
Abstract
Johne’s disease, caused by Mycobacterium avium subsp. paratuberculosis (MAP), is a chronic, progressive bacterial enteritis of ruminants that can cause serious losses in both livestock and exotic species. Infection risk in exotic ruminants is associated with maternal infection status, but the effect of other herdmates on risk of infection has not been reported, to our knowledge. We conducted a retrospective cohort study to evaluate the association between MAP infection status and early-life contact with infected herdmates. The study population included 3,234 individuals representing 128 species at San Diego Zoo Global facilities between 1991 and 2010. Animal movement, health, and pathology records were used to trace enclosure-sharing contacts between members of the study population and any MAP-infected animal. Contact-days were counted by age of the reference animal and the number of unique infected individuals contacted. Herdmate infection status was stratified by stage of infection (180 d prior to diagnosis), age, and whether relevant lesions were found at autopsy. Having an infected herdmate was a strong risk factor for infection (OR = 4.4; 95% CI: 1.9–10.3), and each method of defining herdmate infection status showed significant differences in infection risk. The best predictor was number of contact-days within the first week of life, with a 2-fold increase in risk associated with each doubling in the number of contact-days (OR = 2.1; 95% CI: 1.1–4.0). We conclude that early contact with infected animals is an important predictor of MAP infection risk, although the effect size is smaller than that previously described for maternal infection status.
Introduction
Johne’s disease, or paratuberculosis, is caused by infection with Mycobacterium avium subsp. paratuberculosis (MAP). It is a chronic inflammatory intestinal disease primarily affecting ruminants 8 although infection has been documented in a broad range of other species. 1 Disease has been recorded in numerous exotic ruminant species in zoo collections 8 and, as in livestock industries, heavy losses are incurred in the form of impaired reproduction, early culling, and concomitant loss in reproductive potential and genetic diversity, as well as increased costs as a result of veterinary intervention, surveillance, and control measures. 21
Specific information on transmission pathways in exotic ruminants is limited, but the major route of infection in livestock is direct ingestion of bacteria from the feces of infected animals. 12 Maternal infection status is a commonly described risk factor for domestic ruminants, and infected animals in a zoo setting have also been shown to be significantly more likely to have infected dams compared to their disease-free conspecifics, indicating a likely role of perinatal (such as ingesting contaminated manure while suckling), trans-mammary, or intrauterine transmission in exotic ruminant species. 21
Disease control in intensive livestock systems centers on early intervention and hand-rearing of young stock in disease-free conditions. 2 Early separation of offspring is often impractical in zoologic settings and may prove deleterious to conservation breeding programs. To develop a comprehensive Johne’s disease risk management framework, it is necessary to determine the primary modes of transmission and identify important risk factors in a zoo setting. Building on previous work 21 that identified maternal infection status as a risk factor for MAP infection, we assessed the association between early-life contact with infected animals and MAP infection status. To enable risk stratification, we have categorized potential exposures based on host and infected-contact features including age, species, stage of infection, and the presence of gross anatomic lesions.
Materials and methods
Study population
San Diego Zoo Global (SDZG), including the San Diego Zoo and San Diego Zoo Safari Park, is a multi-site zoo and conservation organization with extensive breeding programs located in southern California. A retrospective cohort study was conducted on ruminants housed at SDZG from 1 January 1991 through 31 December 2010, including some of the same individuals as the previous case-control study of Johne’s disease in this collection. 21 Data collection for our study ended in December 2010, as a result of changes in data collection and surveillance practices within the collection around this time. The study population is defined as the subgroup of ruminants that were born at SDZG during the study period and lived to be at least 7 d of age. Animals in this group were excluded from the study if they had unknown MAP infection status (as defined below), or unknown early life exposure status (because they were hospitalized or moved to assisted care within the first week of life).
Surveillance
The MAP infection surveillance protocol aimed to annually test all ruminants >10 mo old by fecal culture beginning in early 1992. Fecal samples were collected in triplicate from each animal and sent to the Johne’s Testing Center at University of Wisconsin (Madison, WI) for liquid radiometric culture (BACTEC 12B, BD, Sparks, MD) and identification of MAP isolates. 3 This test has not been validated in exotic ruminant species, but sensitivity of 54.4% and specificity of 100% are reported in cattle. 11 Since 1991, all animals from SDZG that died or were euthanized have received full postmortem examinations by a board-certified veterinary pathologist, which includes gross examination, histopathology on a prescribed set of tissues, and ancillary testing, if indicated, based on gross and histologic findings. Acid-fast stains of lymph nodes were examined, and cultures of intestinal content, mesenteric lymph node, and ileum were submitted for examination (to the same laboratory) if gross pathology or clinical signs were suggestive of Johne’s disease (thickened intestinal wall or enlarged mesenteric lymph nodes, unexplained weight loss, or chronic diarrhea).
Case definition
Surveillance culture and autopsy results were used to determine whether each individual developed MAP infection during the course of follow-up. Animals with any of the following features were classified as infected: 1) positive fecal culture surveillance sample, 2) presence of acid-fast bacilli of appearance consistent with MAP in the ileum or mesenteric lymph node, 3) MAP-positive tissue culture at autopsy. Animals with none of these features were classified as not infected only if they had been tested at >2-y-old or had been autopsied at >2-y-old, a time at which the probability of shedding increases in domestic species.9,18 Remaining animals were excluded from further analysis (as outlined under “Study population” above) because their infection status could not be determined reliably.
Contact tracing
For the purposes of our study, a “contact” with an infected animal is defined as any occasion when a member of the study population occupied the same enclosure as a MAP-infected animal for ≥1 d, if the infected animal was >6-mo-old at the time. Contacts of this kind were traced using enclosure movement records. The number of contact-days and the count of unique infected individuals contacted were recorded. Contact was assessed separately for 2 periods in the reference animal’s life: week 1 of life (1–7 d from birth), and the remainder of the first 6 mo of life (8–180 d from birth) to examine whether contact with infected herdmates (animals of the same species or subspecies housed together) within these different time periods was associated with differences in infection risk. One day in contact with a single infected animal is counted as one contact-day; therefore, study animals in contact with more than one MAP-infected animal can accrue more than one contact-day in a calendar day.
Defining contact within each herd
We defined a herd as a group of animals housed together of the same species or subspecies, depending on the level at which each herd is managed. Types of contact with infected herdmates were stratified by linking each contact event to the herdmate’s husbandry, life history, autopsy, and diagnostic data. This allowed the exposure to be further classified according to presence of diagnostic lesions, how long before diagnosis the contact occurred, age of the MAP-infected animal at the time of contact, and age of the reference animal at the time of contact. Contact with an infected animal was then used to calculate the following variables: 1) shedding risk: “high” for contact with an infected animal that ultimately had relevant lesions at autopsy, “low” if the infected animal had no relevant lesions at autopsy, versus “no contact” if the animal had no known contact with an infected herdmate during the targeted periods; 2) infection stage: “late-stage” if the contact event occurred within 180 d of the infected herdmate’s diagnosis (collection date of first positive fecal sample or autopsy date), “early-stage” if the contact event occurred prior to that time, compared with animals with no known contact with an infected herdmate; and 3) age of the MAP-infected animal when enclosure sharing occurred: no contact (reference), 6-mo to 2-y-old, 2–5-y-old, >5-y-old; and 4) age of the reference animal at the time of contact: first 7 d of life, 8–180 d. Variables describing risk and stage of infection of the infected animals were included in the models as categorical variables describing the highest hypothesized risk level of contact experienced by the reference animal at any time during the study period.
Defining contact with members of other species
Associations between contact with infected members of sympatric herds (other species with overlapping ranges in the same enclosures) and MAP infection were evaluated among a subset of the study population that was housed within 13 large, multi-species enclosures (0.4–36.4 ha) at the San Diego Zoo Safari Park. Accurate and complete descriptions of space use by study animals were not available, so occupancy ranges for each herd were delineated manually using survey maps and aerial photographs of enclosures in consultation with 3 experienced animal managers and keepers that worked in the identified enclosures during the study period. Identified ranges were translated into computerized maps using a geographic information system (ArcGIS 9.3, Environmental Sciences Research Institute, Redlands, CA), and spatial overlap in species’ ranges were identified by the intersections of the estimated range polygons.
An animal was deemed to be a member of a sympatric herd if it was of a different species whose approximated occupancy range overlapped with that of the reference animal’s range. Presence or absence of contact with an infected individual of a sympatric herd was determined in a manner similar to that described above: individuals were considered “in contact” if they shared an enclosure with an infected sympatric species during the identified periods and “not in contact” otherwise. Contact with other (non-sympatric) species through general enclosure sharing was also calculated, but too few animals had only this type of contact to be included in the analysis.
Statistical analyses
Screening of all potential predictors of MAP infection status was performed using univariate logistic regression models. All variables yielding p < 0.20 were considered for inclusion as main effects in multivariable analysis. Mixed-effects logistic regression models 6 were used in the multivariable analysis to estimate the association between the different identified contact types and probability of MAP infection in the study population. A random effect was included using “herd” (animals of the same species or subspecies housed together) as the grouping variable to account for differences in baseline disease risk by management unit caused by unmeasured factors. All other predictors were parameterized as fixed effects and added to the model in a forward stepwise fashion. Separate models were fitted using the different risk stratification methods (risk level based on lesions, stage of infection based on time to diagnosis, age of infected contact, same versus different species). Main effects were considered significant if the 2-tailed, Wald-based p value for that term was <0.05. Potential confounders were retained in the model if their inclusion altered the estimated value of another parameter by ≥10%. Potential confounding variables were not limited to those yielding p values <0.20 in the univariate analysis. Continuous variables (such as a count of the days in contact) were included in either raw form or log-transformed (base 2), according to which model fit the data best. The best-fit multivariate model among competing risk stratification methods was chosen based on the lowest Akaike information criterion (AIC) statistic. A term was included in the final model to control for birth year. This variable was included to account for the fact that declining prevalence over time likely leads to a declining level of environmental contamination. Inclusion of this variable is needed to estimate the effect of contact on infection risk, independent of the simultaneous effect of infection prevalence via environmental contamination. All statistical analyses were performed using R 2.15 (https://www.R-project.org); multivariable models were fitted using the function “glmer” in the package “lme4” (https://goo.gl/CYKzqJ).
Results
Over the course of the study period (1 January 1991 through 31 December 2010), 151 animals were diagnosed with MAP infection (and used in contact-tracing between study animals and any infected animal). Test-positive animals were aged 4 d to 13 y (median: 3.2 y; interquartile range: 1.7–6.9) at diagnosis and all were born between 1976 and 2006. Seventy-six of 151 test-positive animals (50.3%) had lesions characteristic of Johne’s disease at autopsy. Sixty of these 151 animals also met the criteria for inclusion in the study population. The study population included a total of 3,234 animals that met the specified criteria, giving an overall prevalence of 1.9% (60 of 3,234).
Univariate analysis (Table 1) showed that higher MAP infection prevalence was associated with male sex (p = 0.044), earlier birth year (p < 0.001), and positive maternal infection status (p < 0.001). The infection odds for animals sharing an enclosure with an infected herdmate at any time during the first 6 mo of life were more than 9 times those of animals with no infected animal contact (OR = 9.1; p < 0.001). All methods of risk stratification also showed significant differences in infection odds among groups (p < 0.001), with the strongest effects being for late-stage infection (last 180 d prior to diagnosis; OR = 19.3) and risk level based on characteristic lesions at autopsy (OR = 15.2).
Univariable analyses. Results of 9 separate logistic regression models examining the association between the listed exposures and MAP infection status among 3,234 ruminants housed at San Diego Zoo Global facilities (1991–2010).
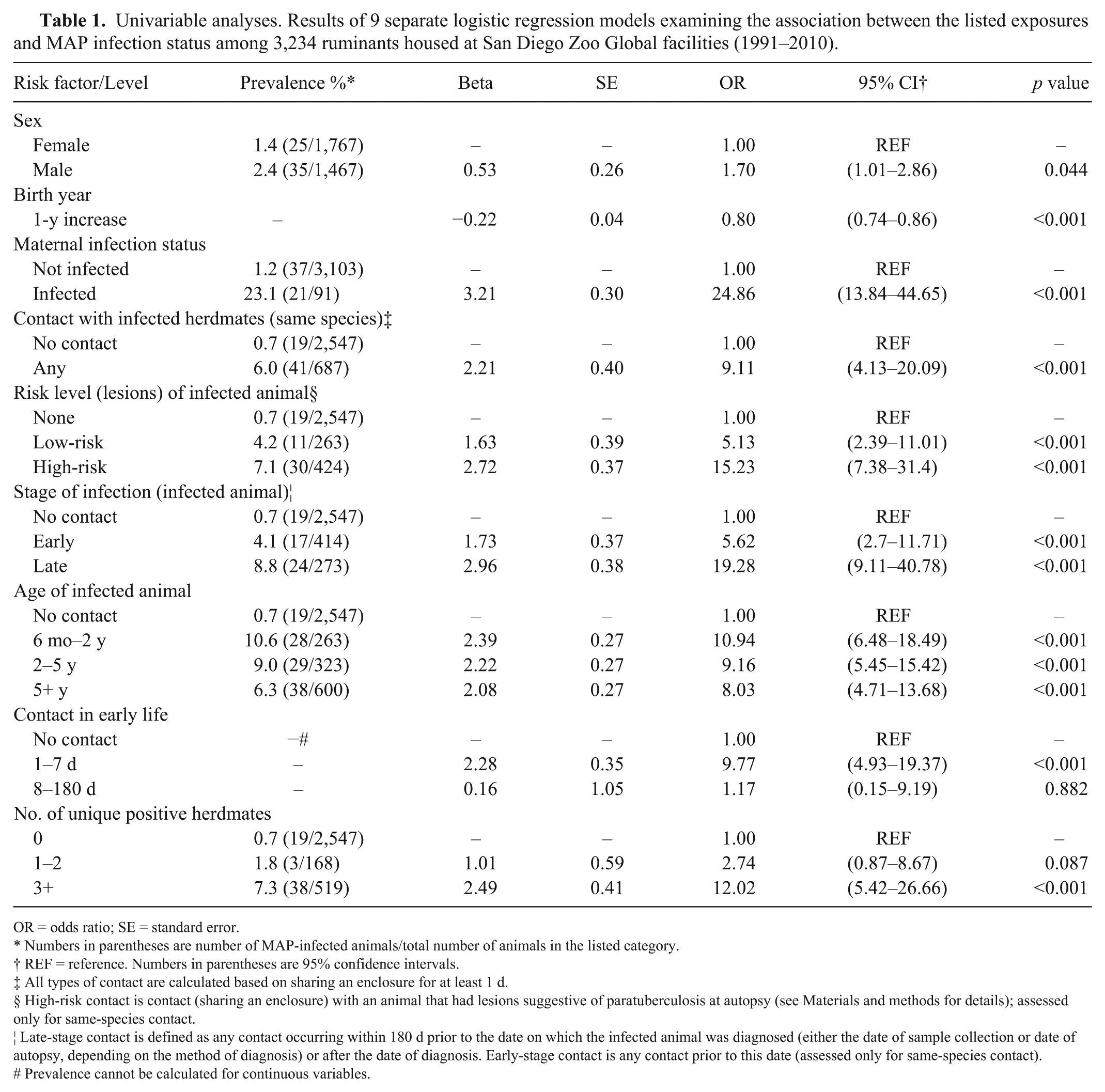
OR = odds ratio; SE = standard error.
Numbers in parentheses are number of MAP-infected animals/total number of animals in the listed category.
REF = reference. Numbers in parentheses are 95% confidence intervals.
All types of contact are calculated based on sharing an enclosure for at least 1 d.
§ High-risk contact is contact (sharing an enclosure) with an animal that had lesions suggestive of paratuberculosis at autopsy (see Materials and methods for details); assessed only for same-species contact.
¦ Late-stage contact is defined as any contact occurring within 180 d prior to the date on which the infected animal was diagnosed (either the date of sample collection or date of autopsy, depending on the method of diagnosis) or after the date of diagnosis. Early-stage contact is any contact prior to this date (assessed only for same-species contact).
# Prevalence cannot be calculated for continuous variables.
Separate mixed-effects logistic regression models were fitted for the different methods of classifying contact, each including variables denoting maternal infection status, birth year, and sex, which were included as potential confounders in all multivariate models (Table 2). Each method of subclassifying contact with infected animals yielded a significant predictor of MAP infection status at the highest level of exposure when compared to the reference group (no contact). The best-fit model based on lowest AIC (Table 3) was that including the number of contact-days with infected herdmates accrued during early life (first 7 d). A 2-fold increase in the number of infected herdmate contact-days (range: 0–70) was associated with an increase in infection odds (OR = 2.1; 95% CI: 1.1–4.0), whereas increase in the number of contact-days accrued after the first week of life (8–180 d after birth) did not lead to a significant increase in infection odds (Table 4). Separate, adjusted models estimated ORs of 4.4 (95% CI: 1.9–10.3) for contact with any infected animal, 5.4 (95% CI: 2.4–12.1) for contact with infected animals having relevant lesions at autopsy, 4.3 (95% CI: 2.1–9.0) for contact with ≥3 individual infected animals, and 4.5 (95% CI: 2.0–10.0) for contact with infected animals within 6 mo before diagnosis. The age of the infected animal was also a significant predictor of infection status; contact with both younger (6 mo–2 y) and older (≥5 y) animals was associated with a significant increase in infection odds, but this was not the case for animals of intermediate age (2–5 y).
Results of 6 separate, mixed-effects logistic regression models examining the association between listed exposures and MAP infection status among 3,234 ruminants housed at San Diego Zoo Global facilities (1991–2010). All models contain a random effect of species or subspecies according to how each herd is managed in the collection (n = 128), and control for sex, birth year, and dam infection status.
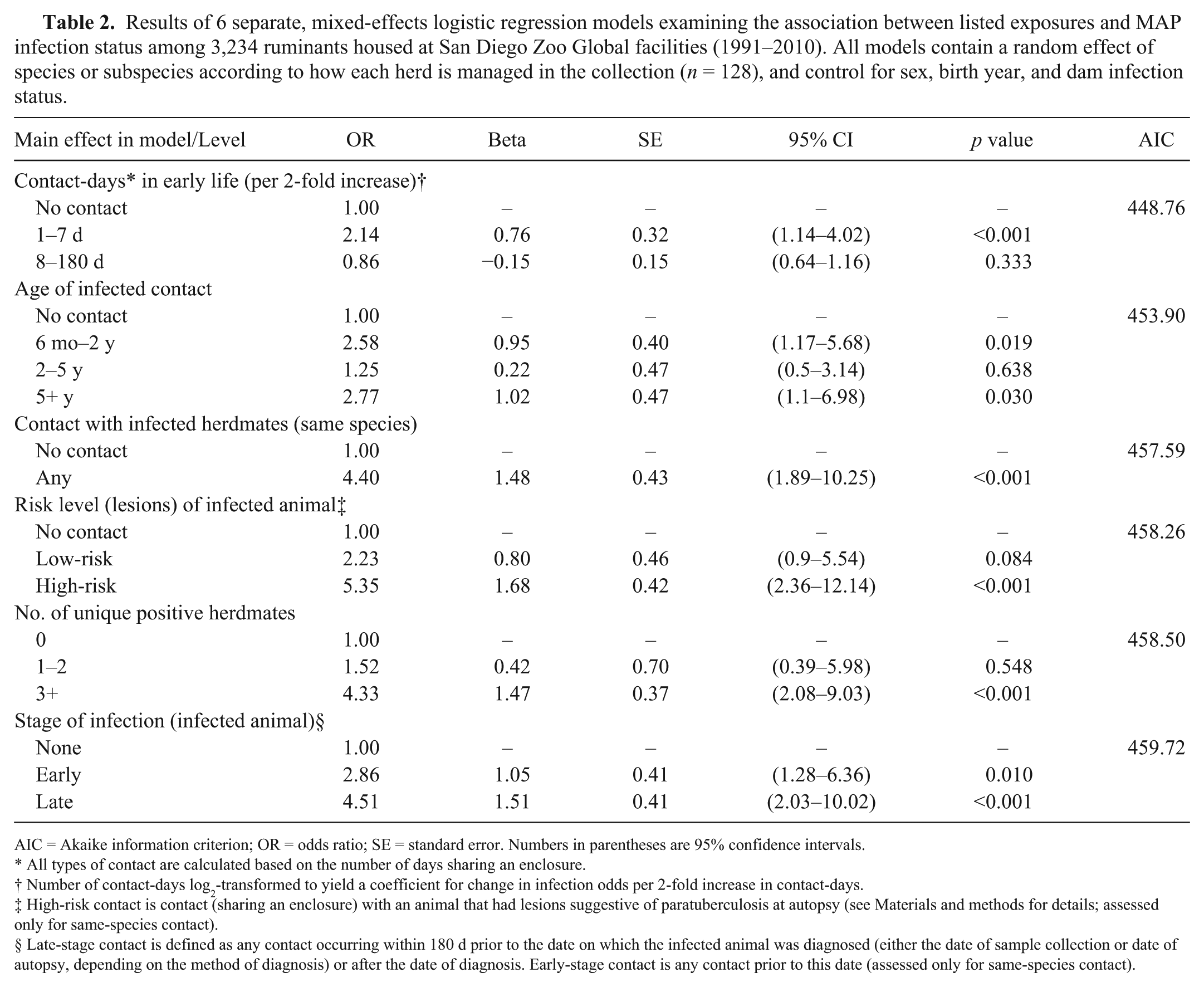
AIC = Akaike information criterion; OR = odds ratio; SE = standard error. Numbers in parentheses are 95% confidence intervals.
All types of contact are calculated based on the number of days sharing an enclosure.
Number of contact-days log2-transformed to yield a coefficient for change in infection odds per 2-fold increase in contact-days.
High-risk contact is contact (sharing an enclosure) with an animal that had lesions suggestive of paratuberculosis at autopsy (see Materials and methods for details; assessed only for same-species contact).
§ Late-stage contact is defined as any contact occurring within 180 d prior to the date on which the infected animal was diagnosed (either the date of sample collection or date of autopsy, depending on the method of diagnosis) or after the date of diagnosis. Early-stage contact is any contact prior to this date (assessed only for same-species contact).
Best-fit (based on the Akaike information criterion) mixed-effects logistic regression model examining the association between listed exposures and MAP infection status among 3,234 ruminants housed at San Diego Zoo Global facilities. Model contains a random effect of species or subspecies according to how each herd is managed in the collection (n = 128).
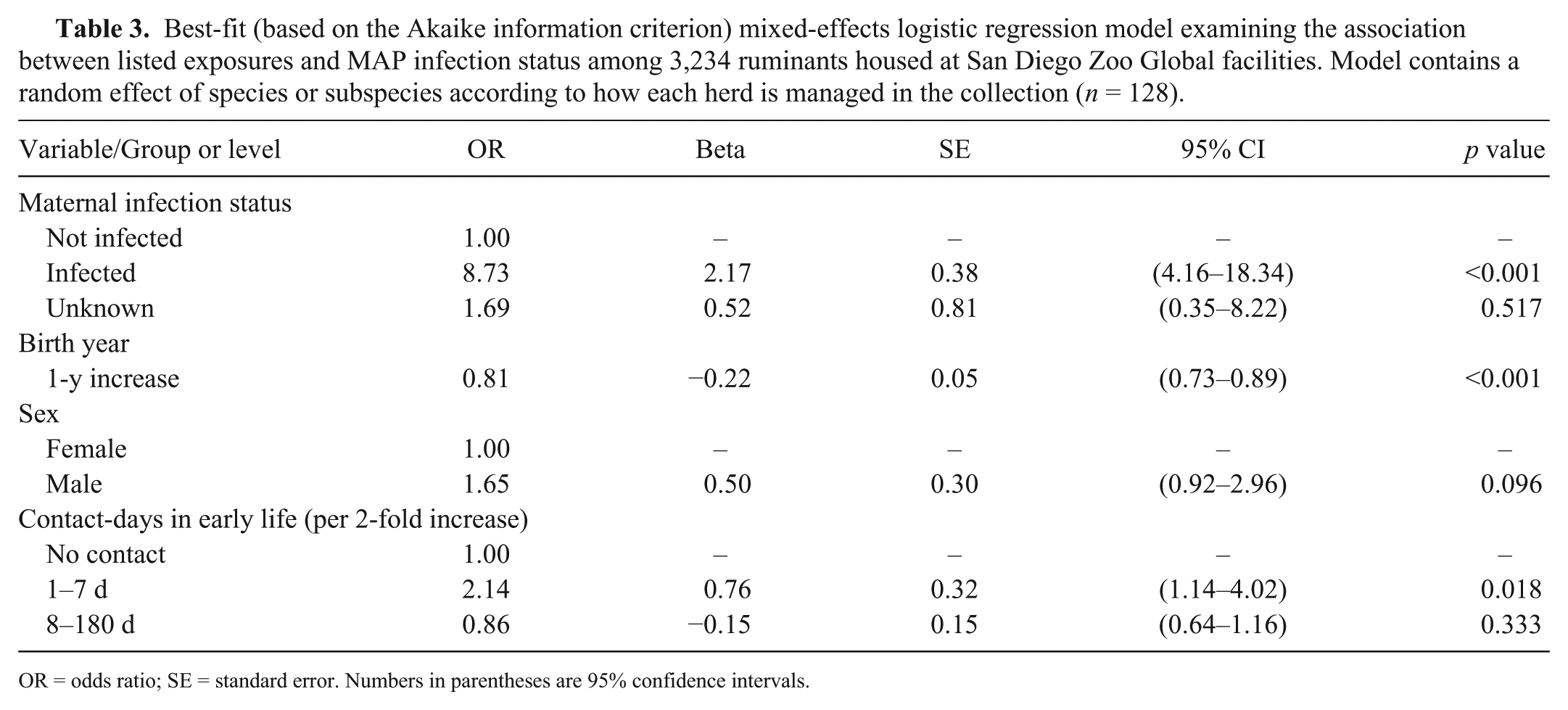
OR = odds ratio; SE = standard error. Numbers in parentheses are 95% confidence intervals.
Best-fit (based on the Akaike information criterion) mixed-effects logistic regression model examining the association between listed exposures and MAP infection status among 1,517 ruminants housed in large multispecies enclosures at San Diego Zoo Safari Park. Model contains a random effect of species or subspecies according to how each herd is managed in the collection (n = 64).
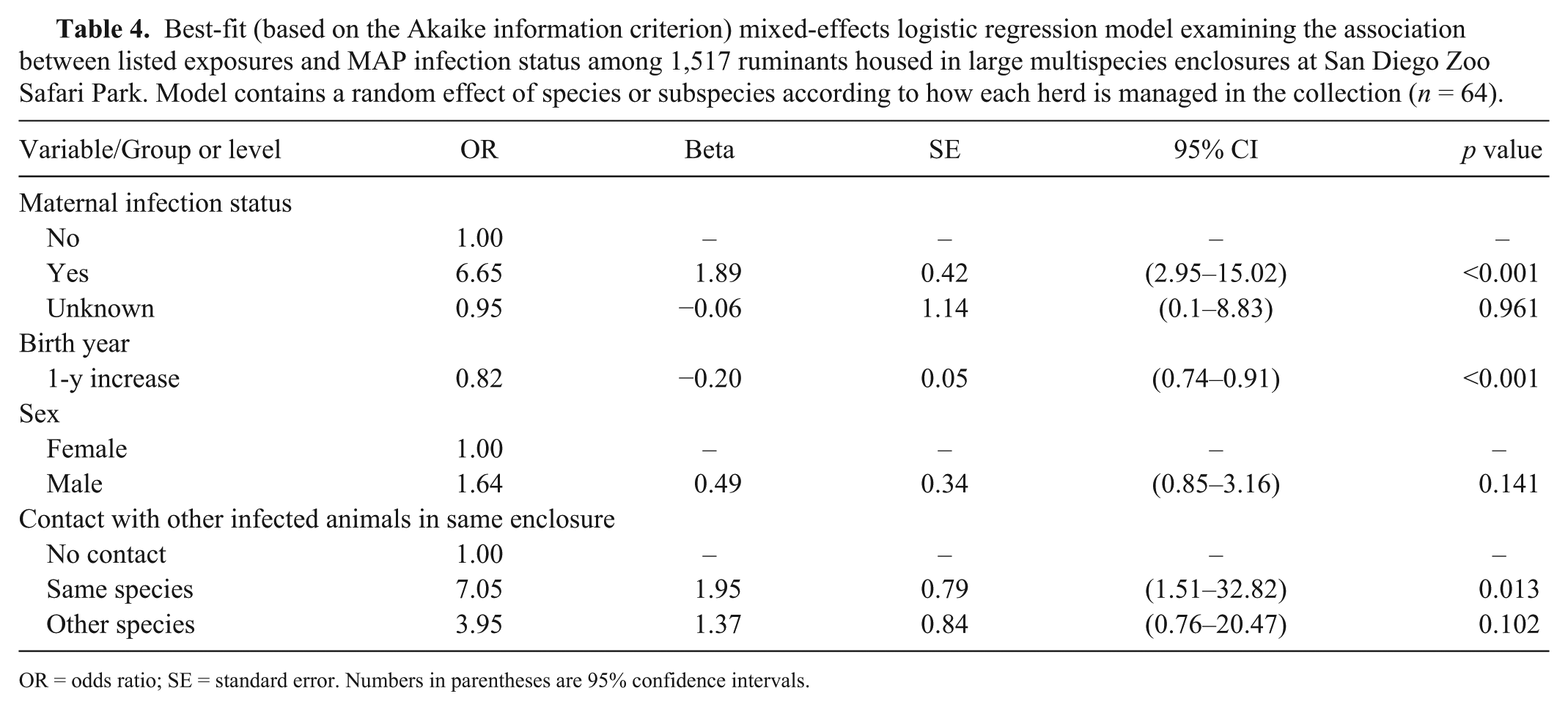
OR = odds ratio; SE = standard error. Numbers in parentheses are 95% confidence intervals.
The subgroup analysis of animals living in the large multispecies enclosures for ≥1 d found significant differences in infection odds for contact with different species of infected animal. In this model, contact with infected herdmates at any time during the first 6 mo of life was associated with a prevalence OR of 7.1 (p = 0.013). Sharing an enclosure with an infected animal of a sympatric species was not statistically significant (OR = 4.0; p = 0.102).
Discussion
We assessed the association between MAP infection in zoo ruminants and characteristics of contact with other infected animals through sharing enclosures. A strong relationship was identified between infection status and different types of contact, even after adjustment for other risk factors. The model based on the number of contact-days with other infected animals during the first week of life provided the best fit to the data. Although the effect size is not as great as that associated with maternal infection status, 21 this may be a very important period for infection risk. In our dataset, 21 of 60 infected animals in the study population had infected dams, and of the remaining 39, 30 had contact with an infected herdmate (same species or subspecies) within the first week of life.
The predictor of infection status that best fit the data was the number of contact-days with MAP-infected herdmates during the first week of life. This variable remained a significant predictor of infection status, even after controlling for sex, maternal infection status, and birth year (as a proxy for the declining level of environmental contamination, because overall prevalence has decreased over time). 21 This finding suggests that the first week of life is a critical period in MAP infection risk for exotic hoofstock, independent of maternal infection status. Changes in susceptibility to infection with age are incompletely understood, even in livestock, but the perinatal period is believed to be the predominant time of transmission of MAP infection in dairy cattle, with fecal ingestion in contaminated birthing pens being specifically implicated. 18 A 2010 review of experimental infection studies in cattle concluded that both infection risk and disease risk decline with increasing age at time of exposure, but cattle of all ages may be infected by high doses of MAP. 20 Infection trials in red deer (Cervus elaphus), on the other hand, showed a strong relationship only with clinical disease, but not infection, 7 and no age-related change in susceptibility has been documented in sheep. Furthermore, the conditions in which zoo ruminants live and give birth differ significantly from those of dairy cattle, with births typically occurring in larger enclosures with less risk of direct fecal contamination and ingestion. Nonetheless, the strong effect of maternal infection status, both in the current study (OR = 8.7) and in previous work (OR = 6.8), 21 is suggestive of an important role for perinatal (fecal–oral, intrauterine, or trans-mammary) transmission in exotic ruminant species. The results of our study suggest that other routes of transmission between members of a same-species herd living in close association are also important. Fecal–oral transmission in this scenario is plausible given that studies of shedding levels in infected animals suggest that only a very small quantity of fecal contamination is required in susceptible young stock.5,10,17,18
Number of unique infected animals contacted, the time-to-diagnosis (stage), and presence of lesions, when analyzed separately, were strongly associated with MAP infection, and the effect sizes were similar. These 3 variables were strongly correlated within this dataset, given that older animals were often closer to their date of diagnosis and frequently had relevant lesions at autopsy, and animals with greater numbers of infected contacts were also more likely to have accrued contact with late-stage or high-risk animals at some time. It is thus difficult to determine which of these is most important in predicting disease risk. Our results do, however, indicate that the number of infected contacts or the time elapsed between contact and diagnosis may be similarly useful as predictors of infection risk as the presence of lesions in the infected contact. The former two are likely to be more cost-effective given that they can be determined solely from management records, rather than requiring an extensive postmortem examination, which may not be sensitive in detecting early lesions. In our dataset, only half of infected (culture-positive) animals (76 of 151) had relevant lesions at autopsy. This is consistent with the pattern observed in sheep and cattle, where histopathology and culture results are often inconsistent, many animals do not show overt gross or histologic changes, and subclinically infected animals are capable of maintaining infection in a herd. 18 In sheep and goats, radiometric liquid culture has even been reported to be more sensitive than intestinal histopathology.16,19 Although fecal culture is traditionally regarded as having 100% specificity,11,15 it is worthy of note that “pass-through” positive fecal culture results for MAP have been documented in uninfected cattle ingesting heavily contaminated feed or water, 13 so the apparent high sensitivity compared with histopathology may in fact include some false-positive results.
Age of the infected animal at the time of contact was not useful to separate levels of infection risk. We hypothesized that older infected animals would be more likely to shed large numbers of bacteria, thus representing a greater infection hazard. The univariate analysis yielded similar ORs regardless of the age of the oldest infected contact. The multivariable model including the number of infected animal contact-days also did not suggest a significant increase in infection risk with age of the infected animal. Contact with either older (≥5 y) or younger (6 mo–2 y) infected animals was associated with greater infection risk than those with no infected animal contact, whereas, the OR for contact with infected animals of intermediate age (2– 5 y) was not significantly different from one. This result should be interpreted with caution, as the group of animals with only this type of contact is small, and most animals had contact with all age groups. Overall, the findings suggest that the age of the infected animal at the time of contact was not predictive of infection risk in this population.
Targeted surveillance or proportional risk sampling can yield increased effectiveness for disease detection, and typically strata with increased risk of disease occurrence or increased risk of disease dissemination are targeted for sampling. 14 A sampling plan that targets animals with infected dams and those that shared an enclosure with an infected herdmate during the first week of life would include 85% of known cases (51 of 60) within the study population. Of the remaining 9 cases, just 1 had a recorded contact with an infected animal (a sympatric species); the other 8 had no recorded contact with any infected animal. These 8 animals may have been infected through contact with undiagnosed cases or general enclosure contamination. We therefore conclude that little further advantage was likely to be obtained from contact-tracing within this population beyond targeting of individuals with infected dams and first-week-of-life contact with infected herdmates. Furthermore, even these methods will not identify 100% of infected animals unless more sensitive testing is available. It is also worthy of note that 98 infected animals were excluded from the study population because of uncertain early life history—principally animals born elsewhere and imported into the collection. Imported animals must also be assumed to represent a higher risk of infection to their herdmates compared with those born onsite but with no detected maternal or early-life herdmate contacts.
No significant sex predilection was identified by our results, but an association cannot be ruled out—increased prevalence and shedding intensity of MAP in male sheep has been reported in the past. 4 In neither study can a biological relationship be established given potential differences in important management procedures by sex, but both results are compatible with the possibility of a behaviorally or physiologically mediated sex predilection to MAP infection. Disease odds declined ~20% with every 1-y increase in birth year. A decline in the frequency with which cases are diagnosed has been anecdotally observed in this collection, and an ~8-fold decline in cumulative lifetime incidence of MAP infection was reported 21 in the same collection between 1991 and 2007. This effect can be attributed to the systematic Johne’s disease surveillance and management program implemented in 1992.
We hypothesized that contact with infected animals of another species may be equally important as contact with herdmates of the same species group, especially when they were known to regularly overlap in their ranges. The effect of sharing an enclosure with infected animals of different species was associated with slightly higher infection odds, but this was not a statistically significant increase in infection risk. Such a relationship cannot however be ruled out—the home range data were qualitatively assessed and cross-sectional in nature. These data are also subject to recall or individual-observer biases and may not be representative of the whole study period or the time period relevant to transmission. It is possible that such a relationship would be detected using more spatiotemporally precise data on overlapping space use by study species.
Results from our study are derived from observations of one of the largest, multispecies, hoofed mammal collections over an extended time period and may be applicable to other zoologic collections, particularly those with extensive, multispecies stocking systems. The effect of species-specific susceptibility has been controlled for analytically, but cannot be specifically quantified by our study. Such differences likely exist, and estimated lifetime incidences ranging from 0.7% in red deer to 32.1% in springbok have been reported 21 in the same collection, which may be partly the result of variable susceptibility to infection between species, but this factor has not been comprehensively evaluated in either captive or free-ranging wild ruminant species.
The zoo environment is uniquely challenging for infection control given the interaction and habitat sharing of multiple species, many of which have been comparatively little studied. The results of our study document the association of specific types of high-risk contact with the transmission of Johne’s disease. Our results indicate that a targeted surveillance system for Johne’s disease in the zoo setting should consider, in particular, targeting individuals that have shared an enclosure with an infected animal of the same species during the first week of life, in addition to the progeny of infected females. Infection risk may also be further elevated if many separate contact events occurred, or the infected animal was in the late stages of infection, or has lesions indicative of Johne’s disease at autopsy. The declining disease risk over time indicates that a systematic test-and-remove procedure can be effective in reducing transmission risk. We hope that the findings of our study can improve the effectiveness and selectivity of Johne’s disease surveillance in zoologic collections.
Footnotes
Acknowledgements
We thank Dr. Ilse Stalis (ZSSD) for data retrieval; PJ Stanley Hamel, Lance Aubrey, and a number of anonymous keepers for contributions to creating the home range data; Drs. Phillip Kass and Heejung Bang (UC Davis) for advice on statistical analysis; and the many keepers, managers, veterinarians, and laboratory personnel for their years of dedication to data acquisition and recording. Study sponsors did not contribute to study design, collection, analysis, and interpretation of data, manuscript preparation, or the decision to submit for publication.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Funding for this study was provided by the Ellen Browning Scripps Foundation and the Sheldon family. One author (TB) received a travel award from the Graduate Students Association, University of California, Davis used only for travel costs associated with presenting this work at the annual conference of the American Association of Zoo Veterinarians.