright and inverted orientations, they manipulated perceptual awareness. When participants reported seeing a face (perception condition), a transient episode of large-scale phase-locking between electrodes was found around 250 ms after stimulus presentation. A period of strong desynchronisation marked the transition between face perception and motor response, which was interpreted as a process of active decoupling of neural ensembles marking the transition between cognitive states. (b) Main results of Melloni et al. (2007). By presenting masked words and controlling the luminance of the masks, they manipulated the visibility of the stimuli, thereby influencing participants’ perceptual awareness. Perceived words were associated with a transient long-distance synchronisation of gamma oscillations across widely separated brain regions, suggesting that perceptual awareness is associated with an early transient global increase of phase synchronisation of oscillatory activity in the gamma frequency range. A: Copyright 1999 by Springer Nature. Reprinted and adapted with permission. B: Copyright 2007, Society for Neuroscience.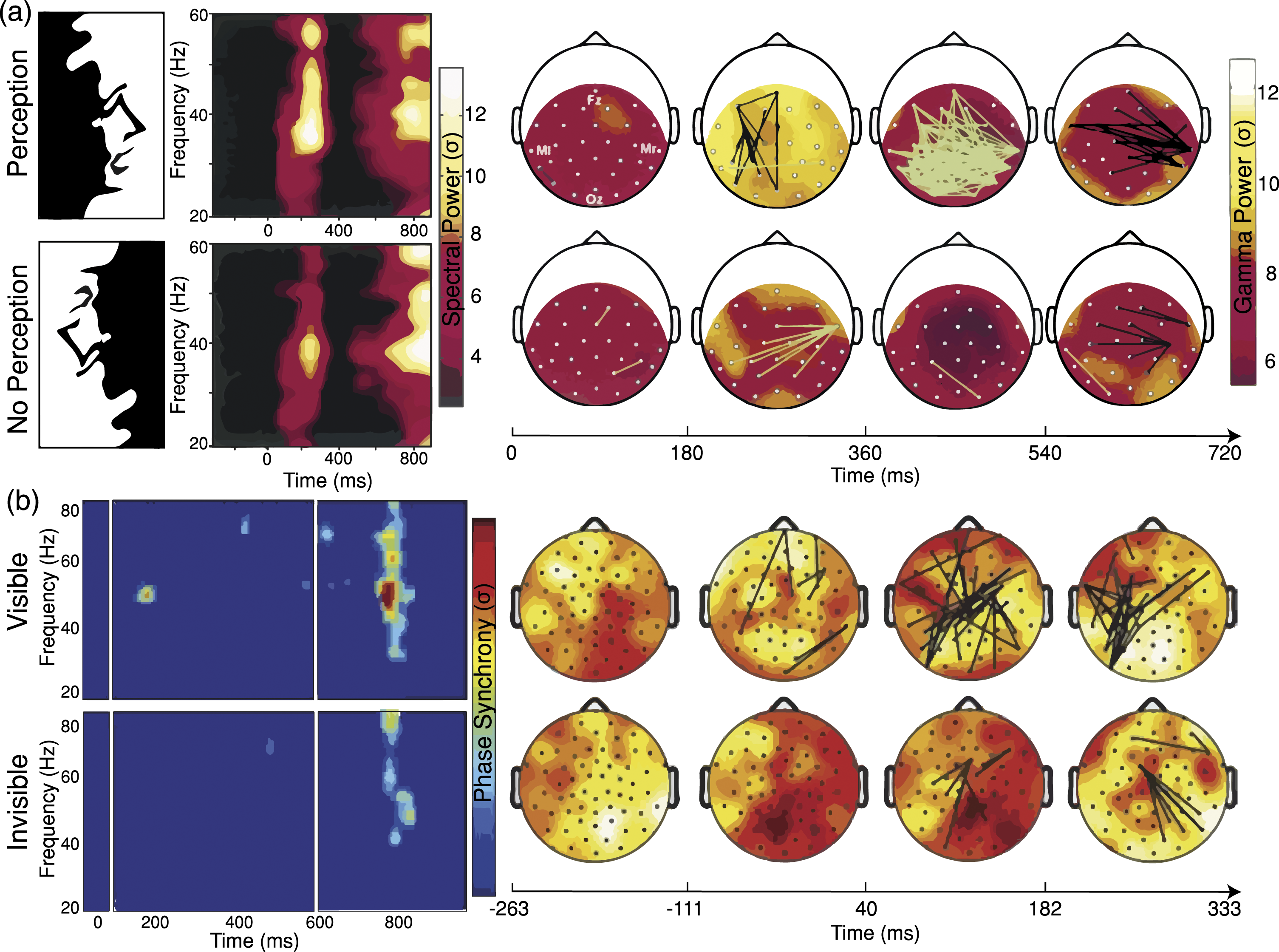
In another study, Melloni et al. (2007) directly compared the EEG signatures of conscious and unconscious processing by asking participants to detect and identify a briefly presented word (33 ms) between masked stimuli. Crucially, they adjusted the strength of the masks such that in half of the trials participants could not report seeing the word. After a 500 ms delay, a second word was presented. Participants had to judge whether the two words were the same or different. Researchers found that an early and transient burst of long-distance synchronisation in the gamma frequency in the visible condition but not in the invisible one (Figure 1(b)). While the amplitude and patterns of gamma oscillations were spatially homogenous and similar for both conditions, the patterns of phase synchronisation significantly differed from each other. Interestingly, this transient period of neural synchronisation was followed by an increase in amplitude (P3 component), a positive deflection in voltage that has been interpreted as a correlate of information transfer into working memory (Jensen & Tesche, 2002; Schack et al., 2005). These findings indicate that conscious processing of visual stimuli may be associated with an increase in phase synchronisation in the gamma frequency band that is independent of spectral power. Studies using binocular rivalry found similar phase synchronisation effects associated with perceptual dominance (Cosmelli et al., 2004; Fries et al., 1997), that is, when comparing having a unified and clear perceptual experience rather than a mixed one when presented with two different images to each eye.
For many years, it was believed that phase synchronisation in the gamma frequency band was necessary for consciousness, especially when it occurred between long distances across the brain cortex. However, these studies did not dissociate visual awareness from selective attention in a rigorous manner, leaving room for the possibility that said findings may not be specific for consciousness. When visual awareness and selective attention are dissociated, high-range phase synchronisation in the gamma band correlates with attention irrespective of stimulus visibility as reported by participants, whereas only mid-range phase synchronisation in the gamma band accounts for stimulus visibility (Wyart & Tallon-Baudry, 2008). Crucially, however, phase synchronisation in gamma has been also found during anaesthesia and NREM sleep (Imas et al., 2005; Murphy et al., 2011), epileptic seizures (Pockett & Holmes, 2009), and unconscious emotion processing (Luo et al., 2009), thus indicating that phase synchronisation can occur in absence of conscious experiences (Canales et al., 2007).
Recent studies have shown that a neural metric specifically indexing distributed information sharing can capture changes in conscious state and conscious content (Canales-Johnson et al., 2020b; Imperatori et al., 2019; King et al., 2013; Sitt et al., 2014). For instance, information sharing has been shown to capture network reconfiguration both in healthy (Canales-Johnson et al., 2020a; Imperatori et al., 2019) and pathologic (King et al., 2013; Sitt et al., 2014) states of alertness. Importantly, due to its sensitivity to highly nonlinear coupling (Imperatori et al., 2019), this metric has been useful for distinguishing conscious contents during bistable perception in the same frequency ranges in which phase synchrony does not (Canales-Johnson et al., 2020b).
Today, gamma-band phase synchronisation has been disregarded as the specific neurophysiological mechanism of consciousness (Koch et al., 2016). However, the evidence still supports the claim that phase synchronisation may play an important role in neural communication between distant brain regions (Arnulfo et al., 2020; Misselhorn et al., 2019), which in turn may be necessary for selective attention (Doesburg et al., 2008; Rohenkohl et al., 2018) and visual integration (Uhlhaas et al., 2009). Recently, for instance, we found that the act of imagining faces is accompanied by long-range gamma-band phase synchronisation (Figure 2), arguably due to visual binding of facial features (Canales-Johnson et al., 2021b). Participants had to study different celebrities’ faces prior to the task. In the imagery condition, they had to imagine those faces inside of a grey oval presented on a screen. They were asked to press a key when the visual image of the face was the most vivid. In the control condition, they were instructed to press as soon as they saw the empty grey oval. Importantly, this occipitoparietal gamma-band phase synchronisation pattern predicted subjective ratings of the contour definition of the imagined faces, thus supporting our interpretation that gamma-band phase synchronisation may be involved in the visual binding of imagined faces. Gamma-band phase synchronisation (wPLI) during visual mental imagery, reported by Canales-Johnson et al. (2021b). (a) Difference in frontoparietal phase synchronisation between frontal and parietal electrodes comparing imagery and control conditions based on cluster-based permutation tests. (b) Difference in occipitoparietal phase synchronisation between occipital and parietal electrodes comparing imagery and control conditions based on cluster-based permutation tests. On the right, topographical representations of such differences between wPLI electrode pairs are shown for each condition. Grey circles represent single-participant wPLI for the clusters depicted on time–frequency charts. Red horizontal lines represent the group mean, whereas the rectangle represents the standard error of the mean. Content licensed under a CC BY 4 international licence.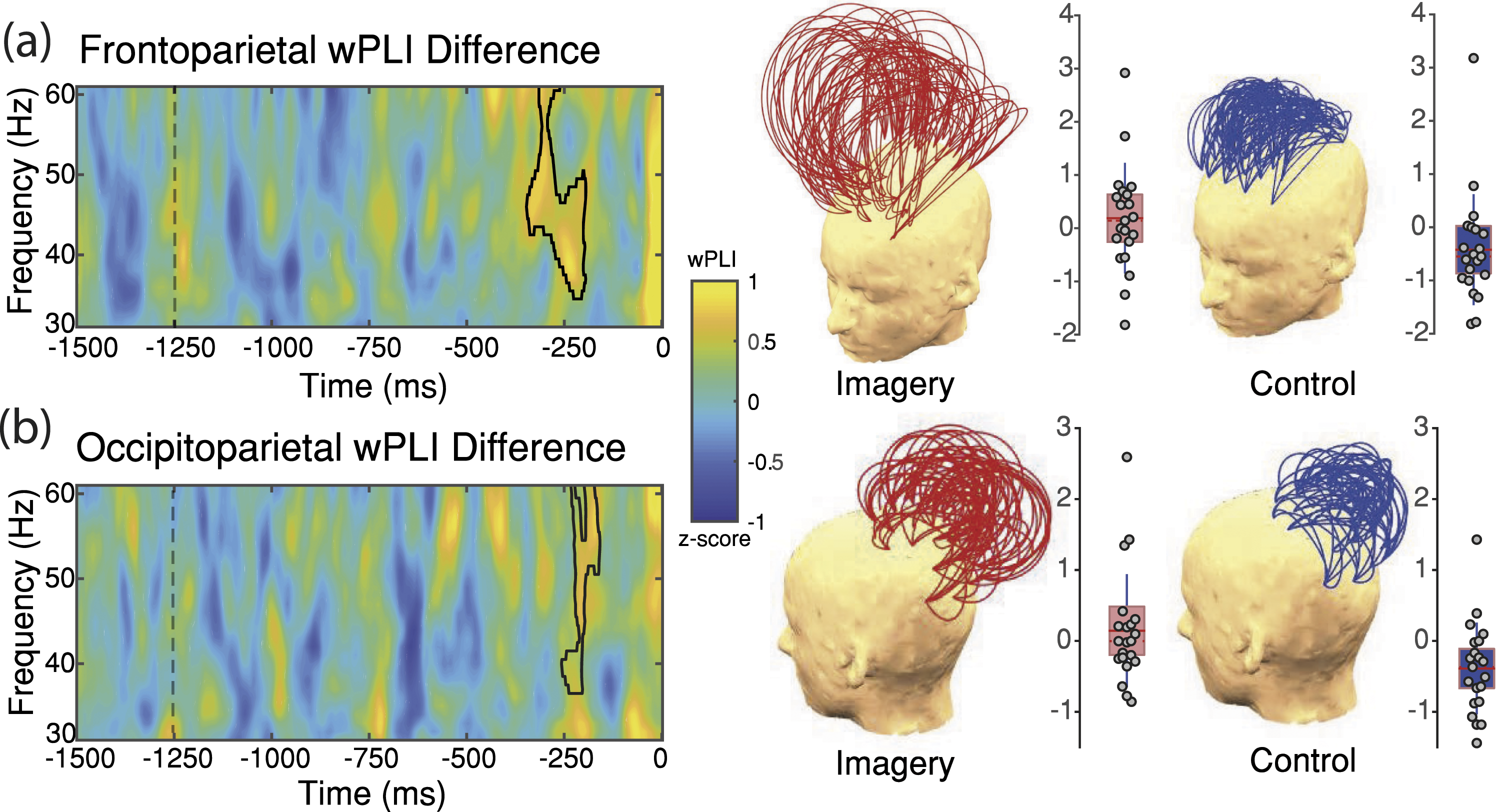
However, EEG gamma-band activity can be affected by the contraction of extraocular muscles, which led researchers to question whether gamma-band activity may have been confounded by miniature saccades (Yuval-Greenberg et al., 2008). While the spikes induced by miniature saccades are not rhythmic, they can still affect power analysis – specifically, inducing a broad-band power elevation between 30 and 100 Hz. However, it has been argued that this concern should not encompass gamma-band phase synchronisation as this measure has been demonstrated using microelectrode recordings of single neurons (Maldonado et al., 2000), pairs of neuron groups (Gray et al., 1989), and local field potentials (Fries et al., 2001) – none of these recordings are affected by the electric fields induced by miniature saccades (Fries et al., 2008).
In summary, Varela laid the groundwork to index and perhaps quantify conscious processing. Despite the fact that phase synchronisation has been rejected as a neural marker specific to perceptual awareness, Varela’s work was of prime importance in the field and inspired countless later developments on the neural signatures of consciousness contents.
Varela was not only interested in time as a biological feature or mechanism of consciousness, as implied in his neural phase synchronisation studies; he was also interested in time as an unavoidable feature of phenomenology. Next, we describe Varela’s approach to study the experience of time.
4.3 The neurophenomenology of time consciousness
How does time manifest itself in our experience? Time is often thought in third person as the simultaneity of moments of a physical event with times infinitesimally continuous in relation to a clock, making it seem objective and independent of consciousness. This is a notion of time inherited from classical physics. While this framework may be homologous to that developed by modern theory of computation, it is not compatible with a neurophenomenological view that focuses on the experience of time (Varela, 1999a).
4.3.1 The triple structure of time
Varela’s understanding of time was deeply influenced by the works of Husserl, who conceptualised time as a triple structure in three inseparable moments: past, present, and future emerge (Husserl and Brough, 1990). If time is solely studied from a third-person perspective, past and future do not exist – they cannot be addressed and only the present time would be susceptible of objective measurement when compared to a clock. Conversely, past and future are conceived as special modes of presentation when adopting a first-person perspective – the past is primary a retentional mode of consciousness of the lived present, attached to the perception of the moment, whereas the future is a protentional mode of consciousness whereby we expect something to happen but without having a content of it (Husserl and Brough, 1990). Rather, the future in this perspective is like a horizon that opened to what could happen despite not knowing what will occur. In summary, retention and protention can be seen as two modes of consciousness in the present time (or two different intentional directions within time consciousness); therefore, past–present–future are three inseparable moments or instances of the structure of time (Vargas et al., 2013). Contrary to the physical view of time, Husserl conceived time as the consciousness of internal time or living present; that is, a description of the experience of objective time.
The past–present–future structure can be exemplified by a study published by Lutz et al. (2002), which involved recording participants’ EEG activity while they were presented with three-dimensional illusions; they were also required to report if they saw the three-dimensional figure and their experience of it. Based on these reports, the researchers identified two states of preparation: prepared (when participants felt ready to see the image) and unprepared (when they did not feel ready and only saw it because their eyes were correctly positioned). Crucially, they found that different neural patterns of large-scale gamma-band phase synchronisation predicted these phenomenological categories: Prior to visual presentation, they found a neural pattern of large-scale phase synchronisation around frontal electrodes in the prepared participants but not in the unprepared ones. In addition, they found that the degree of dispersion in phase synchronisation around posterior electrodes increased alongside the degree of unpreparedness. Finally, they also found a pattern of large-scale synchronisation in prepared participants had an earlier onset (300 ms before motor response) than unprepared participants (600 ms). Here, retention can be understood as neural activity that arose before and remains part of the present neuronal ensemble; that is, neural activity in the present that is associated with the cognitive act immediately preceding (Lutz, 2002; Varela, 1999b). Lutz et al. (2002) interpreted these results as evidence of the recent past being present at the time of consciousness, which fits Varela’s view of a protentional character of the lived present, that is, a form of opening to the next moment of consciousness or ‘disposition for action’ (Varela, 1999b). Furthermore, Varela suggests that affective tone plays a modulating role in this process, thus enabling stays and breaks in the flow of consciousness; this would be exemplified in the feeling of surprise experienced by participants during their unprepared states and in the maintenance of the system in a region of phase space whence the onset of the visual stimulus emerges, thus reorganising the system into a new neural pattern (e.g. involving phase scattering pattern) and taking more time to yield a motor response (Lutz, 2002).
4.3.2 The neural correlates of time consciousness
Similarly, and inspired by Husserl’s work, Varela distinguishes three levels of time consciousness or temporality with the purpose of searching for each of these levels’ neural correlates: First, the minimal duration for consciousness of objects and events, that is, the physical time (also used in experimental psychology), which can be measured by the clock. For example, in the fall of an object, we have the same body passing from one place to another, which is set to correspond in the awareness of an observer with a clock giving us the time. Varela described the neurobiological basis of this level as the neural activity that occurs within the scale of 1/10 (Varela, 1999a; Vargas et al., 2013). For instance, when two lights are flickered one after the other with an interval lower than 0.1 seconds, they are typically perceived as simultaneous (Varela, 1999b). As this interval increases, perception changes and both stimuli are perceived as one moving quickly between two places. EEG studies have found that this transition in perceiving the stimuli as a sequential movement correlates with neural activity in the alpha band (7–13 Hz) and that it significantly depends on whether the presentation onset of the stimuli falls in the positive or negative cycle phase (Gho & Varela, 1988; Varela et al., 1981). Similarly, subsequent studies have shown that even the probability of detecting individual flashes alone can be affected by the phase of spontaneous EEG oscillations at the moment of the flash, particularly in the low alpha and theta bands (Busch et al., 2009; Mathewson et al., 2009) when the stimuli are attended (Busch & VanRullen, 2010).
The second level is the time of the acts of consciousness – the immanent or internal time. For instance, in the fall of an object, how we experience it. Importantly, this does not correspond to the minimal duration for awareness of the object’s change in position but rather to the presence of time in the act of becoming aware of such a change. This level of phenomenological description is what Varela attempted to ground using the scale of 1 in neural activity: integration-relaxation (Varela, 1999b). For instance, Cosmelli et al. (2004) studied the neural correlates of perceptual alternations using magnetoencephalography (MEG). The researchers employed binocular rivalry, which consists in presenting two unmergeable images, one to each eye. Instead of perceiving both images overlapped or mixed, people experience one or the other as wholes; that is, one image exhibits complete perceptual dominance over the other. Participants had to indicate the moment they experienced complete dominance of a stimulus. The researchers found that a widely distributed neural network was activated when participants were observing both stimuli; however, the conscious perception of one of them (perceptual dominance) correlated with the time course of a more specific and synchronised cortical network in the frequency band of the presented stimulus (5 Hz) – a pattern of long-scale phase synchronisation. This study exemplifies the logic behind Varela’s view of time consciousness: temporality would be in the scale 1 of the duration of acts of consciousness, a scale that can be paired with patterns of long-scale synchronous activity observed during perceptual dominance during binocular rivalry tasks. Since the two visual images are invariably presented in this experiment, changes in perception over the leaps between dominant images must be determined by acts of consciousness and not by variations in the physical images shown. Therefore, the duration of phase synchrony patterns can be matched with the duration of perceptual dominance of each one of the two images. In other words, a perceived image, as an act of consciousness, lasts a certain period of time without changing the perceived object (Varela, 1999b).
The third level is the absolute flow of consciousness. Varela defined this level as the flow of consciousness itself. As opposed to the duration of an object (first level) and the time of an act of consciousness (second level), the third level is more subtle and consists of a remaining fluent that is an absolute (or independent) form from what we perceive or experience, that is, our linguistic ability to behave as a self and develop a personal identity (Varela, 1999b). According to Varela, this third level may be specially disrupted or impaired in disorders such as schizophrenia, which typically involve reports of insertion of thoughts and control delusions. Patients oftentimes attribute their thoughts and actions to external agents; however, the sense of ownership of their consciousness flow remains intact (Gallagher & Varela, 2003). While Varela does not propose a candidate for the neural mechanisms of the scale 10, he does suggest it would have to be capable of linking together cognitive moments of the scale 1 in order to form a broader temporal horizon (Varela, 1999b).
In summary, Varela’s understanding of time consciousness mainly (but not exclusively) stems from Husserl’s phenomenology; Varela employs his neurophenomenological framework to study time consciousness by actively linking first-person phenomenological reports with third-person scientific methods (Vargas et al., 2013). Each experiment presented here involves an external observer that links participants’ reports (phenomenology) to the interpretation of their neural data (neurobiology).
5. Conclusion
Consciousness and its contents can be studied scientifically through different methods. However, its conceptual complexity, experiential nature, and philosophical problems make its study of great difficulty. Therefore, different approaches have been taken to empirically address consciousness and multiple theories have been proposed.
Varela contributed to the descriptive and scientific study of consciousness. His enactivist proposal is still relevant today in the study of cognition and consciousness, especially from a philosophical perspective (Di Paolo et al., 2017). The enactivist perspective proposes that consciousness is not limited to brain activity; but rather, it is situated more broadly in a self-organised dynamic system coupled with its environment.
One of Varela’s main contributions to consciousness research was neurophenomenology, a pragmatic approach that attempts to build a bridge between the realms of objective and subjective data. Neurophenomenology requires researchers to deal with qualitative data (e.g. phenomenological descriptions) and quantitative data (e.g. EEG signal) while giving each of them equal importance, care, and rigour. By mutually constraining both sources of data, neurophenomenology allows researchers to find mutual determinations between experience and neural mechanisms, which in turn helps exploring the transitions between contents of consciousness. Therefore, neurophenomenology addresses consciousness using a non-reductive approach that treats experience and neurobiology as equally important sources of data.
Varela also proposed a candidate for the neural correlates of consciousness: large-scale gamma-phase synchronisation. He argued that phase synchronisation could provide a means for different neuronal assemblies to communicate and thereby enable dynamic integration for perceptual awareness. A number of studies suggested that large-scale synchronisation in the gamma frequency range might participate in the emergence of conscious experience. Even though subsequent studies have cast doubt on such claims, phase synchronisation remains a relevant neural correlate of neural communication and possibly of information integration.
One aspect of subjective experience that intrigued Varela was time consciousness. Varela was deeply influenced by Husserl’s phenomenology of time. Varela proposed a neurophenomenological approach to explain how time emerges in the flow of consciousness by using neurobiological methods and asking participants for detailed descriptions of their experience. He found neural signatures that were able to distinguish between these experiential differences and speculated about the role of neural oscillations in the emergence of past, present, and future.
Twenty years after Varela’s death, his work continues to inspire new lines of research. For example, very recently researchers have proposed a neurophenomenology-inspired approach to studying consciousness by linking phenomenological data with computational modelling methods (Ramstead et al., 2021). This approach called computational phenomenology uses generative models to formally describe the structure and dynamic unfolding of lived experience. While this and similar proposals (Vilas et al., 2021) that tackle consciousness from an active inference perspective (Marvan & Havlík, 2021) may not seem compatible with 4E theories at first glance, some argue that active inference accounts are susceptible to non-representationalist enactive interpretations (Ramstead et al., 2020) – generative models would not encode information but rather express or realise embodied activity.
Overall, Varela’s work on consciousness was innovative and multidisciplinary, spanning fields that typically progress separately, such as philosophy of mind, phenomenology, cognitive science, neuroscience and experimental psychology. His contributions still remain relevant today.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by a postdoctoral fellowship awarded by Karolinska Institutet and by a PhD studentship awarded at the University of Edinburgh by ANID/CONICYT to Renzo Lanfranco.
Notes
About the Authors