
Abstract
Pheromones are chemical substances released into the environment by an individual animal, which elicit stereotyped behaviours widely found across the animal kingdom. Inspired by the effective use of pheromones in social insects, pheromonal communication has been adopted to swarm robotics domain using diverse approaches such as alcohol, RFID tags and light. COSΦ is one of the light-based artificial pheromone systems which can emulate realistic pheromones and environment properties through the system. This article provides a significant improvement to the state-of-the-art by proposing a novel artificial pheromone system that simulates pheromones with environmental effects by adopting a model of spatio-temporal development of pheromone derived from a flow of fluid in nature. Using the proposed system, we investigated the collective behaviour of a robot swarm in a bio-inspired aggregation scenario, where robots aggregated on a circular pheromone cue with different environmental factors, that is, diffusion and pheromone shift. The results demonstrated the feasibility of the proposed pheromone system for use in swarm robotic applications.
1. Introduction
Pheromone is one of the most effective communication mediums widely used by animals (Camazine et al., 2003; Wyatt, 2014). It is a complex chemical substance that allows intraspecific communication by being secreted into an environment and detected by agents eliciting behavioural and developmental changes. From yeast and small insects such as ants and bees to mammals such as dogs and humans, a wide range of insects and animals adopt pheromone as a communication medium (Fields, 1990; Hölldobler & Wilson, 1990; Preti et al., 2003). Those species benefit from pheromone that has distinct characteristics compared to visual, audio and tactile communication. Especially, social insects use pheromone communication widely (Hölldobler & Wilson, 1990).
In swarm robotics which is the field of study that how large numbers of simple robots coordinate for a shared goal inspired by nature (Şahin, 2005), pheromonal communication has been adopted by researchers. Pheromonal communication has a number of useful characteristics for swarm robotics as well as in nature. For example, pheromone allows a group of agents, which consist of extremely simple sensory-motor and memory capabilities, to achieve fully decentralised control (Khaliq & Saffiotti, 2015). Furthermore, pheromone optimises behaviour of an entire group of robots. Researchers have implemented pheromonal communication with various methods such as chemical substances, radio-frequency identification (RFID) and virtual environments.
COSΦ is a low-cost light-based artificial pheromone system which ensures high degree of flexibility and fidelity (Arvin et al., 2015). This work and its extension (Na et al., 2019) investigated collective behaviours of a robot swarm with environmental factors including evaporation, diffusion and wind effect on artificial pheromone as they are ever-present phenomena in nature. The work is one of the early studies that embodied environmental effects and investigated impact of environmental factors on collective behaviours of robotic swarm. However, bidirectional communication between the pheromone system and the robots was not achieved; in other words, the robots reacted the given pheromone without releasing any additional pheromone into the system.
In this article, we proposed a novel artificial pheromone system for swarm robotic applications. This article improves the state-of-the-art as (1) highly precise pheromone release mechanism is implemented due to the high resolution of LCD screen. (2) High flexibility of the system is achieved since the characteristics of the artificial pheromones can be easily modified in the software. (3) Cost-effectiveness of the system enhances the ease of utilisation in swarm robotics studies. (4) Environmental effects, for example, wind effect and diffusion, are implemented in the system based on the reliable mathematical models to make it realistic and having high fidelity. We presented a detailed study of impact of environment factors involving bidirectional pheromonal communication on collective behaviour of robots, which we called it pheromone injection. We specifically investigated influence of diffusion in diverse settings including static and dynamic cue settings and pheromone injection on aggregation behaviour of robots that operate based on Φ-Clust algorithm (Arvin et al., 2018), which was a pheromone-based bio-mimetic simulated swarm.
2. Related works
2.1. Pheromone in biology
Pheromone is defined as chemical substance secreted to the outside of an individual and detected by conspecifics triggering them to conduct stereotyped behaviour and/or have developmental changes (Karlson & Lüscher, 1959). Pheromones are essential factors in animal communication and trigger physiological change in a wide range of behavioural and ecological contexts in nature (Baracchi et al., 2017; Chalissery et al., 2019; Hostachy et al., 2019; Okosun et al., 2019; Tateishi et al., 2020). Pheromones play important roles as communication means for diverse taxonomic groups from yeast and insects to mammals (Fields, 1990).
Pheromonal communications were found in vertebrate animals (Brennan & Zufall, 2006). For example, 2-methylbut-2-enal, which is contained in rabbit milk, improves nipple-search behaviour of rabbit pups (Schaal et al., 2003). Several studies have found that the physiological and psychological effects elicited by chemosignals arise also in humans (Mutic et al., 2017; Preti et al., 2003; Stern & McClintock, 1998; Wedekind & Furi, 1997). For example, it is found that female reproductory state is affected by male axillary extract which mediates the hormonal change in female brain (Preti et al., 2003). These results showed that pheromonal communication is widely used across diverse taxonomic groups including human beings.
Although a broad spectrum of animals use pheromonal communication as their communication mechanism, social insects utilise pheromone most effectively (Hölldobler & Wilson, 1990). Pheromone enables a whole group of social insects to communicate effectively as an externalised and spatialised shared memory although the individuals have limited memory and sensory-motor capability. Also, agents only need local sensing ability to detect pheromone rather than global sensing. Moreover, a collective behaviour in a group of social insects can be optimised appropriately deploying pheromones (Denny et al., 2001; Goss et al., 1989). For example, the Argentine ant Iridomyrmex humilis finds the shortest path from its colony to the food source using pheromone and its feedback mechanisms (Goss et al., 1989; Hölldobler & Wilson, 1990).
Monomorium pharaonis, called Pharaoh’s ants, which are usually found in human habitats, utilise multiple pheromones that are vital for food-foraging behaviour in dynamic and competitive environments (Jackson & Ratnieks, 2006). They create pheromone trails to fetch to and return from the food source using three types of trail pheromones: non-volatile attractive pheromone, volatile attractive pheromone and repellent pheromone. The three different kinds of pheromones work, respectively, as long-term memory of the trail, attraction leading to the currently rewarding trail and a stop-sign. The sophisticated use of the three types of pheromones enables the ant colony to create optimal pheromone trails from the nest to the food source in dynamically changing environments. Bombus hortorum, known as bumblebee, leaves chemical cue on flowers which allows detection and avoidance of recently depleted flowers (Eltz, 2006). Similarly, the use of chemical cue enhances the efficiency of foraging behaviours of bee colonies since it prevents meaningless visits of depleted flowers. Majority of social insects utilise queen pheromones. This type of pheromones characterise queen and other reproductive individuals and are extremely important to maintain the whole colony (Holman, 2018; Princen et al., 2019). For example, a queen pheromone of Lasius niger, known as the black garden ant, regulates worker sterility so that the reproduction behaviour of the colony is controlled by the queen (Holman et al., 2010). As another example, if the queen of a colony fails by any reasons including viruses and pesticides, the secretion of queen pheromone in the colony decreases, and as a result the colony might collapse (Steinhauer et al., 2018).
2.2. Pheromone in robotics
Swarm robotics is the study of how large number of simple agents for a collective behaviour achieved by local interactions among the agents and/or between the agents and the environment (Şahin, 2005). The effectiveness of pheromonal communication is significantly aligned with the needs of swarm robotics. First of all, pheromonal communication requires a simple capability for individual robot since it only needs local sensing (Khaliq & Saffiotti, 2015). Second, regardless of the size of the environment, the individuals only need to have a limited memory since the environment contains information about the state and actions via released pheromone (Jackson et al., 2006). Third, its potential to optimise the performance of a group task via use of combination of multiple types of pheromone and feedback mechanisms is highly desirable for robot swarms (Denny et al., 2001). Finally, no external control is needed with pheromone-based self-organisation; therefore, a fully decentralised swarm system can be achieved (Jackson et al., 2011). Several researchers have noticed the potential of pheromonal communication in swarm robotics; thus, they adopted pheromonal communication in the simulation and physical robot systems. One of the early works inspired by pheromone communication was introduced by Russell (1999). This work employed Cinnamomum camphora, known as Camphor, as implementation of trail pheromone for trail following behaviours for the robotic systems which embodied odour releasing and sensing functionality. The results of this work demonstrated feasibility of using pheromone in robotic systems as a communication medium. Similar to the work introduced above, Fujisawa et al. (2014) used ethanol as a pheromone for the robotic system. The work proved that a complete autonomous robotic system that conducts cooperative behaviours with trail following can be achieved using pheromone communication. Despite the successful implementation of chemical-based pheromone communication for robotic systems introduced above, practical difficulties in sensors and actuators are incurred such as cost and resolution of chemical detecting sensors (Purnamadjaja & Russell, 2010).
A number of studies proposed robotic systems using RFID tags as a medium for pheromonal communication (Herianto & Kurabayashi, 2009; Khaliq & Saffiotti, 2015). In these studies, RFID tags were distributed on the floor in the environment where the robots operate. The tags store data transmitted by robots passing above and elicited the corresponding behaviour depending on the data transmitted previously, which works like pheromones in nature. One of the recent works using RFID tags (Alfeo et al., 2019) show its usability in real-world application. This work utilised RFID tag-based pheromone communication for autonomous waste management in urban environment. The work demonstrated pheromone-based communication robotic system outperformed the conventional communication method. Although the approach that uses RFID tags is reliable and usable, it requires appropriately equipped environments with RFID tags, which might challenge users when preparing the environments are not feasible.
Recent studies involved virtual environments to implement artificial pheromone into swarm robotic systems. Campo et al. (2010) proposed a mechanism for path selection by a foraging robot swarm using virtual ants. In this mechanism, robots locally transmitted and received messages. The messages were referred to as virtual ants and they deposited the virtual pheromone on the robots, thereby indicating path. While this work implemented virtual pheromone only within robots, the other works created virtual map to mark deposited pheromone accessible to all the robots. As an example, Reina et al. (2017) introduced the augmented reality for Kilobots (ARK) system that creates a virtual environment where Kilobots swarm (Rubenstein et al., 2012), which has been designed for large-scale robotics experiments, transmits and senses information in real-time. Through the system, the robots equipped the virtual sensors and actors and they accessed to the virtual environments shared by all robots integrating overhead tracking and control. In the virtual environments, the robots could release and detect the virtual pheromones. One of the following works utilised this system to investigate quality-based foraging of robots (Font Llenas et al., 2018). Similar to ARK system, Kilogrid (Valentini et al., 2018) was proposed as a virtualisation environment system that primarily uses bidirectional infra-red (IR) communication between Kilobots and the grids mounted under the arena. It also created virtual pheromones in the virtual environments accessible to the robots and the remote PC in real-time. Although the two virtualisation systems covered large-scale swarm and are flexible to utilise than other methods, the systems have technical limitations. It is stated that both ARK and Kilogrid implicitly involves low communication frequency that leads to considerable delay in the communication.
Besides the works that are grouped with similar works, several researchers have implemented pheromone communication system with diverse means. Garnier et al. (2007) projected light from a video projector mounted on the ceiling to realise artificial pheromone trail. This was the first work utilising light as artificial pheromone. Although this work is one of the early works using light, it did not replicate pheromone in real-life as it implemented evaporation in every 5 s. In another work, Mayet et al. (2010) used phosphorescent paint that was only observable by projecting ultra violet (UV) light. This work demonstrated that use of phosphorescent paint and the robot that embodied appropriate sensors can be alternative of existing methods like chemical substances. However, its use with multiple robots was not investigated as this work only experimented with a single robot rather than swarm robots.
COSΦ is the state-of-the-art artificial pheromone system for swarm robotics that used light-based pheromone trails (Arvin et al., 2015). It used LCD screen as the arena for robots to interact with the pheromones which are the light spots displayed on the screen. With the tracking system via a USB camera mounted above the arena, the system updates the position, orientation and ID of the robots and generates virtual pheromone accordingly. Several advantages of COSΦ as an experimental platform were reported. First of all, this system has a significantly high resolution for implementing pheromone compared to the other works. Unlike the systems using fixed size grids, it offers high-resolution field on which the artificial pheromone can be injected with high precision. Second, diverse environments can be implemented since it is a highly flexible system. From thickness of pheromone trails to evaporation rate of pheromone, all characteristics can be easily modified as needed. Third, it uses a low-cost configuration with a basic digital camera (Krajník et al., 2013, 2014) and a flat LCD screen, which makes it accessible to be utilised by many researcher.
While the only basic leader-follower scenario was demonstrated in the first work using COSΦ system to test its validity, following studies have extended the system and investigated collective behaviour of robots. Sun et al. (2019) utilised multiple colours to implement multiple types of pheromone based on the COSΦ system and investigated food-foraging and aggregation behaviours applying both attractive and repellent pheromones. Moreover, this system extended COSΦ system to realise the diffusion effect. The use of multi-colour artificial pheromone and adding diffusion highly improved the system; however, the movement of pheromone was not implemented. The movement of pheromone is essential for the pheromone system to be reliable since it realises wind and other causes of pheromone shift. Na et al. (2019) extended the system to include diffusion and wind effect, which are the phenomena almost ever-present in nature. The work demonstrated the performance of collective behaviour of robot swarm with different diffusion and wind rates affected considerably. It is found that when the robots aggregate on the pheromone cue, diffusion helped the robots approach the centre of the cue closer than without it. Also, wind affected the robots’ individual behaviour to stay longer on the cue than without wind. To the best of our knowledge, it was the first work that investigated direct impacts of both diffusion and wind effect on collective behaviours of robot swarms. However, this work did not accomplish replicating pheromone communication in real world since pheromone injection was not implemented during the experiments. In other words, the communication between the pheromones and robots was only uni-directional. Since complex swarm behaviours are more likely to emerge from bidirectional interplay between robots and pheromones, the work presented here aims to demonstrate the ability of the proposed artificial pheromone system to emulate realistic swarm-pheromone interactions under natural conditions of variable diffusion and advection.
3. Artificial pheromone system
The whole set of the artificial pheromone system proposed in this work consists of two components: (1) pheromone system that computes pheromone and displays on the horizontally placed flat LCD screen, representing the scenario arena and (2) tracking system that tracks robots and sends their data to the pheromone system. The two components enable the system to implement real-time localisation and pheromone injection on the precise locations.
3.1. Pheromone system
The pheromone system simulates the several types of artificial pheromone and their interaction simultaneously. The resultant pheromone is displayed on an LCD screen, and it triggers predefined responsive behaviours of the employed robots. The system displays the resultant pheromone and the rest part of the arena as a grey-scale image with the size of the screen. The brightness of grey-scale image is represented as

where
While the system is running, the intensity of pheromone released on the screen is constantly updated with discrete time steps. The intensity of updated pheromone is given by
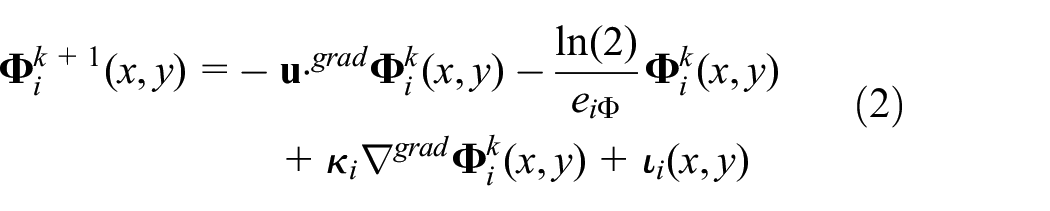
where
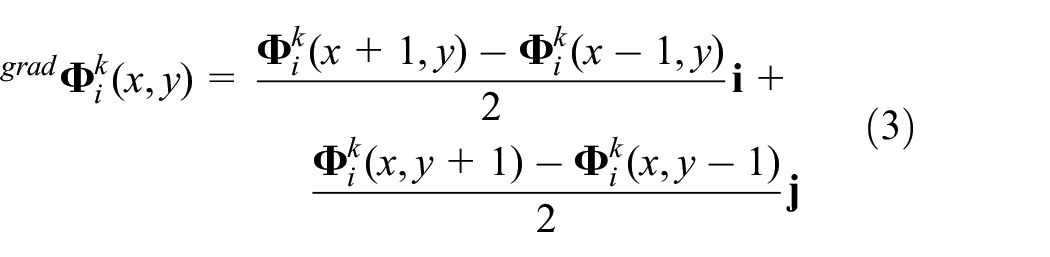
Recomputing equation (2) for every values of
The parameters shown in the right-hand side of equation (2) can be divided into two categories: (1) environmental effects and (2) pheromone injection. Environmental effects include evaporation rate,
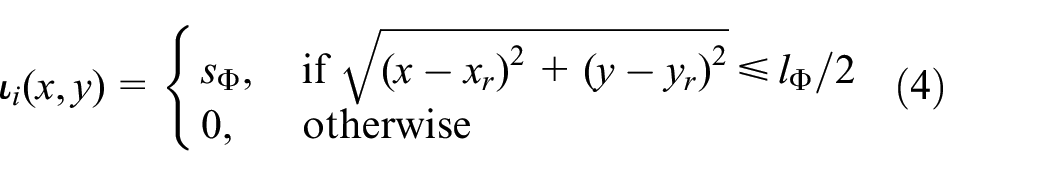
where
3.2. Environmental effects on pheromone
The environmental effects contribute to the versatility of the system in emulating realistic conditions that affect the distribution of the pheromones in the environment over time.
3.2.1. Evaporation
Evaporation is the process by which the surface of a liquid turns into the gas phase. As volatile chemical substances, evaporation occurs in secreted pheromone. In several works on kinetic properties of pheromones, half-life of pheromones, which is the time required for pheromones to decay by half, are investigated as a metric (Vogt et al., 1985). Conforming this practice, the half-life of the pheromone is adopted, that is,
3.2.2. Diffusion
Diffusion is a movement of molecules from a region of higher concentration to a region of lower concentration. Implementing diffusion in the pheromone system is indispensable to be a realistic system since diffused pheromone from the source has a great impact on swarm behaviour (Wyatt, 2003). In this work, diffusion is implemented using Gaussian blur instead of directly using the term,
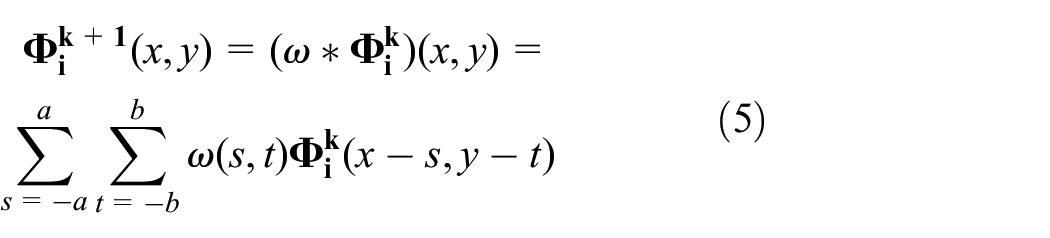
where
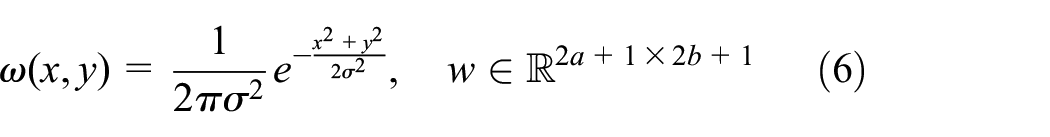
where
3.2.3. Pheromone shift
In real and dynamic environments, the position of the released pheromone can be shifted. A natural cause of the shift in nature is advection, which is the flow of any fluid, for example, air, that transfers pheromone from one position to another. The movement of released pheromone in the same direction is modelled as
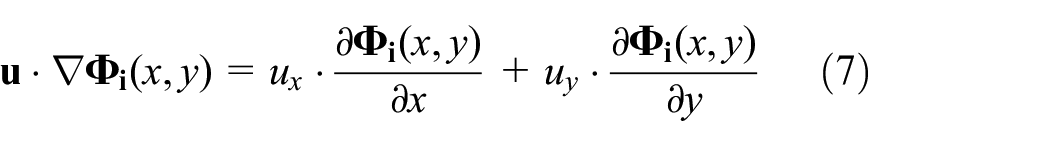
where
3.3. Tracking system
The fast and precise open-source localisation system (Krajník et al., 2014) is used to track the position, orientation and ID of the robots, thereby precisely injecting pheromone at the position of the robots on the LCD screen in real-time. The system captures images using a digital camera mounted above the screen, searches for black-and-white roundel patterns and converts their image coordinates into the real-world coordinates in the Cartesian plane. By attaching unique patterns on the individual robots, each robot can have unique ID. Krajník et al. (2014) claimed that the localisation precision can achieve millimetres scales and hundreds of patterns can be tracked simultaneously in real-time.
By transforming the captured images into the coordinates of the tracked patterns in the defined area and sending the data to the pheromone system, the tracking system allows the pheromone system to release the pheromone at precise positions of the robots in real-time. Precise localisation of the patterns can be achieved through auto-calibration of the system that makes the pheromone system robust under any circumstances, for example, external disturbance during experiments. The system defines the area of the arena in the coordinate system from the image by setting the four corner tags attached on the corners of the frame placed on the screen, and determine the coordinates of patterns inside of the defined area. Since the system requires less than 50 s to calculate one robot position, the difference between the pheromone injection position and the actual robot position is determined by the time required to transfer the image via the USB interface and the delay caused by the graphic interface driver. Nevertheless, the difference between the expected pheromone positions and the actual positions is negligible.
4. Experimental setup
4.1. System configuration
4.1.1. Arena
To implement the artificial pheromone system, a high-definition (HD) 42″ LCD screen with the size of

Experimental setup used for the pheromone system, including a PC for tracking robots and generating pheromone, a digital camera for tracking robots position, a horizontally placed 42″ LCD screen, aluminium frame around the arena and Colias mobile robots.
4.1.2. Robotic platform
Colias micro-robot (Arvin et al., 2014) which was developed for swarm robotic applications was used as a robotic platform. The front and bottom view of the robot is shown in Figure 2. It is a small robot with a diameter of 4 cm having simple functionalities. The robot is a differential wheeled robot which movement is determined by two micro DC gear-head motors directly connected to wheel with the diameter of 2.2 cm. The speed of the robot in the forward motion is maximum 35 cm/s. The rotational speed of each motor is controlled by pulse-width modulation (PWM). Each motor is driven by the embedded H-bridge DC motor driver which draws an average current of 35 ± 5 mA when there is no-load and 150 ± 20 mA at maximum in stall conditions. For its sensing, the robot has three IR proximity sensors which include pairs of IR emitter and receiver in front of the robot. It is used to detect objects, obstacles or other robots, within a distance of approximately 2 ± 0.5 cm. In addition, the robot has two light (illuminance) sensors at the bottom next to the wheels. The light sensors are used to read light intensity on the ground where the robot is located, that is, read the intensity of the pheromone in this work. The robot’s power consumption is approximately 800 mW.

(left) Colias micro-robot, swarm robotic platform and (right) bottom board of Colias with pheromone sensing ability. Different modules of Colias are (a) main processor, (b) IR proximity sensors, (c) digital camera, (d) micro-motors with gearhead,(e) 22-mm wheels, (f) pheromone detectors (light intensity sensors), (g) battery recharging unit, (h) main switch and (i) ISP programming port.
In this work, the behaviour of the robot was determined for achieving pheromone-based aggregation adopted from one of the previous studies (Arvin et al., 2018). The state machine of the scenario is described in Figure 3. The robot begins to move forward after it is switched on. The rotational speed of the left and right wheels,

where

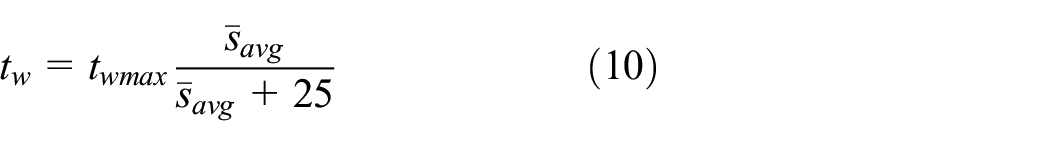
where
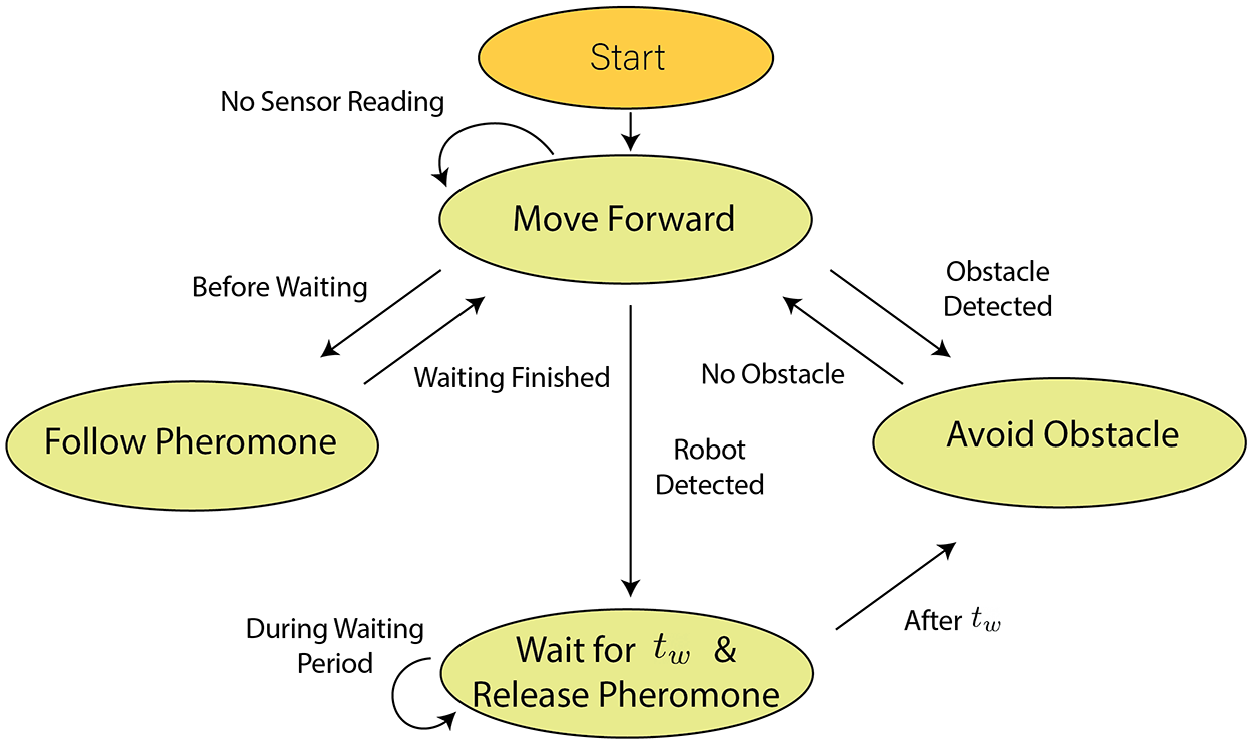
State machine of the implemented swarm scenario.
4.2. Experiments
We designed an experiment plan to systematically investigate the proposed pheromone system. The set of experiments adopted the pheromone-based aggregation scenario. The experiments were implemented with two configurations: (1) without pheromone injection and (2) with pheromone injection by the robots – called as without

The collection of six randomly selected screenshots during experiments. The first row shows the screenshots from an experiment with no diffusion and fast cue speed without pheromone injection and the second row shows the screenshots from an experiment with medium diffusion and fast cue speed with pheromone injection taken at
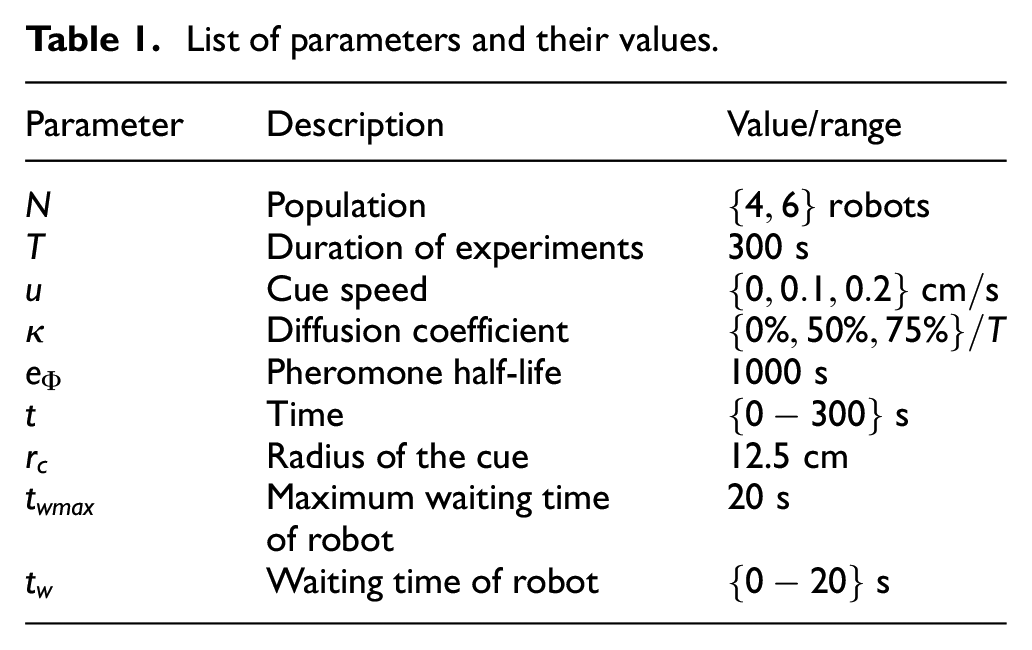
List of parameters and their values.
4.2.1. Static cue configuration
In static cue configuration, the position of the cue (
4.2.2. Dynamic cue configuration
In dynamic cue configuration, the position of the cue (
4.2.3. Metrics
To evaluate the aggregating behaviour of swarm, two parameters were defined: (1) size of aggregate,
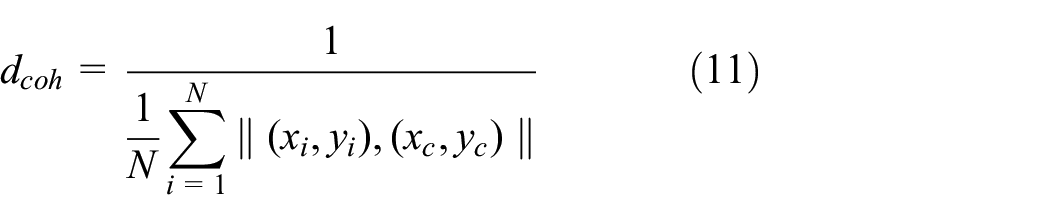
where
4.2.4. Statistical analysis
To statistically analyse the observed results from experiments, analysis of variance (ANOVA) test was conducted. ANOVA test is a test used to analyse the difference among groups caused by difference in factors (Scheaffer et al., 2010). F-statistic (
5. Results
The results of experiments are presented in this section. The results are depicted with line plots. The plots show the aggregation performance, both size of aggregate and cohesiveness with different factors: population, diffusion rate and with different configurations: cue configurations and pheromone injection. In the line plots, a line represents the median of the observed data from five repetition and the shaded region surrounding the line represents the inter-quartile range of the data.
5.1. Static cue configuration
Here, we depict how the aggregation performance of robots varies with different diffusion rates in static cue configuration (
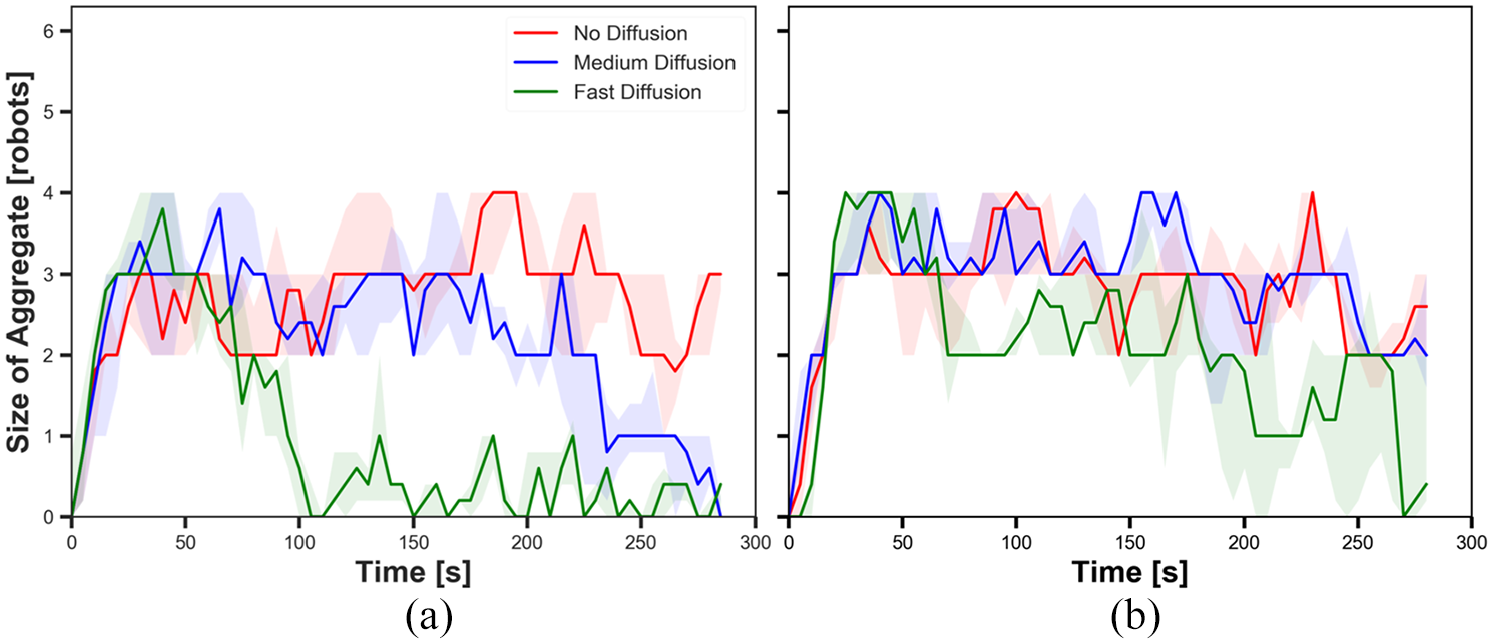
The size of aggregate in experiments with static cue configuration (
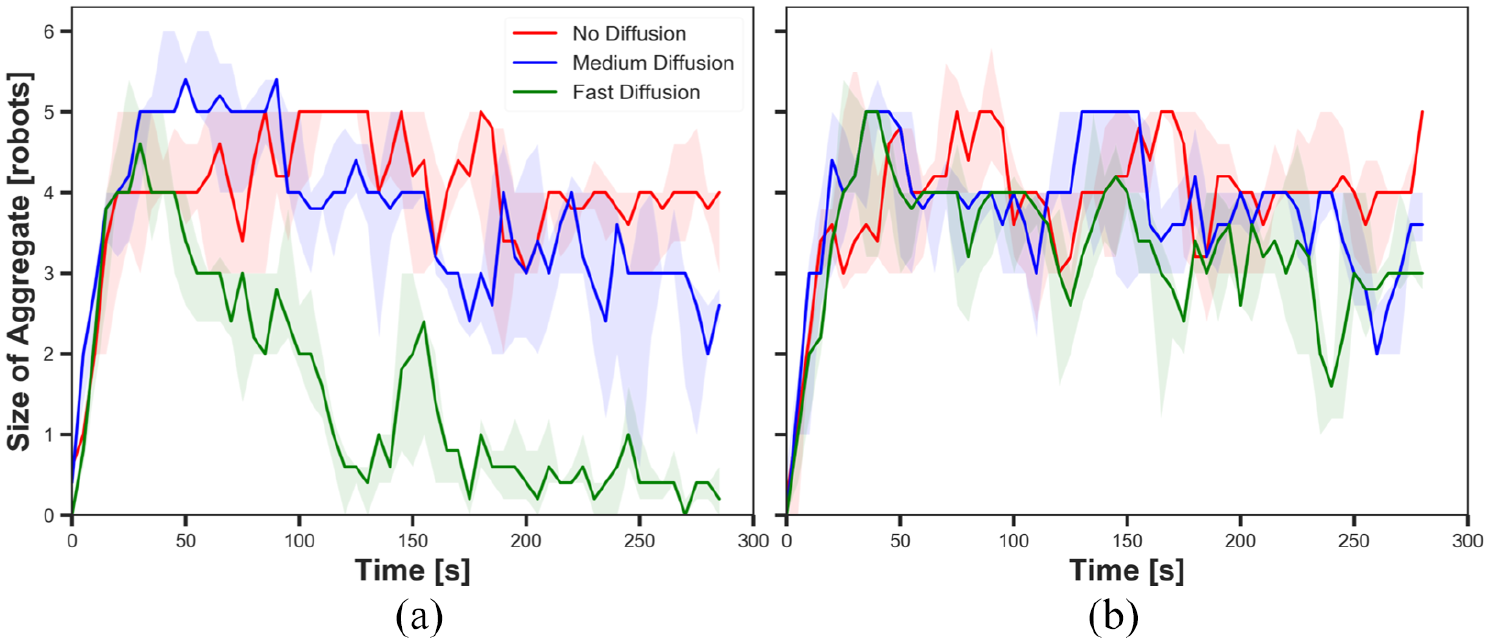
The size of aggregate in experiments with static cue configuration (
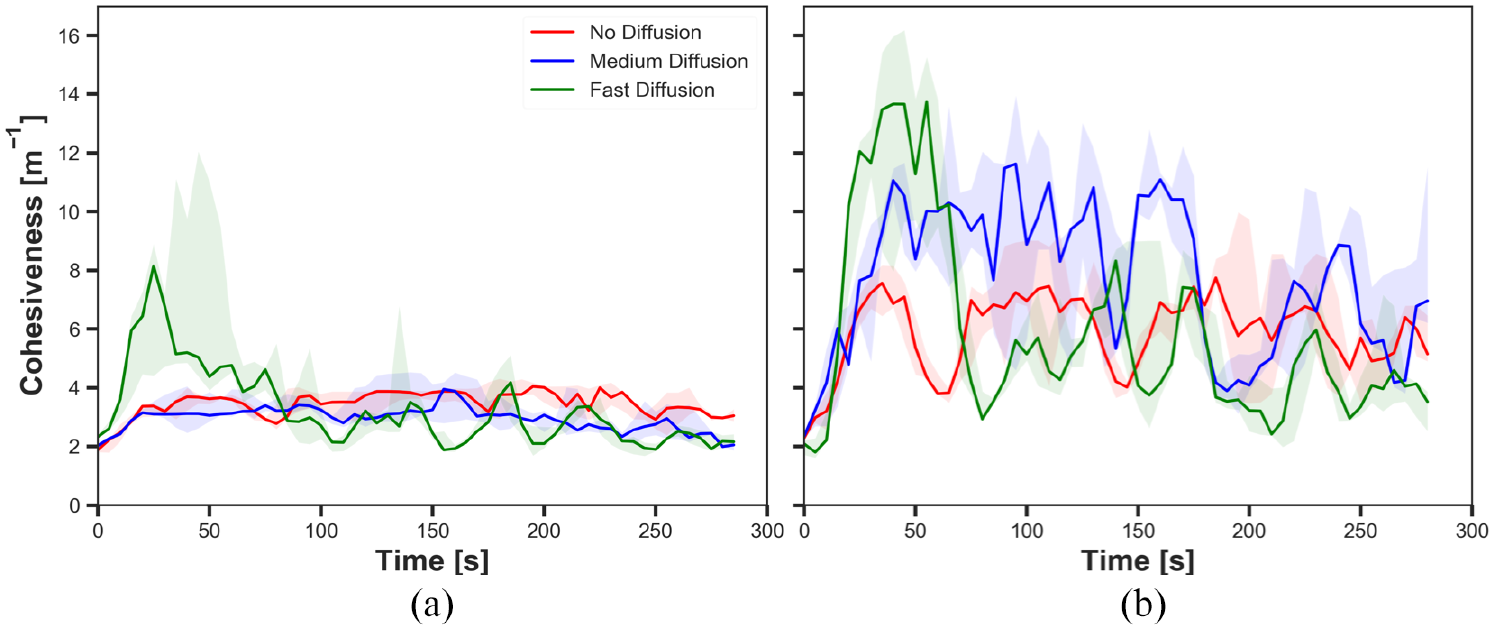
The cohesiveness in experiments with static cue configuration (
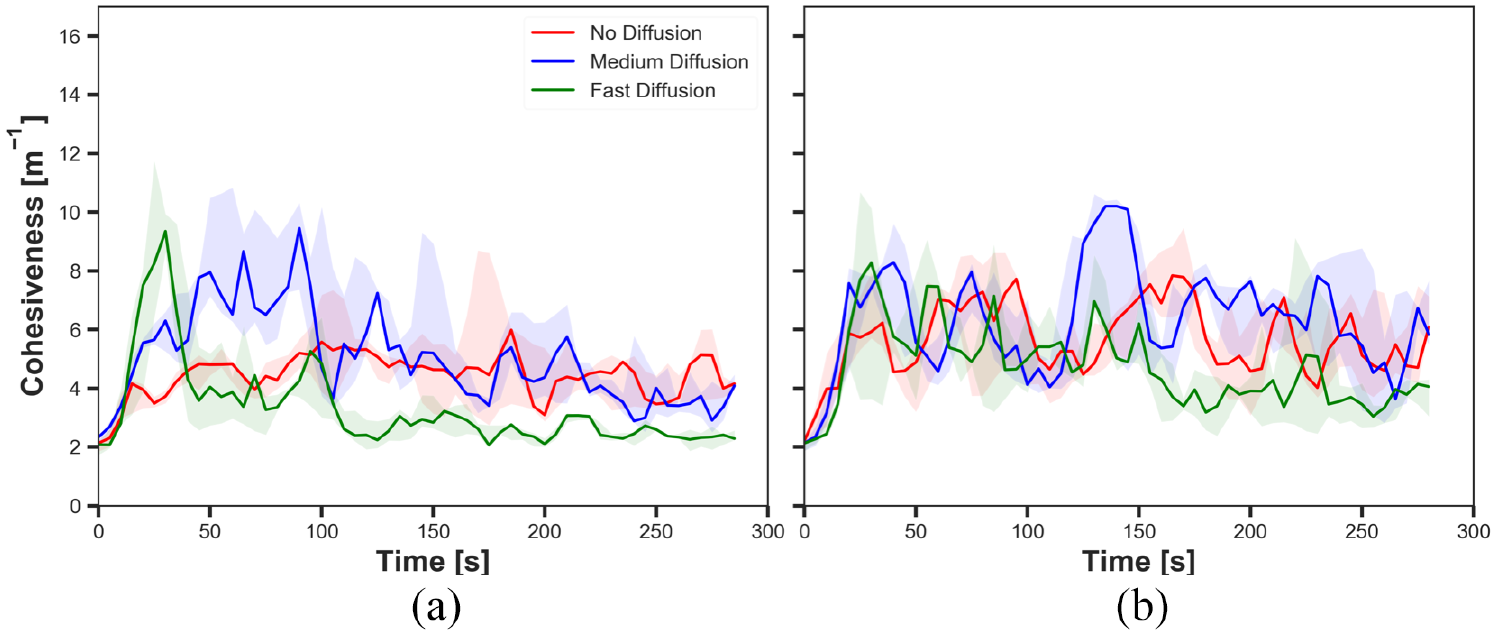
The cohesiveness in experiments with static cue configuration (
5.1.1. Diffusion
First, we investigated the impact of diffusion on swarm behaviour. Figures 5 and 6 show the size of aggregate with three different diffusion rates
Different diffusion rates led different cohesiveness in the robots. In Figure 7(a), cohesiveness with fast diffusion before
As it was expected, diffusion led the gradual decay in pheromone intensity in the system; therefore, robots less decreased their speed when the cue was diffused than when there was no diffusion. The degree of decay in intensity was higher in the outer parts of the cue than in the inner parts. When the robots reached the edge of the diffused cue, they kept moving forward while they stayed on the edge of the cue that was not diffused. As a result, the robots approached closer to the centre of the cue in experiments with diffusion than experiment without diffusion. Therefore, diffusion led high cohesiveness of the swarm while the intensity of pheromone was sufficient for robots to stay on the cue.
5.1.2. Pheromone
Comparing Figures 5(a) and (b), it is shown that robots aggregated in higher probability on the cue throughout the experiments regardless of diffusion rates with
The results suggested that the pheromone injection offsets the impact of diffusion on the size of aggregate. Since the intensity of the cue increases when pheromone is injected, the robots are likely to stay on the cue. Hence, the size of aggregate is higher with
5.1.3. Statistical analysis
To statistically analyse the results, a fully nested ANOVA test with factors of population, diffusion and time was carried out to find significance of the factors. Also, the most effective factor on the size of aggregate and cohesiveness in both configurations: (1) with
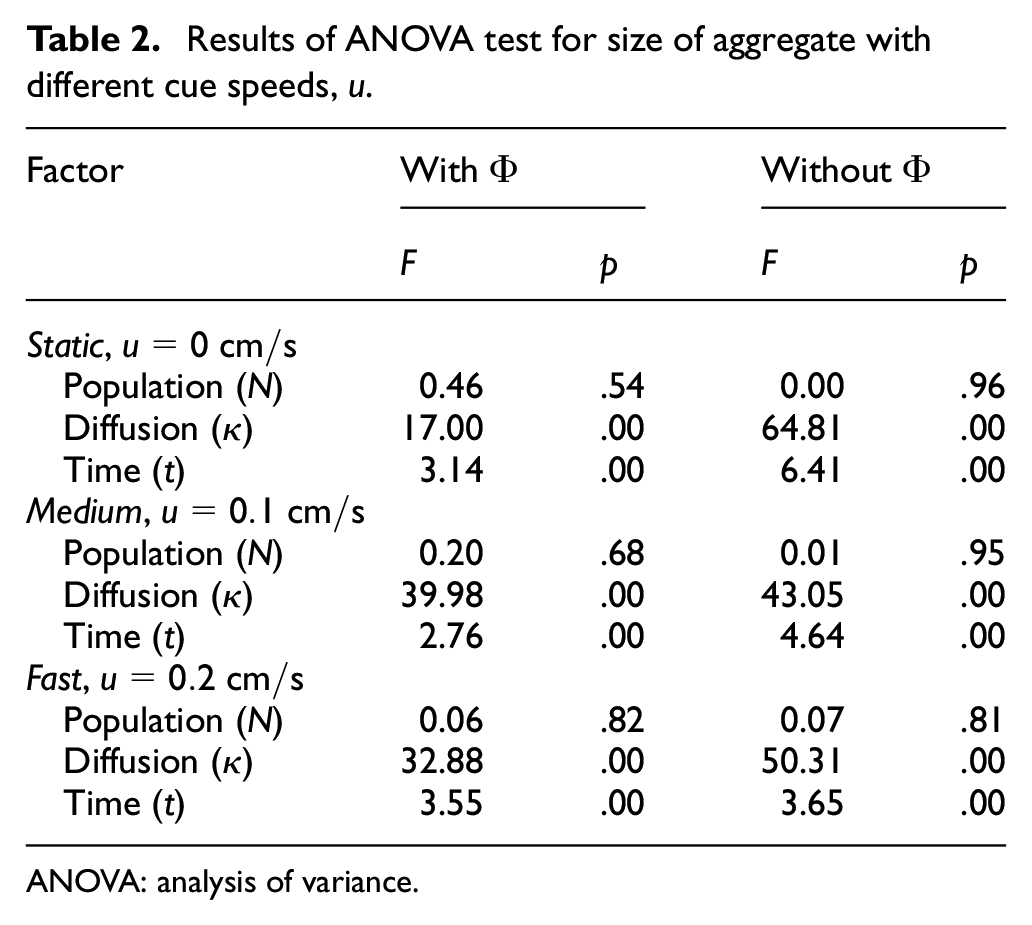
Results of ANOVA test for size of aggregate with different cue speeds,
ANOVA: analysis of variance.
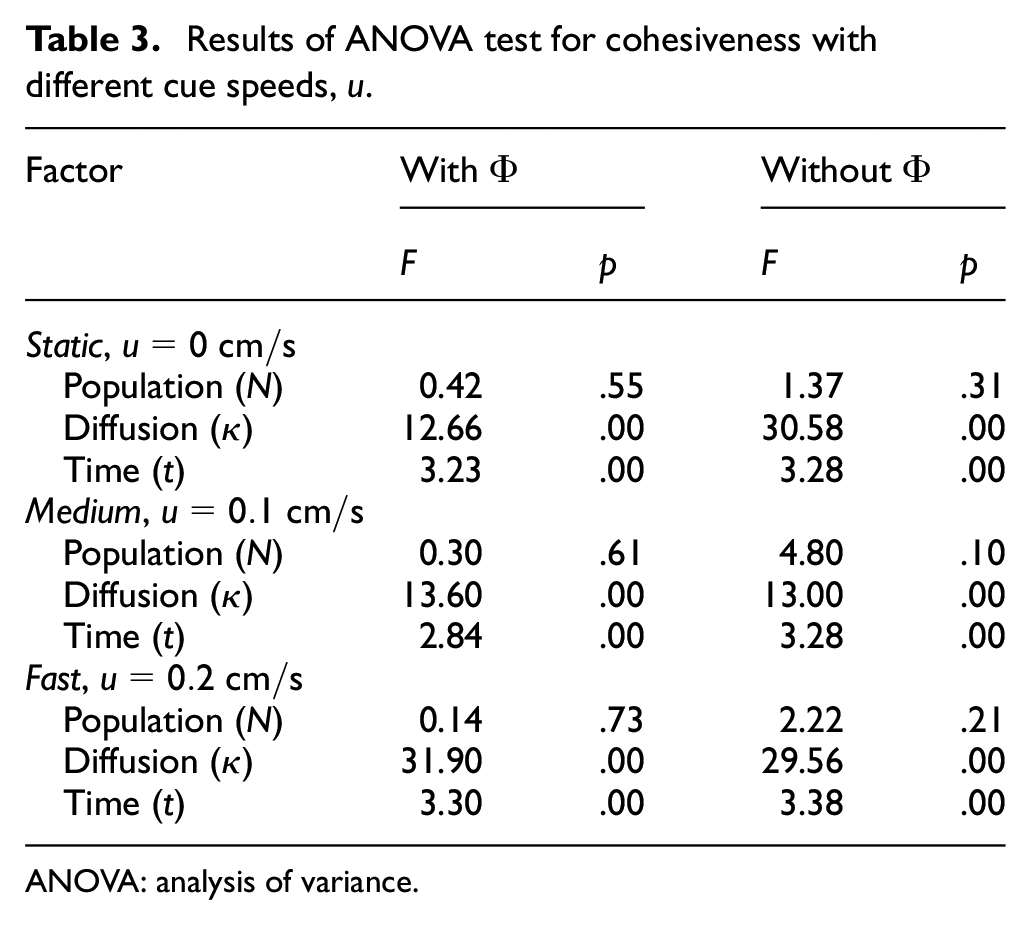
Results of ANOVA test for cohesiveness with different cue speeds,
ANOVA: analysis of variance.
5.2. Dynamic cue configuration
In this section, the aggregation performance of the robots with three different diffusion rates (
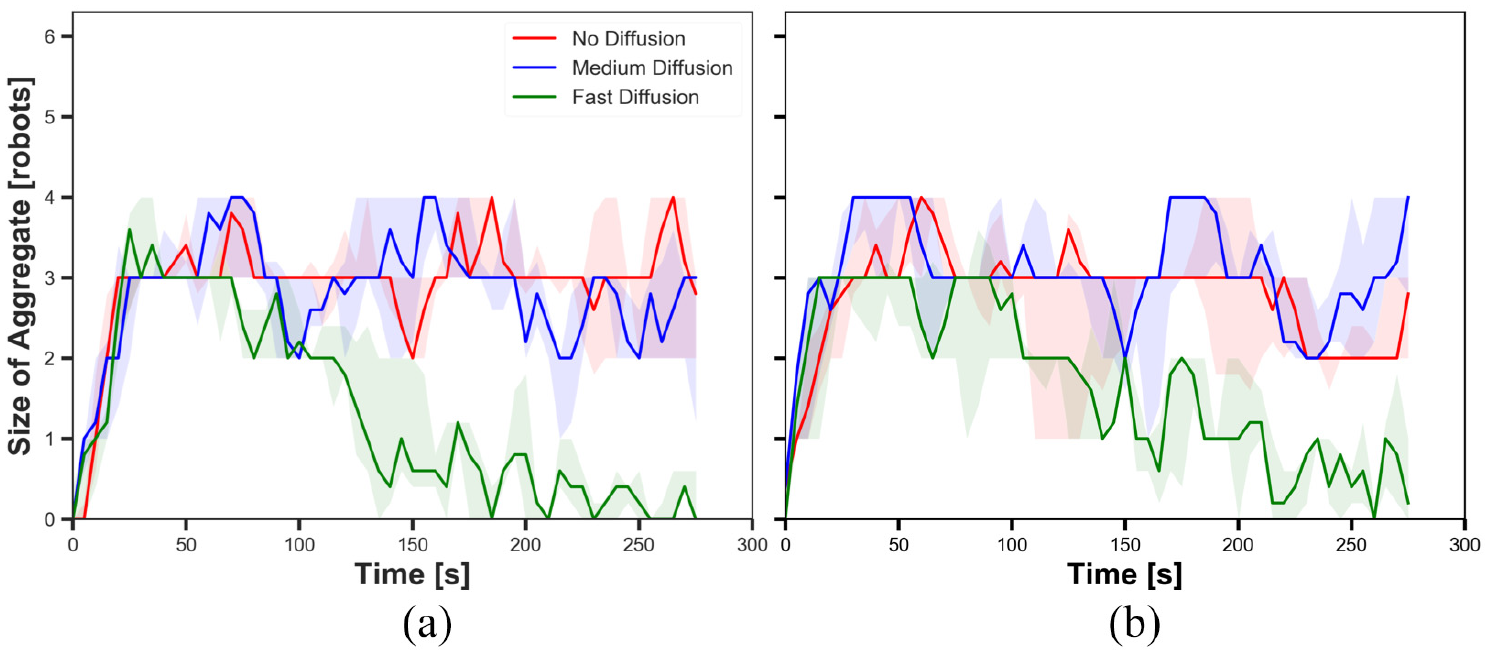
The size of aggregate in experiments with medium cue speed (
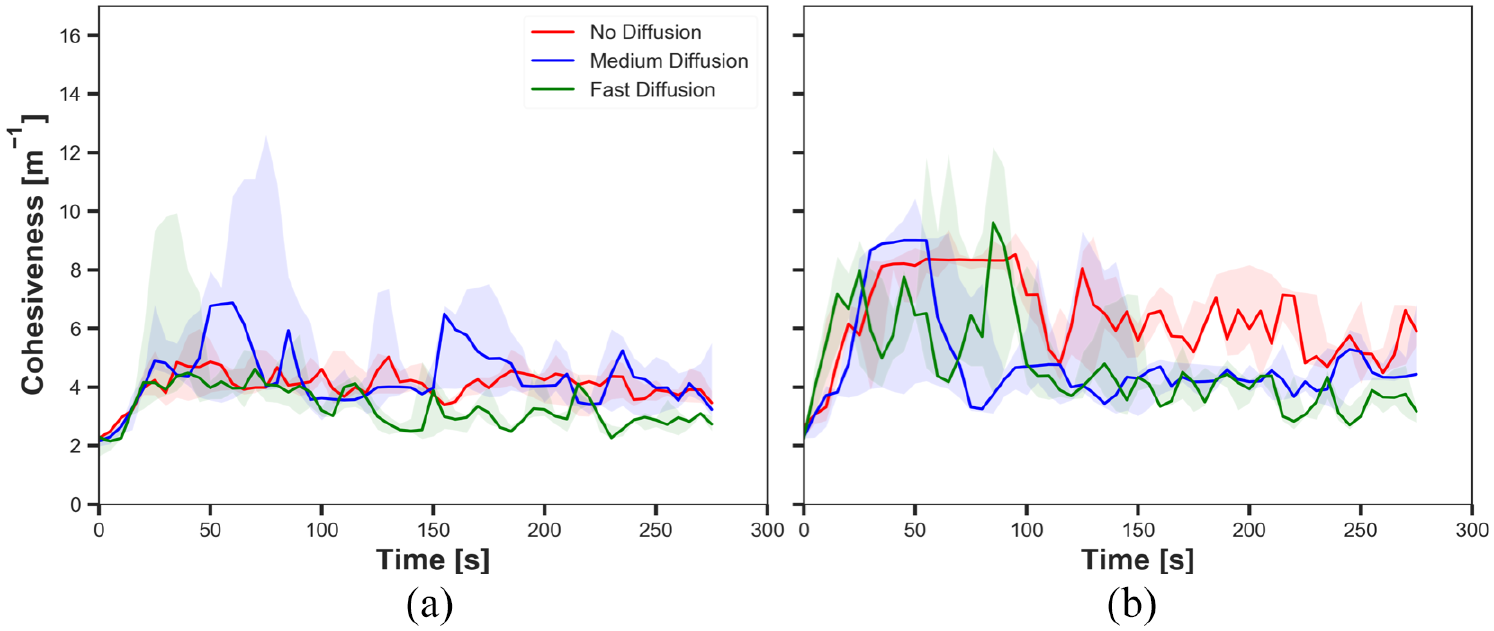
The cohesiveness in experiments with medium cue speed (
The size of aggregate in experiments with medium cue speed (
The cohesiveness in experiments with medium cue speed (
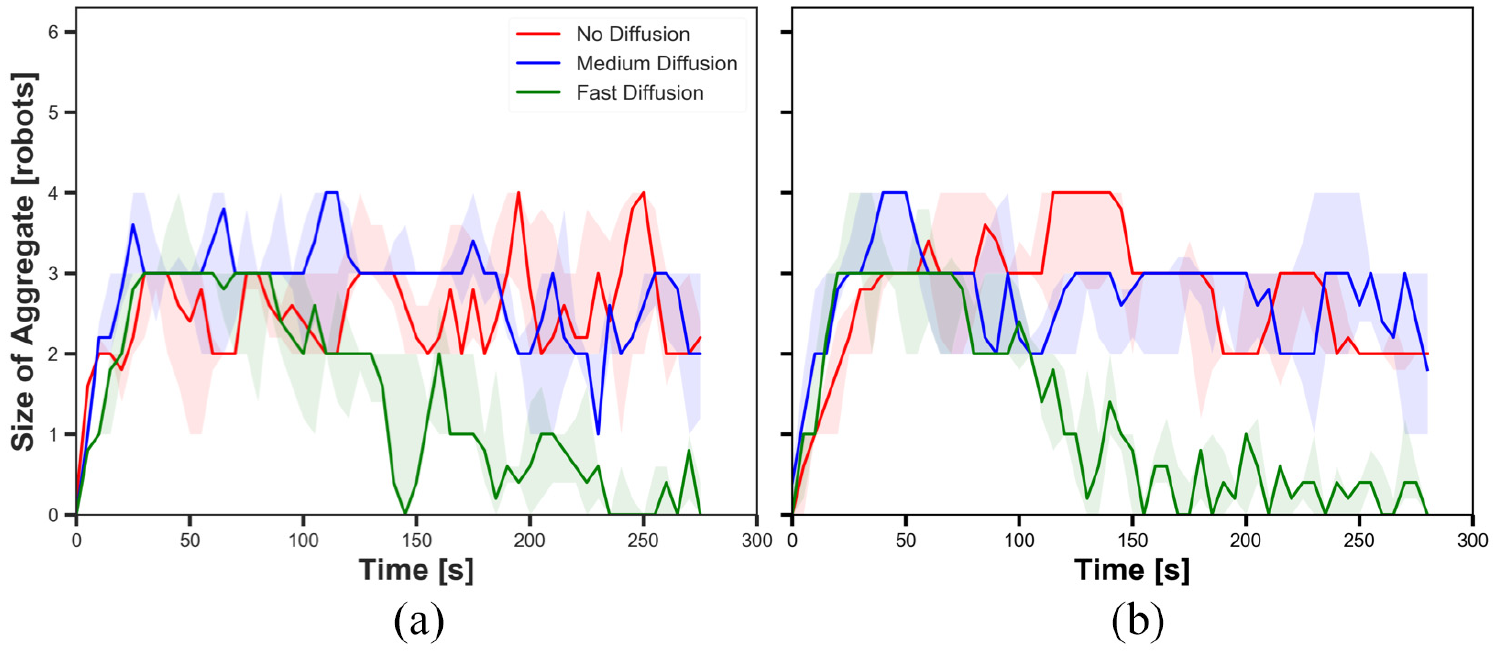
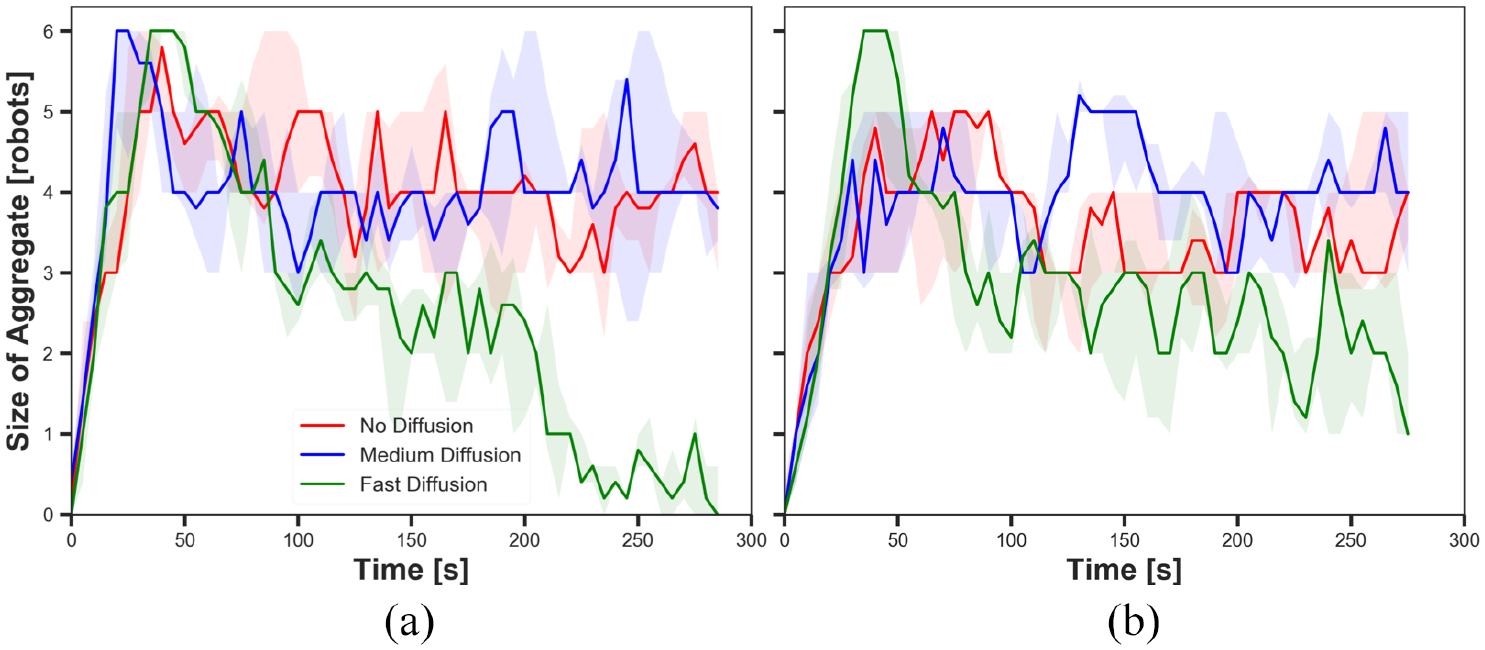
The size of aggregate in experiments with fast cue speed (
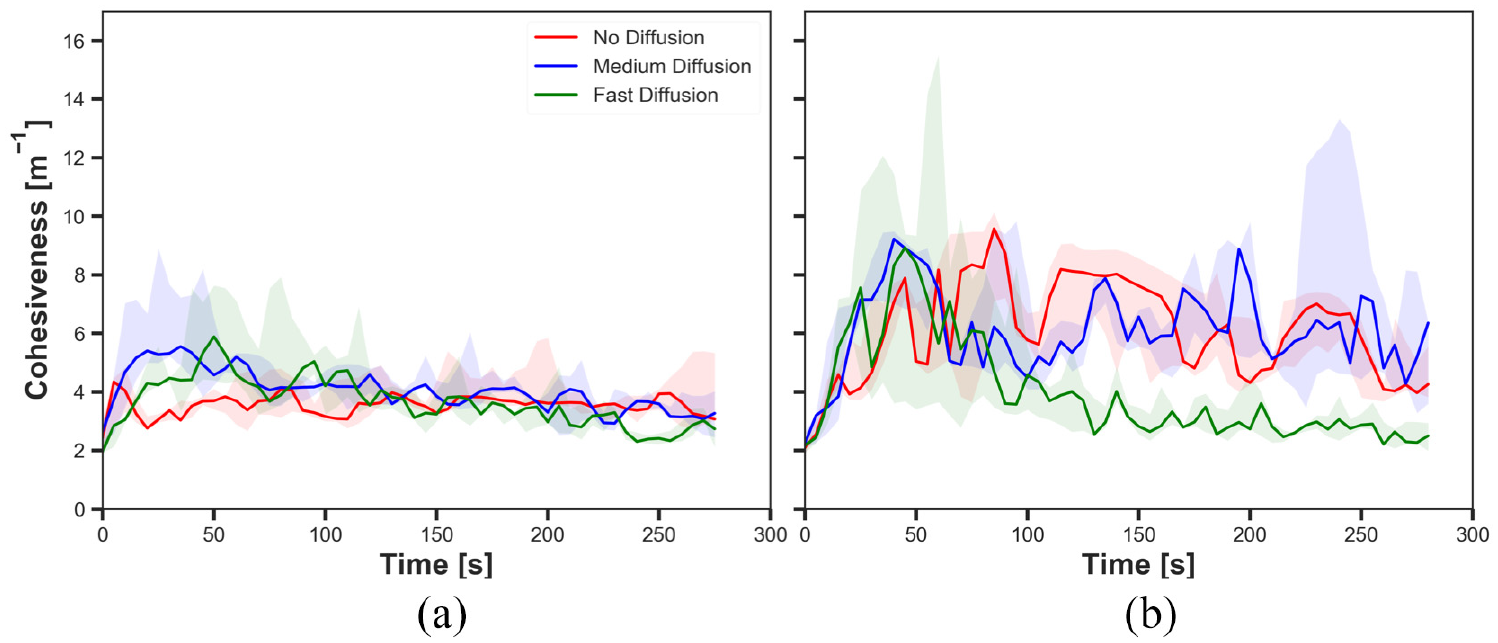
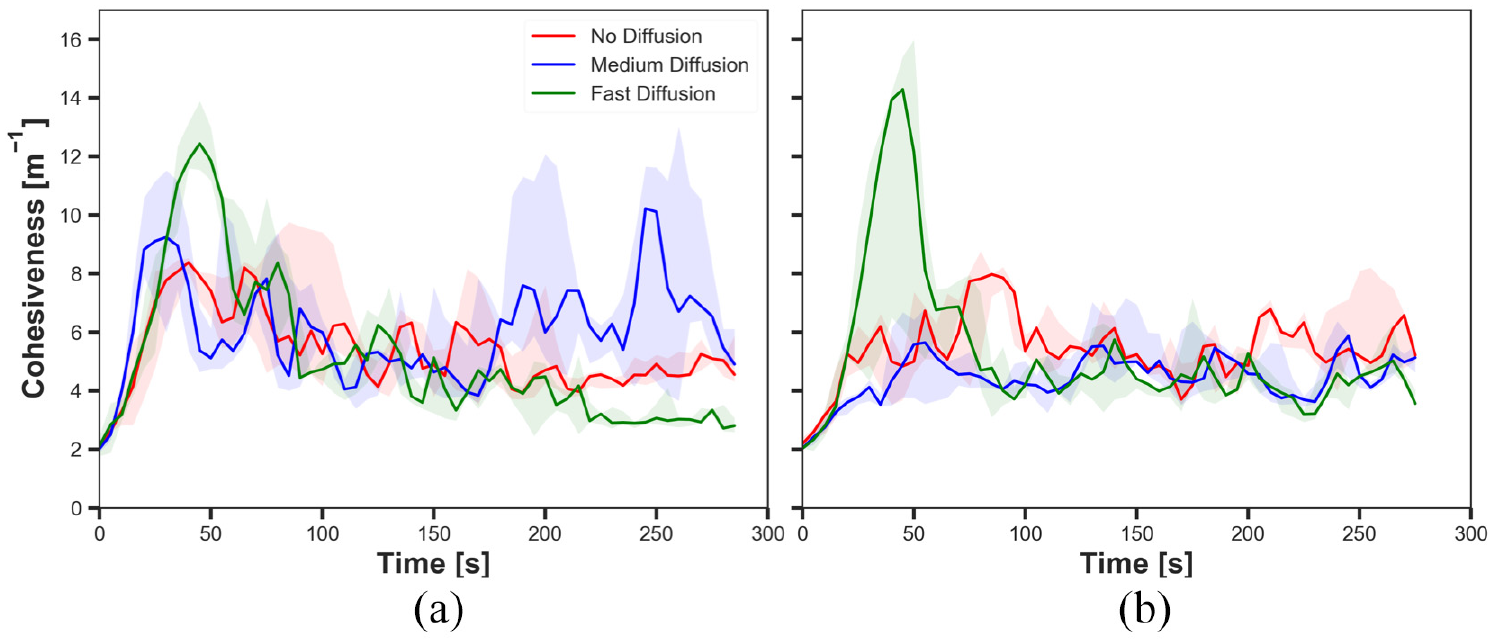
The cohesiveness in experiments with fast cue speed (
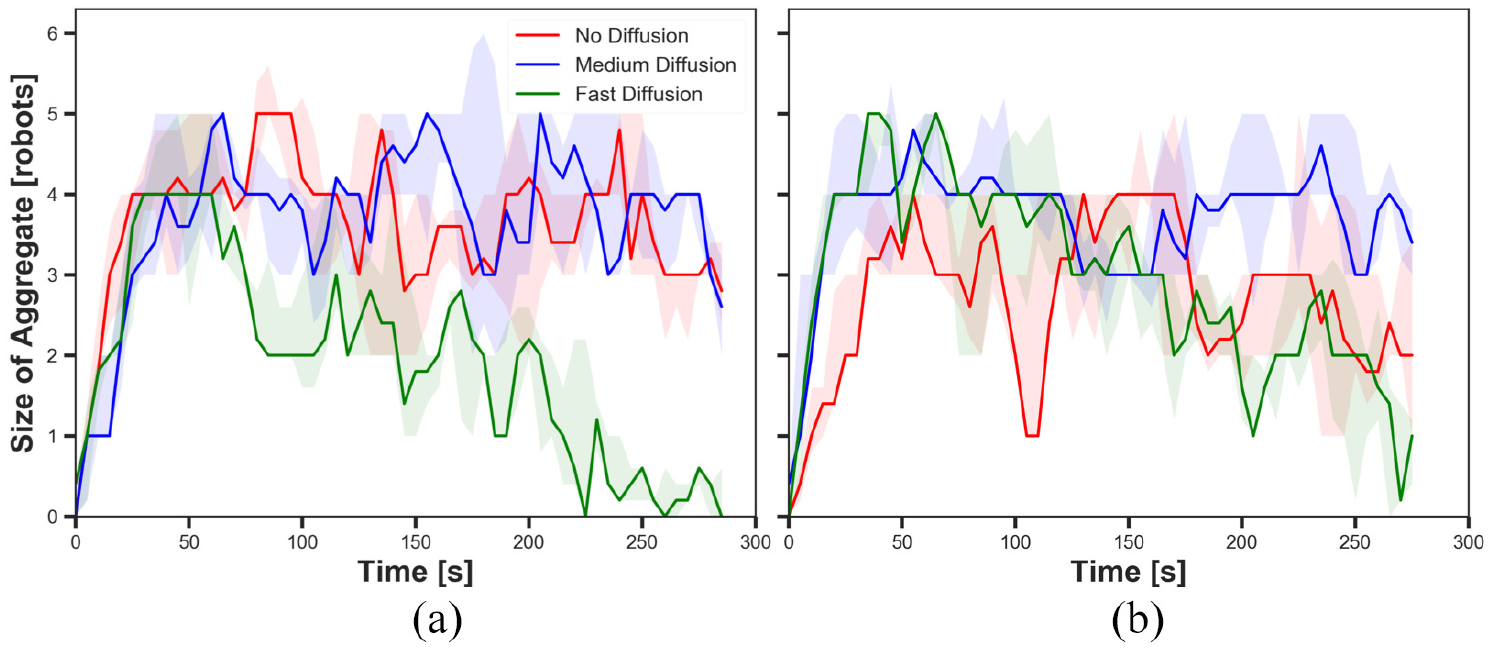
The size of aggregate in experiments with fast cue speed (
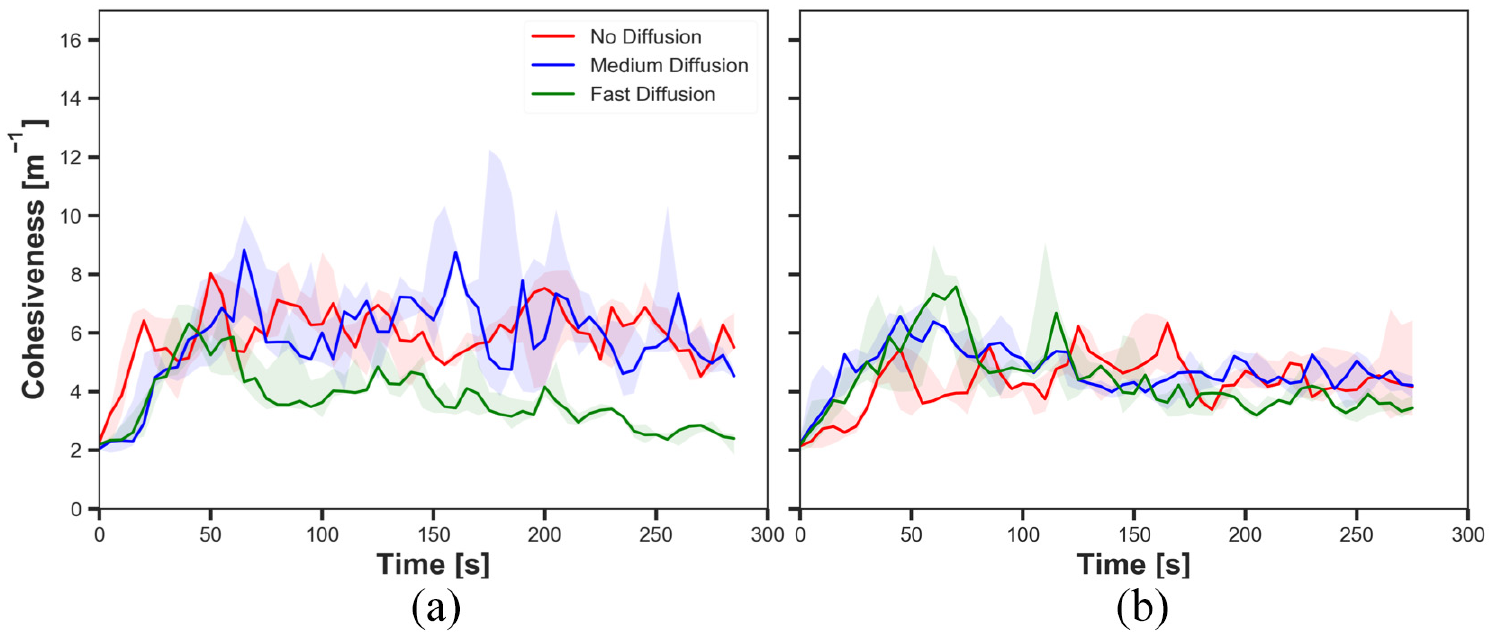
The cohesiveness in experiments with fast cue speed (
5.2.1. Diffusion
The results shown in Figures 9(a), 11(a), 13(a) and 15(a) display the impact of diffusion on the size of aggregate in dynamic cue configuration with two population sizes (
The results shown in Figures 10(a), 12(a), 14(a) and 16(a) demonstrated the impact of diffusion on the cohesiveness in dynamic cue configuration with two population (
The results showed that the effect of diffusion led in decay in the size of aggregate and increase in the cohesiveness as seen in static cue configuration; however, the degree of the effect was smaller in dynamic cue configuration than in static cue configuration. As the cue speed increased, the diffusion effect became less influential. It is because the moving cue dragged the robots with the same direction with the cue and it hampered robots staying closer to the centre of the cue.
5.2.2. Pheromone
The impact of pheromone injection on the size of aggregate is displayed in Figures 9, 11, 13 and 15. In the three Figures 9, 11 and 15, the size of aggregate in experiments with fast diffusion slowly decreased later with
5.2.3. Cue speed
The impact of cue speed was also investigated comparing Figures 9 to 12 with Figures 13 to 16, respectively. The first four figures show the size of aggregate and the cohesiveness in experiments with
The difference observed between the experiments with two different cue speeds was likely to occur because the cue moved away from the robots which were in the waiting phase more quickly with the faster cue speed. The position at which the pheromone was injected was always where the robot was waiting. Therefore, the position of injected pheromone and the cue moved away further from each other in experiments with fast cue speed than medium cue speed. As a result, the greater number of robots tended to stay on the injected pheromone outside the cue with fast cue speed, thereby the decrease in the size of aggregate. The decrease in the cohesiveness by increasing the cue speed was also caused by the same reason why the impact of pheromone injection decreased. Since the cue passed the robots too fast while they were waiting after collision at the centre of the cue, the cohesiveness rapidly decreased especially when it was high. This observation also supports why there was no considerable difference of the cohesiveness between experiments with medium speed and fast speed, where in both cases of no diffusion and medium diffusion was applied.
5.2.4. Statistical analysis
The results from the experiments in the dynamic cue configuration with two different cue speeds were also analysed using ANOVA test (see Tables 2 and 3). The statistical analysis revealed that diffusion and time were two significant factors (
6. Discussion
The observation made from the results suggests that the environmental effects and pheromone injection have an impact on the performance of collective behaviour of robots: size of aggregate and cohesiveness. The impact of the environmental effects and pheromone injection of robot swarm across different population sizes is discussed in detail here.
6.1. Static cue configuration
In static cue configuration, we have found that the trend of swarm behaviour did not noticeably vary between two population sizes (
In the set of experiments without
In the set of experiments with
Results of fully nested ANOVA test.
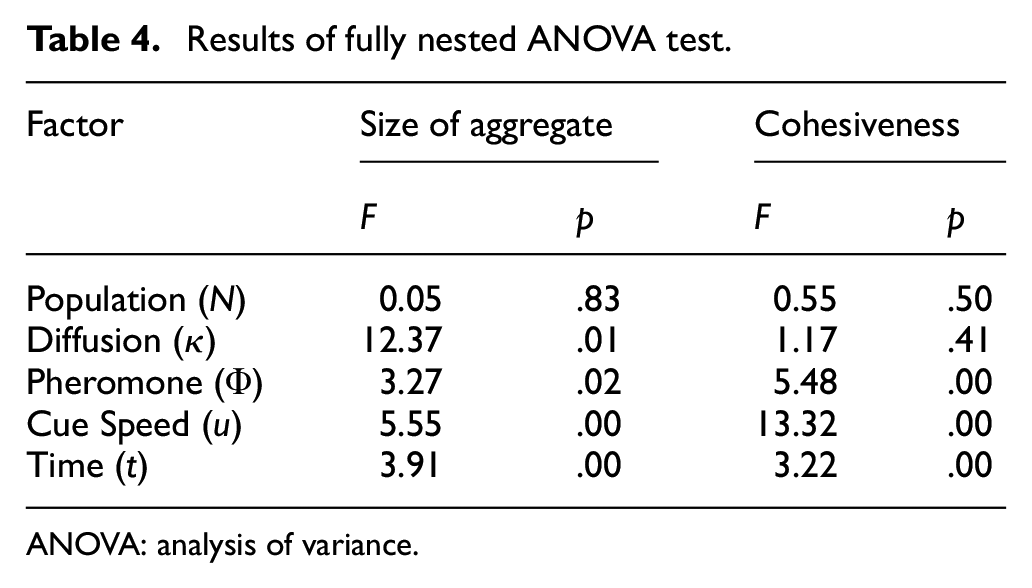
ANOVA: analysis of variance.
6.2. Dynamic cue configuration
In dynamic cue configuration, we investigated the impact of dynamic environment as well as the factors in static cue configuration. As identical in static cue configuration, the population size did not have significant impact on the swarm performance according to the statistical analysis (see Tables 2 and 3).
In the set of experiments without
The effect of pheromone injection in dynamic cue configuration was investigated in the same manner used in static cue configuration. The results demonstrated that the pheromone injection affected the swarm performance. Although the observed impact was identical with static cue configuration, the observation in dynamic cue configuration was not as clear as in static cue configuration. In Figures 9 and 13, the trend of the size of aggregate in experiments with
Figure 17 shows the overall experiments designed to show the impact of cue speed primarily. The results depicted that the cue speed did not deteriorate the swarm performance. Interestingly, in experiments with the medium cue speed, the size of aggregate reached the highest regardless of diffusion and pheromone injection. This observation suggests that the gradient made by moving pheromone with a moderate speed leads the highest responsiveness on pheromone for robots. This guidance effect using the gradient in pheromone trails was reported in Nieh et al. (2004). As well as the observation of maximised dragging effect with the medium cue speed, it is worth noting that the size of aggregate with the fast cue speed fluctuated shown in Figure 17(d). It was caused by the definition of the cue location. As mentioned in ‘Results’ section, it is shown that the swarm aggregate was formed around the cue, rather than roaming. Therefore, the swarm was still robust with this extreme environmental effect. The statistical analysis showed the cue speed was significantly influential to the robot swarm. This phenomenon that the increase in cue speed sustains the size of aggregate against the environmental effects was also reported in Na et al. (2019).
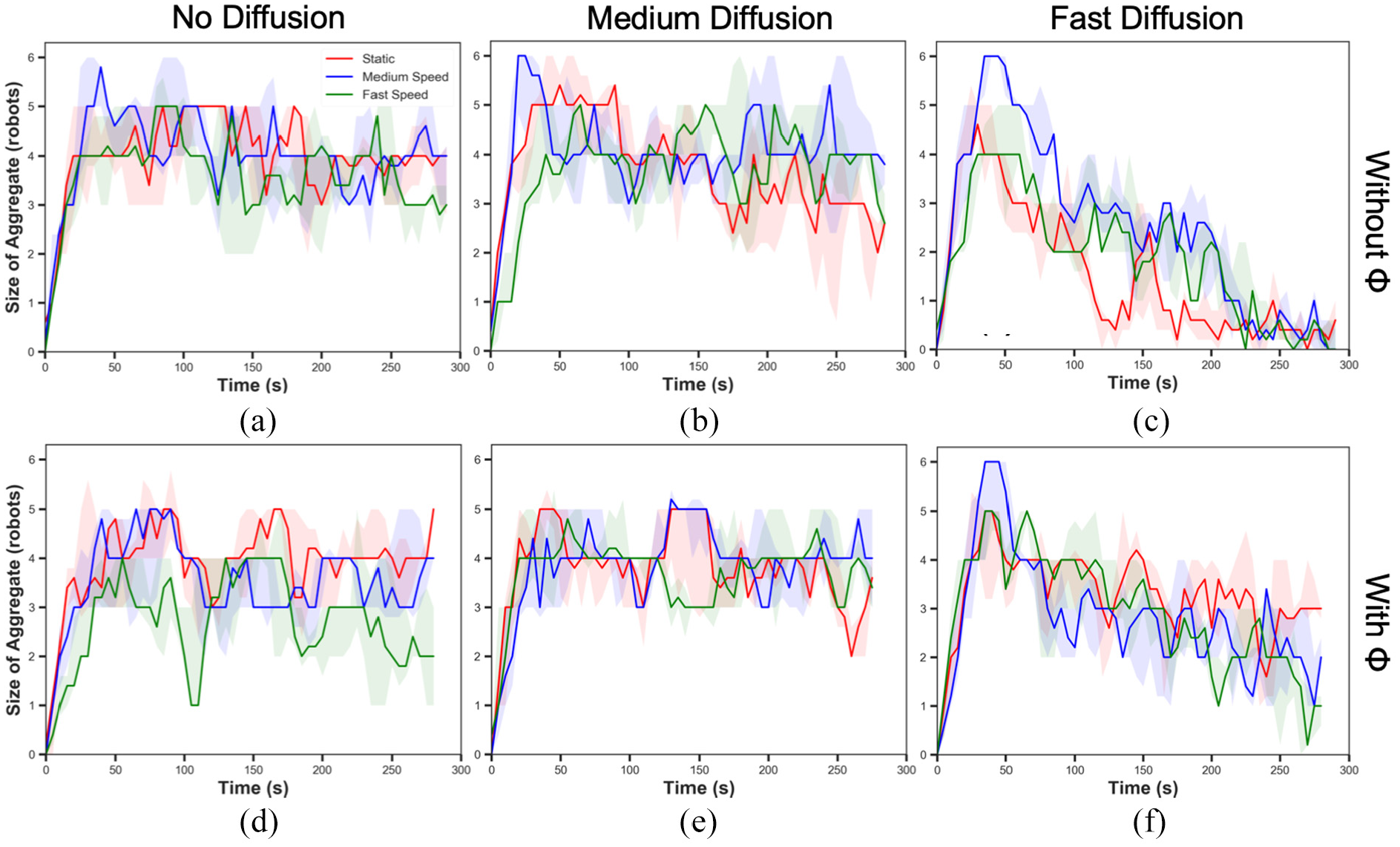
The size of aggregate in experiments with different cue speeds (u ∈ {0, 0.1, 0.2} cm/s) without pheromone (Φ) injection - (a), (b) and (c) and with pheromone (
7. Conclusion
In this study, we proposed a state-of-the-art artificial pheromone system, which, unlike the previous systems, allows efficient and realistic emulation of environmental effects on the pheromone distribution. We demonstrated that emulation of realistic spatio-temporal development of pheromone, involving pheromone diffusion and advection, can bring new insights into the interplay of the swarms, released pheromones and the environment. One of the interesting findings is that the ability of the swarm to release pheromones counters adverse effects of moving cue, which can displace environmental cues. In future works, we will realise scenarios with more pheromone types implemented by different colours. Using pheromones with different diffusion, evaporation and advection rates will allow investigation of richer pheromone-swarm-environment interactions emulating complex biological swarms in the real world. Since the effect of time is paramount to the swarm behaviours, we will combine the pheromone field with the system for on-the-fly recharging systems for swarm robotics.
Footnotes
Acknowledgements
The first author would like to express special thanks to the University of Manchester for supporting his PhD study with the President’s Doctoral Scholar Award.
Handling Editor: Geoff Nitschke, University of Cape Town, South Africa
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This article was supported by the UK EPSRC projects RAIN (EP/R026084/1), RNE (EP/P01366X/1) and OP VVV funded project CZ.02.101/0.0/0.0/16_019/0000765 ‘Research Center for Informatics’ and CSF project 17-27006Y STRoLL.