
Abstract
A 10-y-old, castrated male Boxer dog that was born and had lived in Missouri without any travel history to other states, except for a few trips to Kansas, was presented with a distended abdomen and declined health. Ultrasonographic examination revealed a large hepatic mass, and the dog was euthanized. A postmortem examination revealed that the left liver lobes were largely replaced by a white-to-tan multilobular mass with a cobblestone surface. The lesion also involved the diaphragm. Histologically, hepatic architecture was effaced by large areas of necrosis with numerous, ≤0.2-cm, cystic structures that stained positively with periodic acid–Schiff stain and contained calcareous corpuscles. Gross and microscopic hepatic lesions were compatible with alveolar echinococcosis (AE) caused by Echinococcus multilocularis. PCR examination confirmed E. multilocularis, and results from genotyping were consistent with the E4 haplotype. To our knowledge, this is only the second canine AE case and the third pet dog that has been confirmed to be infected by E. multilocularis in the contiguous United States. E. multilocularis is a serious health risk for both pet dogs and humans.
Keywords
In January 2018, a 10-y-old castrated male Boxer dog was presented to a veterinary clinic in Columbia, MO, USA with a distended abdomen and declined health. The dog was born and had lived almost all of its life in central Missouri, with a few trips to Kansas. The dog had been fed dry kibble. The owner reported that the dog had occasional vomiting and inappetence as early as the summer of 2016. Although the clinical signs flared up several times over the past 1.5 y, the dog was considered to be otherwise bright, alert, and responsive. Ultrasonographic examination of the distended abdomen revealed a large irregular mostly hypoechoic hepatic mass with possible extension to the diaphragm. The owner elected to euthanize the dog given the declined health state and the hepatic mass.
An autopsy was declined; however, the owner consented to a postmortem examination limited to a midline abdominal incision to reveal the nature of the hepatic mass. The left liver lobes were largely replaced by a white-to-tan multilobular mass with a cobblestone surface. A similar change was noted along the diaphragm adjacent to the hepatic mass. Other abdominal viscera were grossly unremarkable. Only an affected liver sample was collected and placed in 10% neutral-buffered formalin for histologic examination. Grossly, the lesion in the formalin-fixed liver specimen consisted of many small (~0.1–0.2 cm) cysts, giving a sponge-like appearance on cut surfaces.
After routine histologic processing, H&E-stained sections were prepared and examined. Normal liver architecture was almost completely effaced by large areas of necrosis with numerous, ≤0.2-cm, cystic spaces, which contained ≤40-µm thick, pale eosinophilic and acellular, cystic and ribbon-like structures (Fig. 1A) that stained positively with periodic acid–Schiff stain (Fig. 1B), consistent with the laminated layer of hydatid cyst walls. The hydatid cysts often contained a few ≤10-μm, pale basophilic calcareous corpuscles (Fig. 1C). The laminated layer of the cyst walls was denuded of the germinal layer in most areas; however, remnants of the nucleated germinal layer remained in a few places (Fig. 1D). The areas of necrosis were often mineralized and were occasionally surrounded by granulomatous inflammation and fibrosis with small numbers of foreign body–type multinucleate giant cells and eosinophils. No protoscolices were observed in the tissue sections examined. Gross and histopathologic findings were compatible with alveolar echinococcosis (AE) caused by infection with the larval stage of Echinococcus multilocularis.
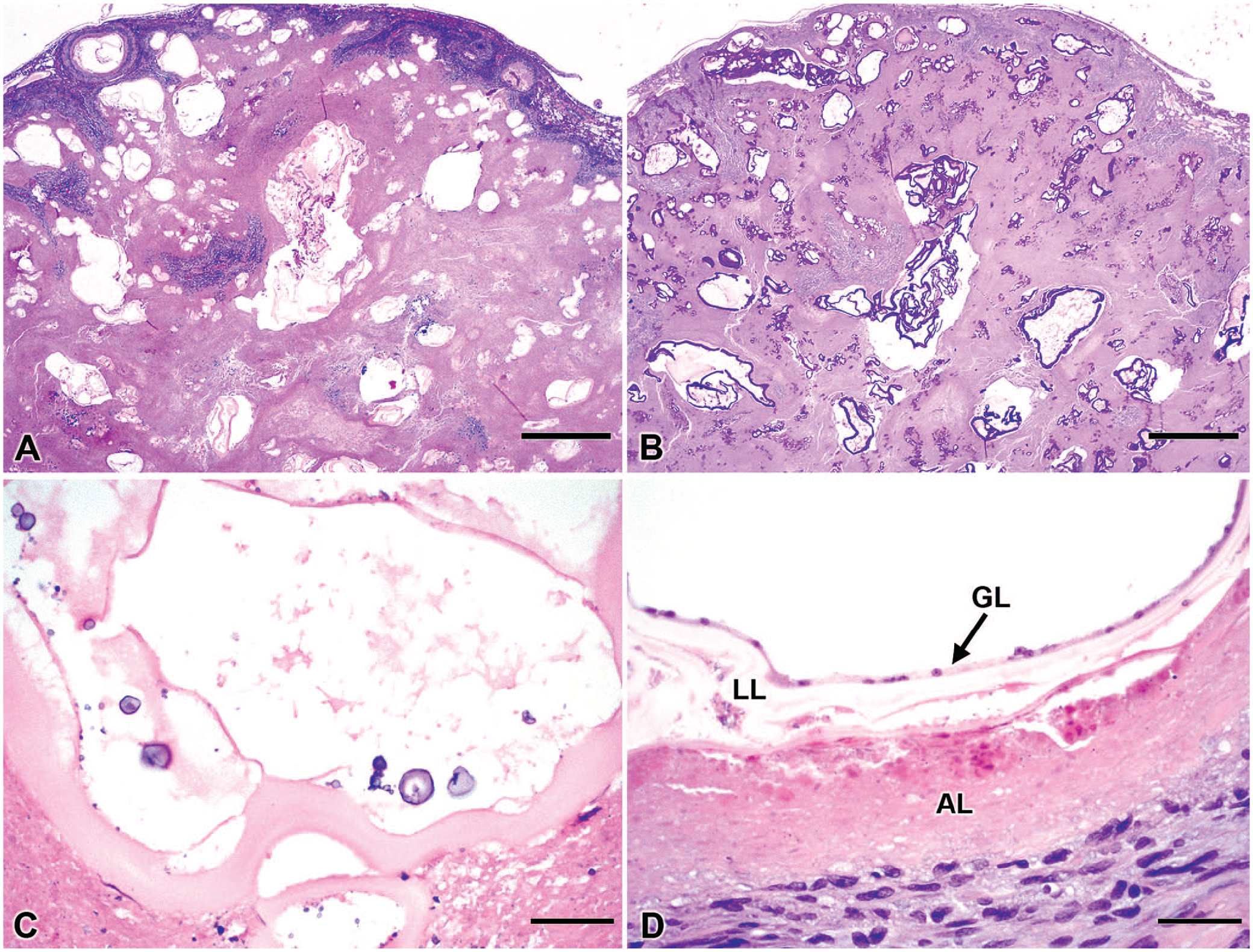
Hepatic alveolar echinococcosis in a Boxer dog.
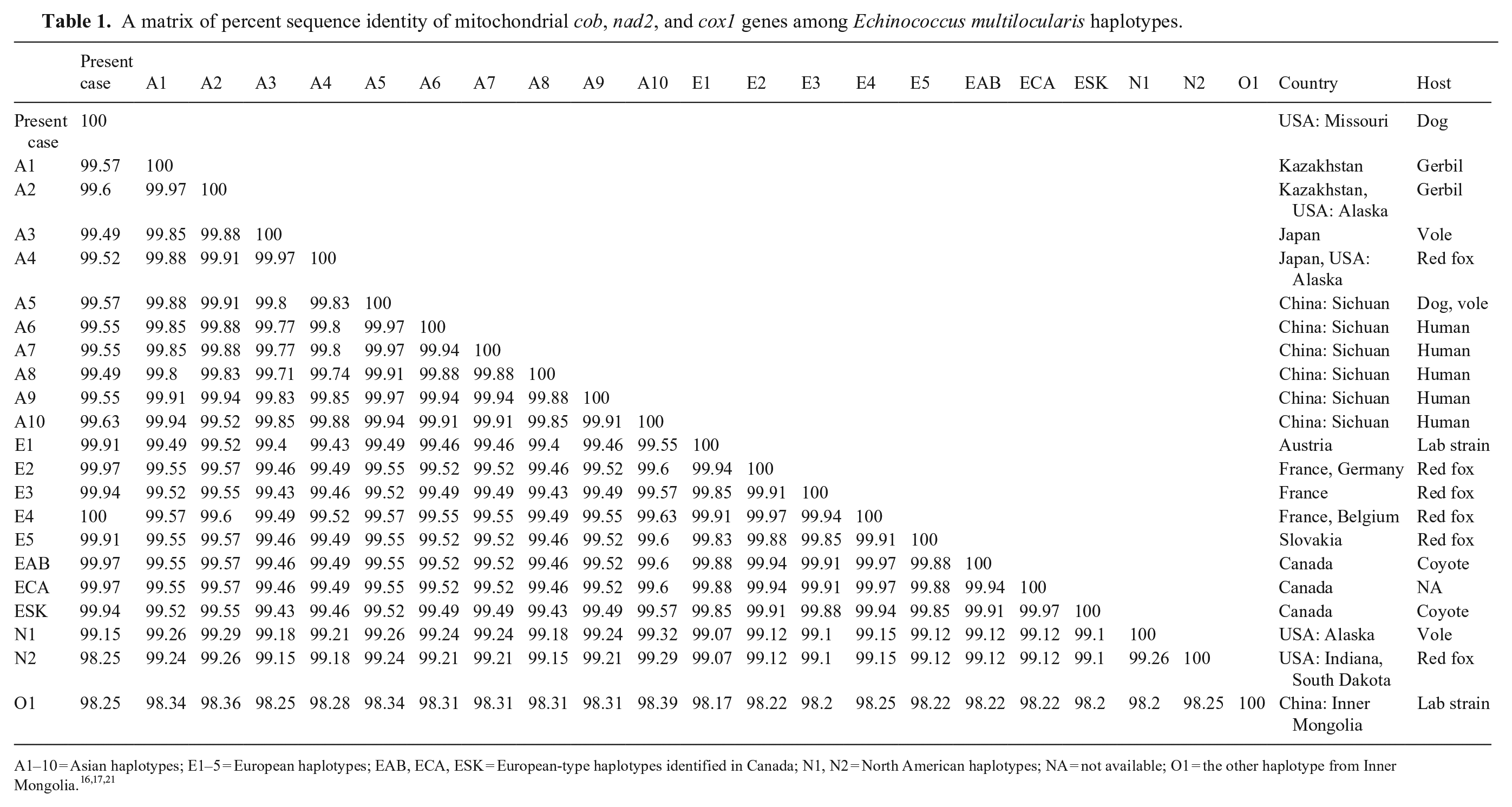
From the remaining formalin-fixed liver specimen, DNA was extracted at the National Institute of Infectious Diseases, Tokyo, Japan. Mitochondrial genes cytochrome b (cob), NADH dehydrogenase subunit 2 (nad2), and cytochrome c oxidase subunit 1 (cox1) were amplified (KOD FX Neo DNA polymerase; Toyobo) as described previously. 15 Sequence analyses of these 3 genes have identified >20 haplotypes of E. multilocularis that have been grouped in 4 basal clades (i.e., Europe, Asia, North America, Inner Mongolia).7,8,11,16,21 We employed validated E. multilocularis species–specific primers.15,21 The PCR products of cob, nad2, and cox1 were directly sequenced in both directions; nucleotide identities were calculated with SIAS (http://imed.med.ucm.es/Tools/sias.html), and the sequences were aligned (MEGA X v.10.2.6; https://www.megasoftware.net/). 14 Sequence data for the cob, nad2, and cox1 of our case (GenBank LC645085, LC645086, and LC645087, respectively) were 100% identical to E. multilocularis sequences from the E4 haplotype, one of the European haplotypes, detected in red foxes in France and Belgium (AB461413 21 ; Table 1). A phylogenetic tree was generated using the maximum-likelihood method (HKY+G model of nucleotide substitution) with E. granulosus (AF297617, AF297617, AF297617) as an outgroup (Fig. 2).
A matrix of percent sequence identity of mitochondrial cob, nad2, and cox1 genes among Echinococcus multilocularis haplotypes.
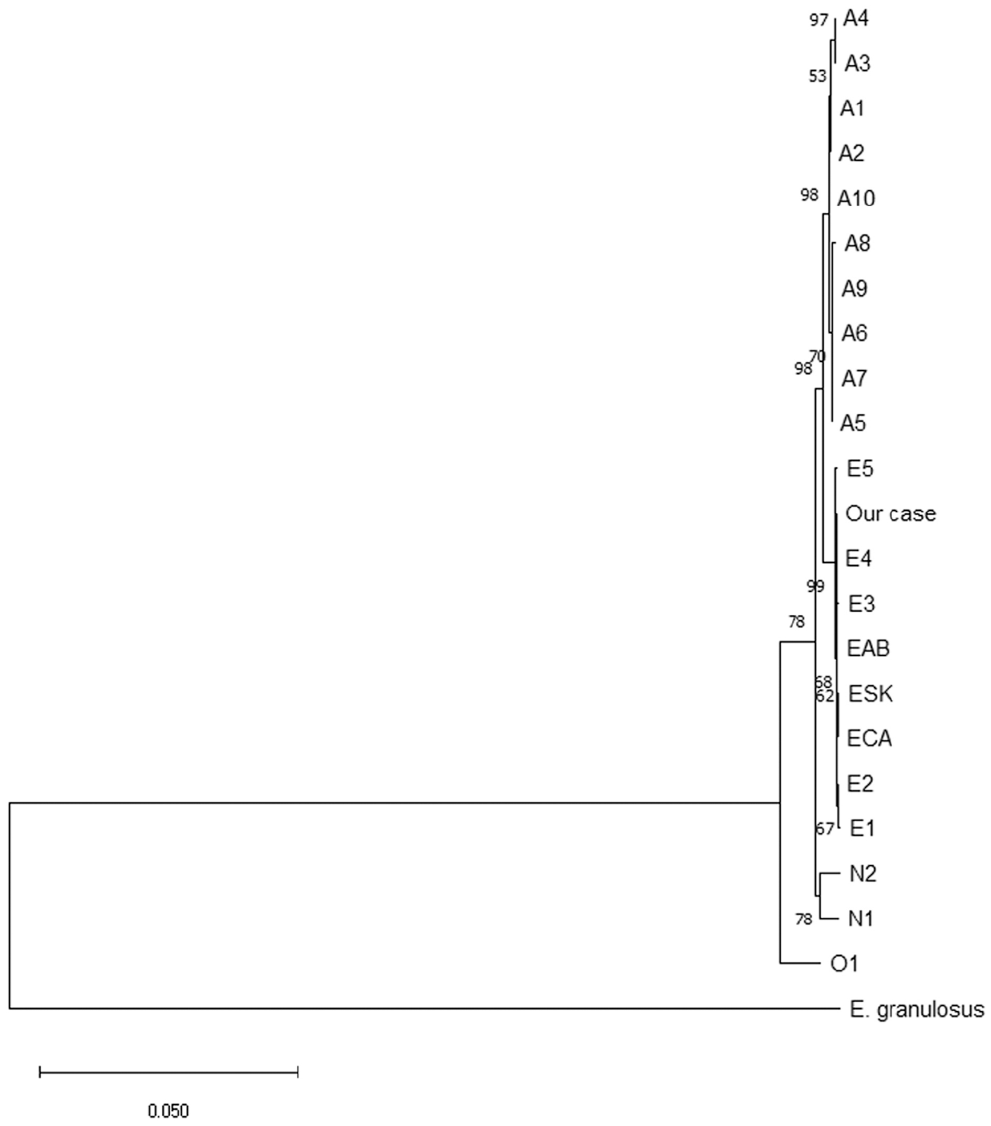
Maximum-likelihood phylogenetic tree of Echinococcus multilocularis haplotypes with E. granulosus as an outgroup.
Echinococcosis is an important zoonotic disease around the world.2–4 The 2 major Echinococcus spp. of medical importance are E. granulosus and E. multilocularis. The life cycles of E. granulosus and E. multilocularis, which are the causative agents of human cystic echinococcosis (CE) and AE, respectively, involve canids as definitive hosts.2–4 CE caused by infection with the larval stage of E. granulosus is most common in areas in which dogs have access to infected livestock offal, particularly from sheep.4,19 In the United States, a small number of autochthonous CE cases have been recognized in Alaska and a few western states. 19 One pediatric CE case has been reported in Missouri; however, the infection was probably not acquired in the Midwest because the patient lived in Alaska before moving to Missouri. 10
AE is widely distributed in the northern hemisphere.2–4,19 Small rodents act as the natural intermediate hosts for E. multilocularis.2–4 Although the main definitive hosts for E. multilocularis are wild canids (e.g., foxes and coyotes), dogs can also be definitive hosts.2–4,15 Humans and dogs can become aberrant intermediate hosts and develop AE when eggs of E. multilocularis are ingested accidentally. Historically, 2 endemic regions of E. multilocularis were recognized within North America: the Northern Tundra Zone of Alaska and the Canadian Arctic, and the North Central Region (NCR), including 3 Canadian provinces (Alberta, Saskatchewan, Manitoba) and 13 U.S. states (Illinois, Indiana, Iowa, Michigan, Minnesota, Missouri, Montana, Nebraska, North Dakota, Ohio, South Dakota, Wisconsin, Wyoming).2,4,17 In the Canadian part of the NCR, E. multilocularis was detected in 10 of 43 (23%) coyotes in Manitoba in the 1970s, 23 21 of 91 (25%) coyotes in Alberta in 2009–2011, 1 and 9 of 10 (90%) coyotes in Saskatchewan in 2010–2012. 9
Although E. multilocularis was not known to occur in British Columbia and Ontario, these 2 provinces are now recognized endemic regions of Canada based on the information published since 2009. The first reported case of AE in a pet dog in North America was from British Columbia in 2009. 11 Further investigation revealed the establishment of E. multilocularis in the local wildlife in British Columbia. 7 There have been several canine cases of AE reported in Ontario since 2012.13,25 An investigation of fecal samples collected from 460 wild canids (416 coyotes, 44 foxes) in Ontario during 2015–2017 revealed that 23% were positive for E. multilocularis. 13 These findings from British Columbia and Ontario suggest that the range of E. multilocularis is either shifting and/or is much wider than we anticipated previously. There were 12 human AE cases between 2001 and 2014, and at least 6 more human AE cases since 2016, in Canada.16,17
In the U.S. part of the NCR, the prevalence of E. multilocularis in definitive wild canid hosts can be estimated at ~15%, with the highest prevalences (>20%) in South Dakota, Nebraska, Indiana, and Ohio, based on a report on adult cestode recovery at autopsy. 4 Central Indiana had been considered the southern limit; Ohio and Michigan had been considered the eastern limit for this parasite in the contiguous United States based on published data.4,18,27 Evidence of endemicity of E. multilocularis in Missouri was based on unpublished data in foxes 24 until a report of the first confirmed case of intestinal echinococcosis (IE) in a juvenile pet dog in Missouri in 2020, 15 which was the first detection of a European strain (E4 haplotype) of E. multilocularis in a pet dog in the contiguous United States. 15 In addition, a canine AE case had been reported in an adult pet dog in Virginia. 30 Molecular investigation with mitochondrial gene nad2 revealed that the E. multilocularis in the Virginia dog was consistent with European strains. 30 Although, outside Alaska, only one autochthonous human AE case has been reported in the United States in 1979, 6 E. multilocularis might have spread in a much wider region of the contiguous United States than thought previously.
Our understanding of the global distribution of genetic variants of E. multilocularis was initially established by identifying mitochondrial DNA haplotypes in 76 geographic isolates; designated as E1–5 (European haplotypes), A1–10 (Asian haplotypes), N1 and N2 (North American haplotypes), and O1 (other haplotype in Inner Mongolia). 21 In Alaska, 2 Asian mitochondrial haplotypes (A2 and A4), 1 North American haplotype (N1), and a unique EmsB microsatellite profile, have been detected in wildlife.12,21 EmsB genotyping is based on the analysis of complex PCR profiles defined by the size, number, and height of the peaks obtained after amplification and separation of PCR products using capillary electrophoresis. 12 The N2 haplotype has been considered endemic in the NCR based on genotyping of limited samples.21,29 Molecular investigations with mitochondrial genes cob, nad2, and cox1 stemming from a canine AE case in British Columbia, Canada, in 2009 led to the discovery of a new European strain of E. multilocularis, designated as the BC1 haplotype in a wild coyote population in British Columbia, and 3 more new European strains: ECA, EAB, and ESK haplotypes in local animals in the Canadian NCR.7,8,11,16 Furthermore, 17 new haplotypes of E. multilocularis were detected in wild animals in Canada through sequencing of nad1. 9 Detection of European strains from canine cases in the contiguous United States implies that there might be higher than expected heterogeneity of E. multilocularis in the contiguous United States, similar to observations in Canada.
Because of the presence of fox and coyote populations in suburban and urban areas in the United States, there is growing concern of spillover from infected foxes and coyotes and potential human exposure from the contaminated environment. Pet dogs can develop IE and become definitive hosts by consuming infected wild rodent intermediate hosts 19 ; pet dogs become aberrant intermediate hosts through ingestion of E. multilocularis eggs. After egg ingestion, alveolar hydatid cysts develop in internal organs, most commonly the liver, leading to a slowly progressive and serious health condition.3,22 From the liver, larvae can spread by direct extension to adjacent organs, 22 as in our case. Germinal cells of the larvae can also disseminate via lymph or blood vessels to distant organs. 22 Cases of canine AE in the contiguous United States, along with our previously reported IE juvenile dog case, 15 are not only of pathology interest, but also of significant epidemiologic and public health concern.
Dogs can be a source of human infection when they are definitive hosts (IE); dogs can be considered sentinels of environmental contamination with E. multilocularis eggs when they serve as aberrant intermediate hosts (AE). 4 Canine AE cases can be considered indicators of establishment of E. multilocularis among local predator-prey communities with environmental contamination. We suspect that our case became infected with E. multilocularis by ingesting eggs from the feces of a local wild canid with the intestinal infection. Although concurrent infections of the intestine and the liver with adult and larval stages of E. multilocularis have been reported in dogs, 3 echinococcosis was not suspected at postmortem examination, and hence the small intestine and feces were not examined for adult worms in our case. IE should be ruled out even in cases of canine AE, and people in close contact should be investigated for exposure to the parasite in cases of concurrent intestinal infection. It is important to raise awareness of E. multilocularis among animal and human health professionals and dog owners in the United States.
In IE cases, veterinarians should be aware that infection in dogs can be missed because, on fecal flotation, ova of E. multilocularis are indistinguishable morphologically from those of Taenia spp., the more common tapeworms in dogs in the United States. 5 Proglottids of Echinococcus spp. are so small (~1-mm long) that they can be overlooked macroscopically. In addition, they are fragile and easily digested while passing through the intestines, leaving only ova in feces. 28 It is unknown if canine echinococcosis is truly a rare disease in the contiguous United States or if cases are being misdiagnosed or overlooked.
It is important to consider molecular characterization of taeniid ova detected in feces and immediately treat dogs with praziquantel, an approved treatment for IE. Diagnosis of hepatic AE can be a challenge because it is a slowly progressive disease and distinguishing AE lesions in the liver from neoplastic liver lesions by imaging modalities is difficult. 20 In humans, the diagnosis of AE relies mainly on findings by imaging modalities, supported by positive serologic tests. To our knowledge, serologic testing for E. multilocularis is not available at veterinary diagnostic laboratories in the United States. Although serology is of value for diagnosing canine AE, in conjunction with imaging findings, possible cross-reactions with Taenia spp. should be considered. 26 To our knowledge, PCR testing for E. multilocularis for veterinary species is not provided by veterinary diagnostic laboratories in the United States. Development and validation of a test for E. multilocularis in veterinary species among U.S. veterinary diagnostic laboratories is desirable. Further investigation into the distribution, mitochondrial haplotyping, and pathogenicity of E. multilocularis in the contiguous United States is warranted.
Footnotes
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Our work was supported partially by a grant to Yasuyuki Morishima (21HA2001) from the Ministry of Health, Labor and Welfare, Japan.