
Abstract
Infectious diseases have contributed to the decline in the health of koala (Phascolarctos cinereus) populations in the wild in some regions of Australia. Herein we report the development and validation of 2 multiplex real-time PCR (rtPCR) panels for the simultaneous detection of Mycoplasma spp., Ureaplasma spp., Bordetella bronchiseptica, and Chlamydia, including speciation and quantification of Chlamydia, in ocular, reproductive, and nasal swab samples in addition to semen and male urogenital and reproductive tissues, from koalas. Each rtPCR panel was developed for use as a single-tube reaction using pathogen-specific primers and fluorescently labeled probe sets. DNA extracted from reference strains and isolates was used for validation of sequence gene targets for the multiplex rtPCR panels. Each panel was shown to be sensitive and specific in detecting and differentiating the bacterial pathogens. The multiplex rtPCR panels were used to screen clinical samples from free-ranging and hospitalized koalas for multiple pathogens simultaneously. The multiplex rtPCR will improve turnaround time compared to individual-pathogen rtPCR methods used, to date, for confirmation of diagnosis and will provide the wildlife clinician with the ability to make treatment decisions more rapidly.
Keywords
Introduction
Koala (Phascolarctos cinereus) populations throughout Australia, particularly in southeastern Queensland and New South Wales, are under significant threat of collapse, leading to localized extinction of populations. 25 The decline in population numbers can largely be attributed to habitat loss and fragmentation, 19 motor vehicle trauma,8,28,29 domestic dog attacks, 17 and infectious diseases.15,25,35 In Australia, there has been a 24% decline of koala numbers over the past 20 y, with Queensland koalas being considered the most vulnerable, declining by up to 53% over this same period. 1 Two of the most widely recognized disease agents impacting the koala are Chlamydia, an obligate intracellular bacterium, and species Koala retrovirus (KoRV). 31 It has also been proposed that immunomodulation induced by KoRV is contributing to the high rates of chlamydiosis observed in koala populations.6,30 Although chlamydiosis in the koala is well documented as a major contributor to the decline of population numbers in large parts of eastern Australia, disease prevalence varies among these populations and infected animals may not show clinical signs.9,25,37
Chlamydia pecorum and Chlamydia pneumoniae are known infectious agents of the koala, with C. pecorum being the more pathogenic of the 2 species.7,13 Surveys of wild koala populations in several Australian states have also revealed highly variable infection rates of 0–100% for C. pecorum and 0–53% for C. pneumoniae.3,23,25 C. pecorum infects the ocular, urinary, and genital mucosae leading to blindness, cystitis, and infertility, respectively.2,35 Infections by C. pecorum often produce no clinical signs, especially in the reproductive tract, and therefore go unnoticed during population surveillance. 25 C. pneumoniae is a common human and animal pathogen associated with a wide range of upper and lower respiratory tract infections. 20 Clinical signs of C. pneumoniae infection include sneezing, coughing, dyspnea, rhinitis, and in severe cases, pneumonia.7,10,36 Together, chlamydial infections contribute not only to the overall health decline of koala populations, but also to reduced reproductive rate.4,12,22
In addition to Chlamydia spp., a number of other bacterial organisms, including Mycoplasma spp. and Ureaplasma spp., may contribute to population decline, but their epidemiology and pathogenicity have yet to be fully characterized in the koala. In humans and many animal species, Mycoplasma spp. and Ureaplasma spp. have been responsible for lesions similar to those of chlamydiosis, including expression of clinical disease in urogenital, ocular, and respiratory infections. 34 To date, the prevalence and pathogenicity of these 2 bacteria in koala populations is unknown.
In contrast, Bordetella bronchiseptica infections have been well documented in captive koala populations.2,5,18 In the koala, B. bronchiseptica respiratory infections exhibit clinical signs similar to those of C. pneumoniae, such as mucopurulent nasal discharge, sneezing, and/or coughing. 2 Respiratory disease caused by B. bronchiseptica infection is extremely serious and severe and has been known to affect individual animals as well as groups; if left untreated, infection rapidly results in mortality. 2 Unlike C. pneumoniae, a vaccine for B. bronchiseptica is currently available for koalas. 2
To better assess the prevalence and understand the epidemiology of infectious diseases in wild koala populations, rapid and reliable screening methods for a range of common koala disease agents are required. 14 Multiplex real-time PCR (rtPCR) assays provide a fast, highly specific DNA-based tool for the simultaneous amplification of more than one target sequence in a single reaction, which is becoming a rapid, economical, and convenient means in research or clinical diagnostic laboratories to simultaneously detect multiple pathogens causing similar signs while minimizing sample handling and risks of cross-contamination. 21 We report the development and validation of 2 novel multiplex rtPCR panels to simultaneously screen for C. pecorum, C. pneumoniae, Mycoplasma spp., Ureaplasma spp., and B. bronchiseptica in the koala.
Materials and methods
Multiplex primers and dual-labeled fluorogenic probes
Nucleotide sequences for the multiplex rtPCR primers and dual fluorophore-labeled hydrolysis probes were designed (Beacon Designer v.8.2, PREMIER Biosoft International, Palo Alto, CA) based on sequence data available from GenBank (https://www.ncbi.nlm.nih.gov/genbank/; Table 1). Commercial sources provided the primers (GeneWorks, Adelaide, SA, Australia) and the dual-labeled probes (Biosearch Technologies, Novato, CA) used in our study. The dual-labeled probes were prepared by labeling reporter dyes to the 5’-terminus and black hole quencher dyes to the 3’-terminus of synthesized oligonucleotides. Primer sequences included in each multiplex were compared using the basic local alignment search tool, BLASTn (https://www.ncbi.nlm.nih.gov/BLAST/).
Oligonucleotide sequence of primers and fluorogenic probes for 2 multiplex real-time PCR panels.
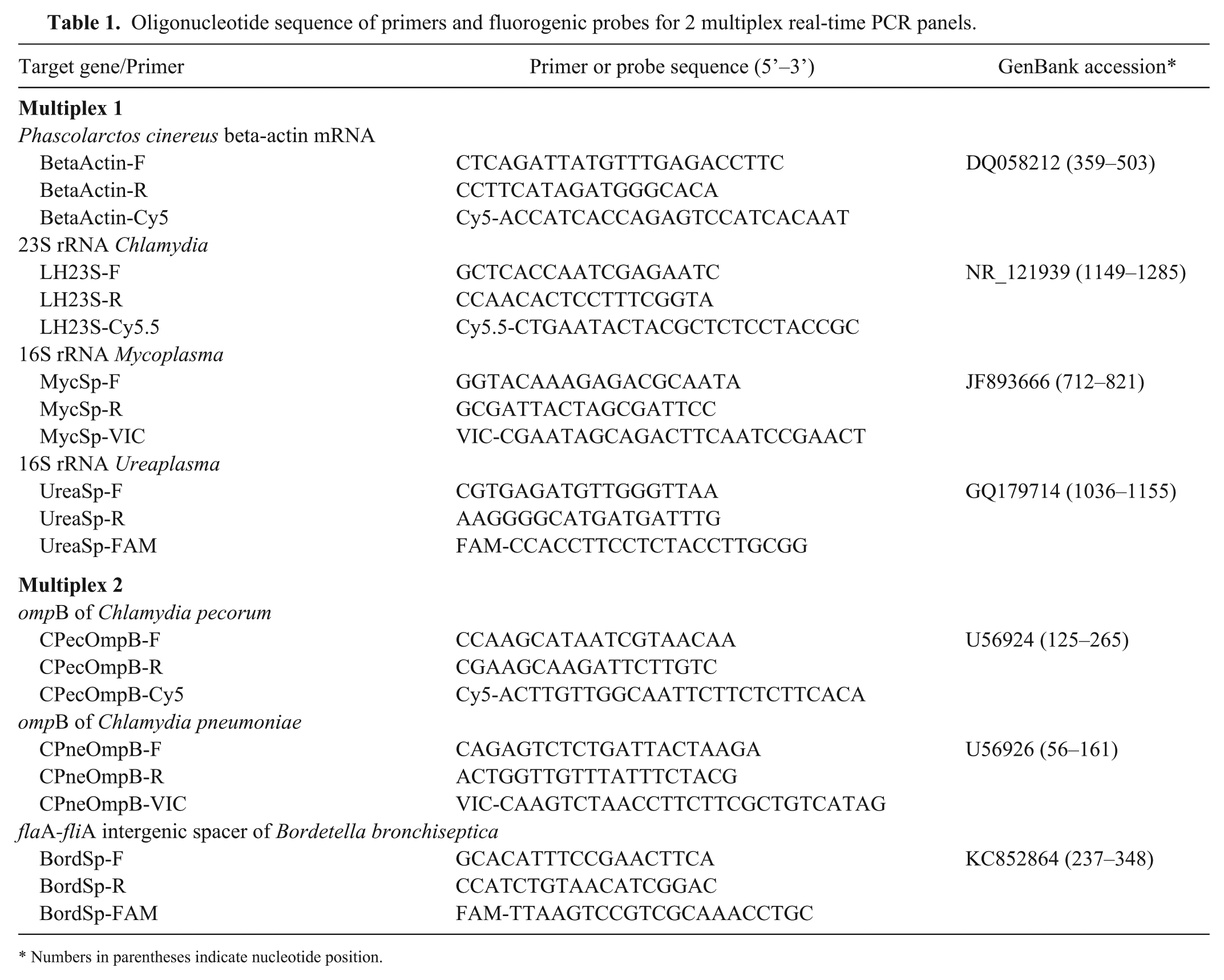
Numbers in parentheses indicate nucleotide position.
Multiplex rtPCR
Two multiplex rtPCR panels were designed. Individual samples were assayed with both multiplex panels. Multiplex panel 1 included the reference gene koala beta-actin, in order to confirm the presence of host cells in the sample and as a control for DNA quality. As well, multiplex 1 included universal primers targeting conserved regions of the relevant genome for the detection of isolates of Chlamydia, Mycoplasma, and Ureaplasma. The multiplex 2 PCR panel was designed to be run consecutively after multiplex 1 and, therefore, repeat screening of the endogenous control gene was not required. Multiplex 2 serves to validate Chlamydia results from multiplex 1, by identifying koala-specific Chlamydia species (including detection of all genotypes) of C. pecorum and C. pneumoniae, and also to detect B. bronchiseptica. Inclusion of specific pathogens in each multiplex panel was decided based on the priorities of a University of Queensland research project (KRG005—The pathology, incidence, treatment and management of chlamydiosis in the male koala).
Analytical specificity of reference strains
A total of 22 bacterial strains or isolates were used for evaluation of specificity of multiplex PCR panels (Table 2). We also included 5 bacterial strains of non-target species identified as either natural flora within the conjunctival, urinary, and reproductive tract of koalas (Higgins DP. Chlamydial disease in koalas. In: Diagnostic pathology of the diseases of aquatic, aerial and terrestrial wildlife. Wildlife Pathology Short Course, Sydney, 2012:222–230) or close relatives of B. bronchiseptica.
Specificity of the multiplex real-time PCR panels against target and non-target reference strains and isolates.
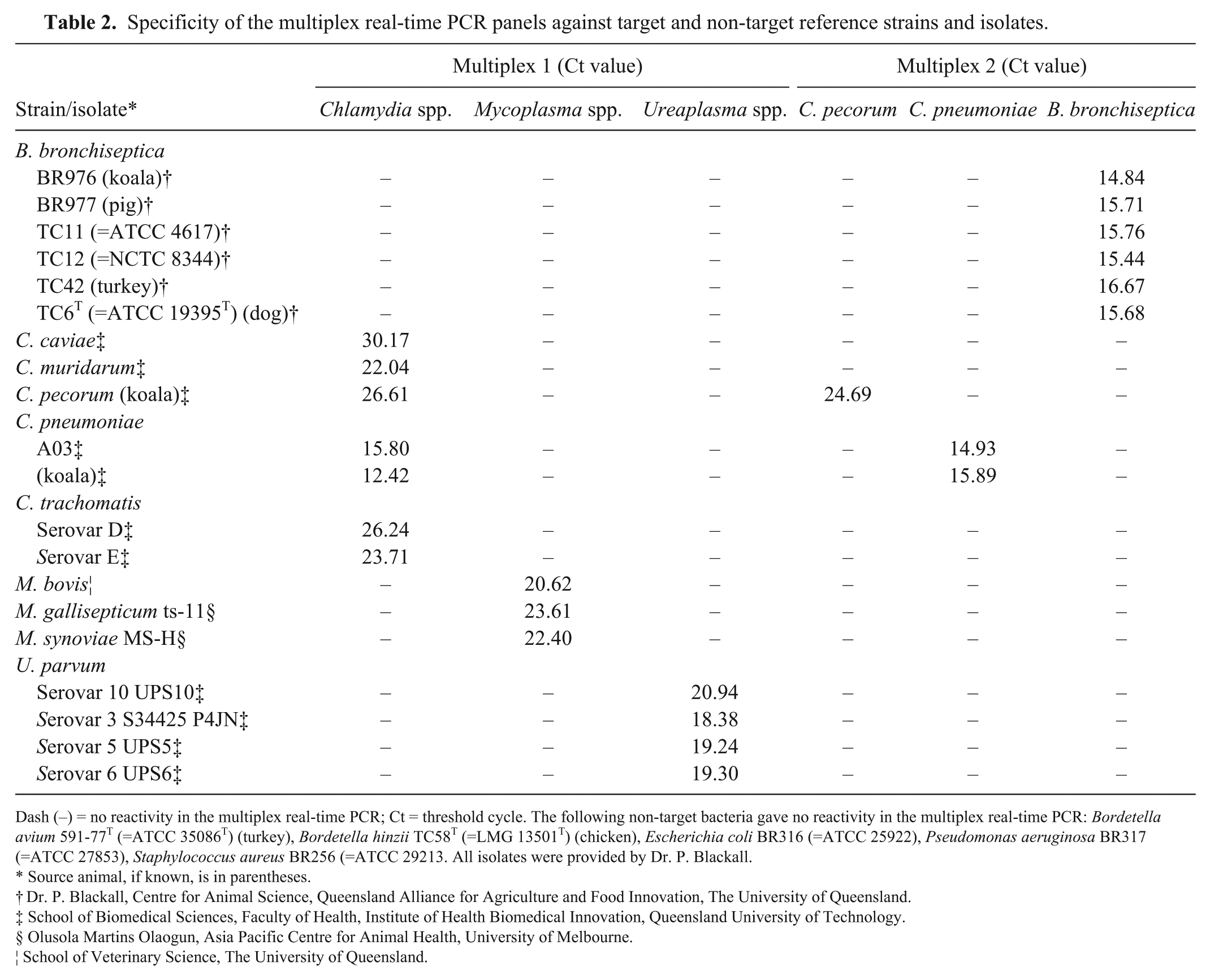
Dash (–) = no reactivity in the multiplex real-time PCR; Ct = threshold cycle. The following non-target bacteria gave no reactivity in the multiplex real-time PCR: Bordetella avium 591-77T (=ATCC 35086T) (turkey), Bordetella hinzii TC58T (=LMG 13501T) (chicken), Escherichia coli BR316 (=ATCC 25922), Pseudomonas aeruginosa BR317 (=ATCC 27853), Staphylococcus aureus BR256 (=ATCC 29213. All isolates were provided by Dr. P. Blackall.
Source animal, if known, is in parentheses.
Dr. P. Blackall, Centre for Animal Science, Queensland Alliance for Agriculture and Food Innovation, The University of Queensland.
School of Biomedical Sciences, Faculty of Health, Institute of Health Biomedical Innovation, Queensland University of Technology.
Olusola Martins Olaogun, Asia Pacific Centre for Animal Health, University of Melbourne.
School of Veterinary Science, The University of Queensland.
DNA amplification
Both multiplex rtPCR panels were carried out independently with a total reaction volume of 25 µL per multiplex assay, containing 10 µL of master mix (SensiFASTProbe No-ROX kit, Bioline, London, UK), 0.4 µM of each primer and 0.2 µM of probe, 8.2 µL of sterile distilled water in multiplex 1 and 8.65 µL in multiplex 2, and 5 µL of DNA template. Cycling parameters for the multiplex rtPCR panels consisted of initial denaturation at 95°C for 5 min, 38 cycles at 95°C for 5 s, and 10 s at either 57°C (for multiplex 1) or 58°C (for multiplex 2; Rotor-Gene Q, Qiagen, Doncaster, VIC, Australia). Fluorescence data were acquired during the annealing/extension phase. DNA extracted from the reference strains or isolates were used as the relevant positive control. A no-template control sample containing water instead of DNA template was included in every reaction run.
A standard curve for the C. pecorum PCR assay in multiplex 2 was established with a view to quantifying bacterial load in a sample. The C. pecorum quantitative PCR (qPCR) assay was calibrated using a known standard of quantified cell culture–derived, semi-purified koala C. pecorum genomic DNA diluted to 107–101, as determined by spectrophotometry (NanoDrop 1000 spectrophotometer, Thermo Fisher Scientific, VIC, Australia), and run in triplicate to determine the detection limit of chlamydial copy number.
Quantification of C. pecorum in clinical samples
To illustrate the versatility of this assay for use as a quantitative reporting platform, we further utilized the C. pecorum assay in conjunction with a standard curve to determine bacterial load for use in various clinical samples. By isolating DNA from quantified reference C. pecorum strain culture concentrations in infectious-forming units per mL (IFU/mL), we constructed a standard curve range of 74–740,000,000 IFU/mL (Supplementary Fig. 1). Using this standard curve reference, the level of detectable DNA concentration in various types of koala clinical samples was determined.
Validation of multiplex rtPCR panels
Validation of both multiplex PCR panels using reference strains and isolates to establish accuracy, followed published guidelines, 27 whereby specificity of all oligonucleotide sequences were verified using BLASTn, and each individual target was assayed in triplicate as a singleplex (SensiFAST SYBR No-ROX, SensiFAST Probe No-ROX, Bioline), using the reference strain for each assay to establish sensitivity. Melt-curve analysis of the SYBR assay was used for specificity. Specific reactions should result in a single melting peak corresponding to the PCR product being amplified. In contrast, multiple melting peaks imply that the reaction is either nonspecific, in that it originates from more than one amplicon, or that primer–dimers are being formed. Amplicon length was analyzed by gel electrophoresis and compared to the expected size of each target. Sequencing of PCR product was performed at the Animal Genetics Laboratory, School of Veterinary Science, University of Queensland (Gatton, QLD, Australia; Hitachi 3130x genetic analyser, Applied Biosystems, Scoresby, VIC, Australia). Alignment of sequencing data against GenBank target sequences was compared using BLASTn.
Application of multiplex rtPCR on wild and non-captive koala clinical samples
Clinical samples were collected as part of 2 University of Queensland research projects. For validation of the multiplex rtPCR assays in clinical samples, conjunctival, urogenital, and nasal swabs, in addition to male reproductive tract tissue and semen collected via electroejaculation, 16 were collected from koalas admitted to Moggill Koala Hospital, Currumbin Wildlife Hospital, and Australia Zoo Wildlife Hospital, and from male and female koalas of wild populations located at St. Bees Island (N −20° 55’ 0.012”, E 149° 25’ 59.988”) and Mount Byron (N −27° 6’ 55.008”, E 152° 40’ 23.016”). Opportunistic sampling of koalas admitted to wildlife hospitals included animals exhibiting clinical signs of disease and also signs unrelated to infectious disease (e.g. trauma, ill thrift, poor body condition). All samples were stored at −20°C until DNA was extracted (MagJET genomic DNA kit, Thermo Fisher Scientific) and purified (KingFisher Flex, Thermo Fisher Scientific).
Results
Verification of design of oligonucleotides
Alignment between primer sequences within each multiplex panel showed no cross-reactivity among target sequences. All of the sequence similarity values (Supplementary Table 1) were in the acceptable range (percent alignment >85 and expectation value <5). 24 Direct sequencing of amplification product revealed alignment to target the GenBank accession for each reference strain.
Comparative analytical sensitivity of multiplex real-time assays
The multiplex PCR system showed threshold cycle (Ct) results equivalent to the singleplex, confirming the multiplex PCR assay to be as sensitive as the singleplex format (Supplementary Table 2). The SYBR Green I assay yielded no amplification product for 3 of 6 reference target strains and for the host DNA. Further, each reference strain in both multiplex rtPCR panels showed no cross-reactivity with other primer/probe sequences (data not shown) or loss of amplification yield for individual assays when compared to singleplex rtPCR assays. The SYBR Green I melt curves and quantification data for each rtPCR assay are presented in Supplementary Figures 1–7.
Comparative analytical specificity
The isolates and strains reacted only in the relevant assay (Table 2). Both multiplex rtPCR panels showed 100% specificity.
Multiplex PCR detection of Chlamydia, Mycoplasma, Ureaplasma, and Bordetella in clinical ocular, reproductive, and respiratory samples
DNA was extracted from 228 conjunctival, 275 urogenital, and 38 nasal swabs, and the extracts were examined by both multiplex rtPCR panels (Table 3). Although C. pecorum was the predominant pathogen detected in both conjunctival and urogenital swabs, there was a low prevalence of Mycoplasma detected; Ureaplasma was not detected from either swab site. Bordetella was detected in nasal swabs.
Multiplex real-time PCR results from conjunctival, urogenital, and nasal swabs collected from hospitalized and wild male and female koalas.
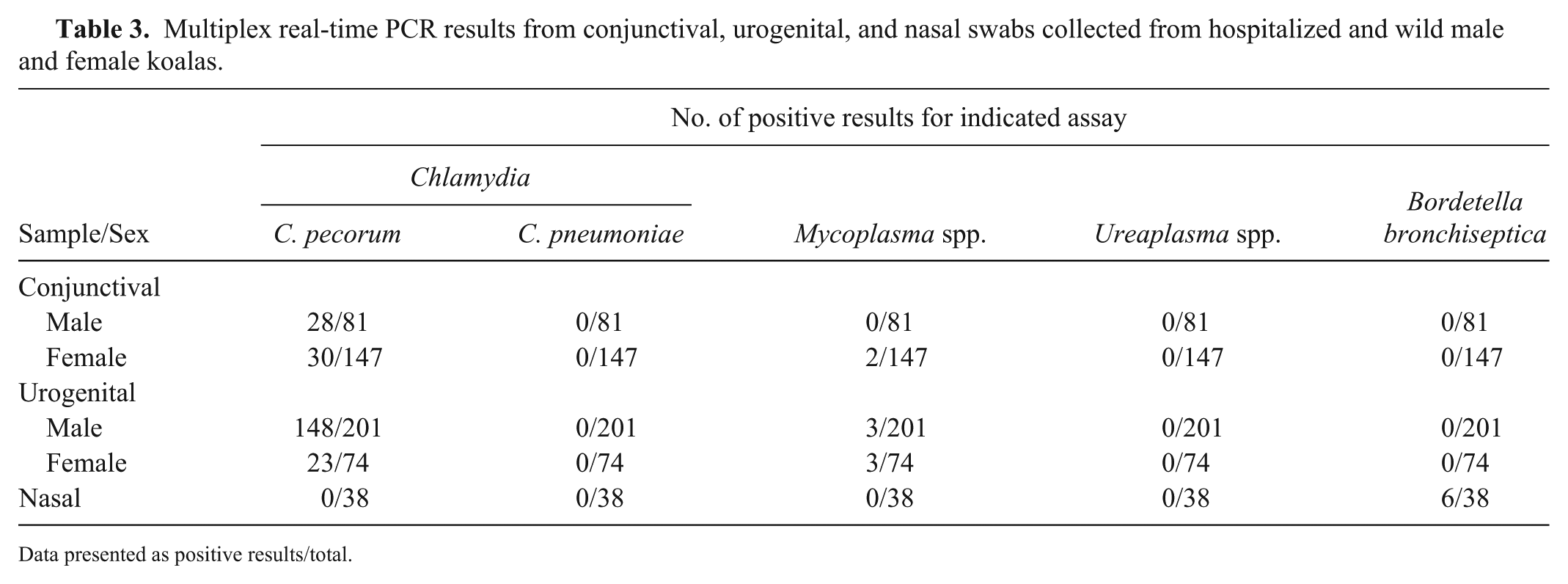
Data presented as positive results/total.
Chlamydial DNA was detected in both urogenital and reproductive tissues (Supplementary Table 3) and semen samples (Table 4). The predominant pathogen detected in urogenital and reproductive tissues and semen was C. pecorum. However, Mycoplasma was more common in male urogenital tract tissues and semen than in the penile swabs, suggesting that this organism may be a commensal. Various chlamydial loads were detected from 5 individual koalas across 3 clinical samples (urogenital swabs, semen, and prostatic tissue; Table 5).
Multiplex real-time PCR results from wild hospitalized male koalas.

Data presented as positive results/total.
Detection and quantification of Chlamydia pecorum in 3 types of male koala clinical samples from 5 animals.

IFU = infectious-forming units.
Discussion
We demonstrated that our B. bronchiseptica primer gene target exhibits specificity to the pathogen from various isolate sources, but we are mindful of cross-reactivity with B. parapertussis and the possibility of false-positive results based on the significant similarity of nucleotide sequence between the 2 strains. 32 To date, there have been no reports in the literature of B. parapertussis detection in koalas, to our knowledge, even though this pathogen is documented in other animal species. 33
We further demonstrated the versatility of this assay to quantify C. pecorum bacterial load through the use of a standard curve prepared with a reference strain of known concentration. In addition, we demonstrated that bacterial copy number can vary across animals and samples, which may reflect differences in infection localization and clinical outcomes. In addition to C. pecorum, this multiplex assay permits future quantitative analysis of the microbial species, and could be used for epidemiologic investigation of the impact of diseases in individual koalas or in populations.
The sensitivity of the multiplex rtPCR was comparable to each assay in the singleplex when comparing Ct values between assays (r² = 0.9). In contrast, assays performed with SYBR Green I were negatively correlated with the same values from multiplex assays, indicating reduced sensitivity and specificity to detect the reference strains. One of the major drawbacks of using SYBR Green I in rtPCR assays is the lack of specificity, 26 although the SYBR Green rtPCR method is relatively cost effective compared to the fluorescently labeled probe application.
Results from wild and hospitalized koalas revealed a high percentage of male koalas submitted to koala hospitals in southeast Queensland with Chlamydia PCR-positive penile swabs in the wild populations (74%), reproductive tract (67%), and semen (58%). These data suggest that Chlamydia infection in male koalas may be more prevalent than previously thought and that further studies investigating the significance of this infection to male infertility are necessary.
Testing of both hospitalized and wild koala populations revealed a relatively low prevalence of both Mycoplasma and Ureaplasma in the koala urinary and reproductive tract, semen, and conjunctiva, suggesting that they are unlikely causes of any major disease significance at a population level. Nevertheless, further research to determine whether Mycoplasma or Ureaplasma play any role in disease expression, 11 especially in regard to koala infertility (prostatitis or vaginitis), is warranted. Given that Mycoplasma was detected in the tissues of the male reproductive tract and semen (~5% of all ejaculates tested), it is possible that Mycoplasma may not only be associated with koala prostatitis but also have an effect on spermatogenesis (orchitis) or sperm maturation and storage (epididymitis).
Supplemental Material
SF1_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF1_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
SF2_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF2_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
SF3_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF3_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
SF4_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF4_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
SF5_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF5_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
SF6_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF6_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
SF7_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, SF7_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Supplemental Material
Tables_S1-S3_JVDI_10.1177_1040638718770490 – Supplemental material for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala
Supplemental material, Tables_S1-S3_JVDI_10.1177_1040638718770490 for Development and application of two multiplex real-time PCR assays for detection and speciation of bacterial pathogens in the koala by Lyndal S. Hulse, Danica Hickey, Jessica M. Mitchell, Kenneth W. Beagley, William Ellis, Stephen D. Johnston in Journal of Veterinary Diagnostic Investigation
Footnotes
Acknowledgements
We gratefully acknowledge Patrick Blackall from the University of Queensland for donating the majority of isolates for specificity testing and also providing advice regarding the manuscript. Additional strains or isolates were kindly donated by Olusola Martins Olaogun, Asia Pacific Centre for Animal Health, University of Melbourne; Emily Bryon, School of Biomedical Sciences, Faculty of Health, Institute of Health Biomedical Innovation, Queensland University of Technology, Kelvin Grove, QLD, Australia; and Tina Maguire, School of Veterinary Science, University of Queensland, Gatton, QLD, Australia. We also thank Chiara Palmieri, School of Veterinary Science, University of Queensland, Gatton, QLD, Australia.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This research was funded by the Department of Environment and Heritage Protection, Queensland Government; project no. KRG005.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.