
Abstract
Mycoplasma bovis is emerging as an important pathogen of farmed bison in North America and is associated with high morbidity and mortality in affected herds. We developed an in-house ELISA to detect antibodies against Mycoplasma spp. in bison sera. The aims of the study were to estimate the seroprevalence against Mycoplasma spp. in bison herds with or without past history of M. bovis–associated disease, and to determine potential risk factors for seropositivity to Mycoplasma spp. in farmed bison in western Canada. A total of 858 serum samples were collected from bison >1 y of age from 19 bison herds. The individual and herd-level seroprevalence of Mycoplasma spp. was 12% and 79%, respectively. The proportion of seropositive animals was 0–41% and 0–9% for herds with or without a history of M. bovis–associated disease, respectively. Mycoplasma spp. appear to be widespread in bison in Manitoba, Saskatchewan, and Alberta. Eight of 11 herds with no history of M. bovis–associated disease were seropositive for Mycoplasma spp., which suggests that bison can be subclinically infected with Mycoplasma spp., or that infection may be underdiagnosed. Although not specific to M. bovis, the in-house ELISA developed to detect antibodies against Mycoplasma spp. may prove to be a valuable herd-level screening tool, providing insight needed for the development of appropriate prevention and control measures for Mycoplasma-related disease in bison herds.
Introduction
Since the first anecdotal report of disease associated with Mycoplasma bovis in bison in 1999 (D Hunter, pers. comm., 2012), this pathogen has become a major health issue for farmed bison (Bison bison) in North America. M. bovis outbreaks in bison herds are associated with a high incidence of respiratory and joint disease that tend to become chronic and lead to death, particularly in mature and yearling bison.4,8,14,24 A 2016 study of M. bovis – associated outbreaks in western Canada revealed that 88% of affected herds had introduced new animals to the herd, on average 8 wk prior to detection of first clinical signs within the herd; clinically sick bison were detected for up to 2 y after the first outbreak; and subsequently, no clinical signs were detected in the herd, even though control measures were generally perceived as ineffective at the time of the outbreak. 5 This may indicate that affected bison herds were naive to M. bovis prior to the outbreak and that the introduction of subclinically infected bison led to the introduction and spread of M. bovis, high mortality of clinically affected bison, and some survivors potentially developing effective immunity. However, there are no peer-reviewed reports to date describing the seroprevalence of antibodies against Mycoplasma spp. in farmed North American bison.
Serologic studies of M. bovis have been conducted in cattle as screening assays for detection of herd-level M. bovis infection.2,11,12 A study comparing the performance of 2 commercial indirect ELISA kits developed for cattle sera to an in-house ELISA developed for bison sera concluded that 1 of the commercial ELISA kits was not adequate for the detection of antibodies against M. bovis in bison sera but the other warranted further study. 23 Therefore, we aimed to develop an in-house ELISA using antigens from M. bovis strains (Mb300, Mb304, and Mb331) isolated from farmed bison in a 2016 study. 28 We expected that the use of recent strains would minimize challenges caused by potential antigenic mismatch between the ELISA proteins and the specific anti–M. bovis antibodies resulting from the rapid changes in the expression and phenotype of M. bovis surface antigens such as surface lipoproteins.3,25,26 We used the assay to estimate the seroprevalence against Mycoplasma sp. in bison herds affected with clinical disease associated with M. bovis infection and in herds with no history of M. bovis–associated disease, and to determine potential risk factors for higher Mycoplasma sp. antibody titers in farmed bison.
Materials and methods
Herds, animals, and sampling
Our study was approved by the Conjoint Health Research Ethics Board and Veterinary Sciences Animal Care Committee of the University of Calgary. Bison herds were selected from a previously reported case-control study of M. bovis outbreaks in the last 5 y in western Canada. 5 A control herd was defined as a farmed bison operation without clinical disease and with laboratory tests of bison mortalities negative for M. bovis infection during the same time period. In control herds, 60 animals were to be sampled in order to be 95% confident the herd was naive to Mycoplasma sp. A case herd was defined as a farmed bison operation with a laboratory-confirmed diagnosis of M. bovis and with disease onset between April 30, 2009 and April 30, 2015. In case herds, 40 animals were to be sampled to estimate the true prevalence with a precision of 20%, an expected prevalence of 30%, according to a preliminary survey conducted in 2011 and 2012 (unpublished data), and a 95% confidence level (EpiTools, http://epitools.ausvet.com.au). In both calculations, an estimated herd size of 100 bison was used (Statistics Canada, https://goo.gl/p4OGD5).
Between October 2014 and May 2015, blood was collected from a convenience sample of bison >1 y of age at bison operations where appropriate handling facilities were available or when bison were slaughtered at a nearby abattoir. A total of 858 blood samples were collected from farmed bison in 19 herds located in western Canada (Manitoba, Saskatchewan, Alberta, and British Columbia). All sampled animals were apparently healthy, with no obvious clinical signs of M. bovis–associated disease at the time of sampling. Mean sample size was 38 sera (SD = 15.0) in case herds and 50 sera (SD = 14.4) in control herds. Given the risks associated with handling bison, herd visits were arranged for a time when producers were handling the herd for other purposes (e.g., processing bison, winter pregnancy checking), which sometimes limited the number of bison available for sampling. Blood was collected from the coccygeal or jugular vein in 10-mL tubes without anticoagulant (Vacutainer, Becton Dickenson, Franklin Lakes, NJ) and refrigerated until processing. Serum was separated by centrifugation at 1,200 × g for 20 min within 24 h of collection. Serum aliquots were harvested and stored at −80°C until ELISA analysis.
Data concerning herd-level characteristics of each operation were obtained from an on-farm survey, 5 which collected information regarding type of bison operation (cow–calf herd, feedlot, or combined operation), whether M. bovis had been detected previously in bison on the operation, the clinical presentation of the disease, and various management practices and herd demographics. Combined operations included those with any combination of cow–calf, backgrounding, or feedlot production units.
Preparation of M. bovis whole-cell extract and membrane fractions
Three previously characterized M. bovis isolates were used. 28 These isolates were obtained from tissue samples from bison that died from disease associated with M. bovis infection: Mb300 from lung tissue of a bison in Saskatchewan, Mb304 from lung tissue of a bison in Manitoba, and Mb331 from joint fluid of a bison in Alberta. A standardized protocol was used to process and characterize the samples as published previously.21,24
Each of the selected bison isolates (Mb300, Mb304, and Mb331) was grown in 100 mL of modified Hayflick broth (PPLO [pleuropneumonia-like organism] broth + 0.01% yeast extract + 0.1 mg/mL ampicillin + 20% horse serum) at 37°C in 5% CO2 for 48 h. Whole-cell extracts were prepared as described previously. 18 The bacteria were collected by centrifugation at 4°C at 3,000 × g for 15 min. The cell pellet was washed twice in 100 mL of phosphate-buffered saline (PBS) and suspended in 0.5 mL of lysis buffer (9 M urea, 1% dithiothreitol, 2% 3-[(3-cholamidopropyl) dimethylammonium]-1-propanesulfonate, 2% Pharmalyte 3–10 [Sigma-Aldrich, St. Louis, MO], and protease inhibitors [Sigma-Aldrich]). The cells were disrupted by sonication and the lysate cleared by centrifugation at 30,000 × g at 4°C for 1 h. The cleared supernatant containing whole-cell extract was quantified for total protein concentration, aliquoted, and stored at −80°C until further use.
For preparation of membrane fractions from M. bovis isolates, a Triton X-114 fractionation method was used. M. bovis cultures were grown and pelleted as done for the preparation of whole-cell extracts; however, only a single wash with 50 mL of PBS was performed on the pellets. The M. bovis pellets were collected by centrifugation and suspended in 2 mL of fractionation buffer (Tris-buffered saline [TBS] containing 1% Triton X-114 [Sigma-Aldrich] and 1 mM phenylmethylsulfonyl fluoride [PMSF; Sigma-Aldrich]). The pellets were suspended gently and incubated for 30 min on ice. The detergent phase fraction was separated by centrifugation at 14,000 × g, 4°C for 20 min. Five-minute incubation at 37°C was done before separation of phases by centrifugation for 5 min at 8,000 × g at 4°C. The detergent phase was recovered and washed twice using TBS containing 1 mM PMSF, and 10 volumes of methanol were used to precipitate membrane proteins followed by suspension in 2 mL of lysis buffer. The M. bovis membrane fractions were quantified for total protein concentration by standard methods, aliquoted, and stored at −80°C until use in ELISA experiments as described below.
Immunoblots
Immunoblotting of ELISA antigens (whole-cell extracts and membrane fractions) prepared from M. bovis isolates was carried out using bison sera with high and low titers against M. bovis. The high-titer bison serum was collected from an experimentally infected bison 28 d after infection, as described elsewhere (Register K, et al. 2015. Relative virulence in bison and cattle of bison-associated genotypes of Mycoplasma bovis. Available at: https://goo.gl/fSxOHi), and shown to be strongly seropositive in an in-house ELISA developed for bison sera. 23 The low-titer pooled bison sera were collected from 5 bison at the Western College of Veterinary Medicine, Specialized Livestock Research Facility (WCVM-SLRF; University of Saskatchewan, Canada). This bison herd has been closely monitored for the last 10 y: there have not been any clinical cases of respiratory disease, and all mortalities have undergone postmortem examination, of which, none have demonstrated pulmonary lesions.
The antigens (10 μg per lane) from bison isolates (Mb300, Mb304, and Mb331) were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to nitrocellulose membranes, and blocked with 3% bovine serum albumin for 1 h at room temperature. The blocked nitrocellulose membranes were washed 3 times using TBS with 0.05% Tween-20 (TBST) and later incubated with 1:500 dilution of specific bison sera for 1 h at room temperature. Three washes were carried out again using TBST, and 1:5,000 dilution of the phosphatase-labeled goat anti-bovine IgG (H+L) secondary antibody (ReserveAP, KPL, Gaithersburg, MD) was added and incubated for 1 h at room temperature. The nitrocellulose membranes were washed once with TBST and then twice with alkaline phosphatase (AP) buffer. The membrane was incubated with AP buffer containing 165 μg/mL of nitro blue tetrazolium chloride (Sigma-Aldrich) and 165 μg/mL of 5-bromo-4-chloro-3’-indolyphosphate p-toluidine salt (Sigma-Aldrich) until color appeared (1–2 min). The reaction was stopped by the addition of distilled water, and nitrocellulose membranes were dried for further analysis.
ELISA protocol
The ELISA protocol was previously optimized for detection of antibodies to M. bovis in cattle serum samples. 21 Based on a standardized in-house ELISA protocol, further modifications were done to optimize it for bison serum samples. Plates with 96 wells (Immulon U-bottom plates, Thermo Fisher Scientific, Rochester, NY) were coated with 2 μg/mL of total antigen (M. bovis bison isolates Mb300, Mb304, and Mb331 whole-cell extracts or membrane fractions) in coating buffer (100 μL/well), then sealed and incubated at 4°C overnight. Plates were washed 5 times with TBST, followed by addition of 200 μL of TBST with gelatin (TBST-G; 5 mL of 10% gelatin in 100 mL of TBST), as a blocking agent, to each well. Incubation was done for 2 h at room temperature followed by 5 washes with TBST. Bison serum samples were serially diluted with starting dilution at 1:100 in TBST-G, followed by a 4-fold serial dilution conducted 6 times in TBST-G (1:100, 1:400, 1:1,600, 1:25,600, 1:102,400, and 1:409,600). These serum dilutions were prepared in separate macro-well strips, and 100 μL was transferred to coated ELISA plates and incubated for 2 h at room temperature. Plates were washed as before, and 100 μL per well of phosphatase-labeled goat anti-bovine IgG (H+L) secondary antibody (KPL) was added at 1:5,000 dilution in TBST-G. Incubation was done for 1 h at room temperature followed by 3 washes. The substrate P-nitrophenyl phosphate (KPL) was prepared at 1 mg/mL in diethanolamine buffer (added 100 μL per well) and incubated for 45 min at room temperature. Plates were read using a microplate spectrophotometer (xMark, Bio-Rad, Philadelphia, PA) at 405 nm (A450) with reference filter at 490 nm. The serum titers for samples were calculated by the intersection of least-square regression of A405 versus logarithm of dilution as described previously. 21
Reference samples to determine cutoff values
A negative reference sample of 27 sera from bison was randomly selected from among the 46 available animals in the bison herd at WCVM-SLRF. This herd was considered to be disease-free, as described above, thus these animals were classified as presumed uninfected according to published standards. 13
A positive reference sample of 50 sera was collected 68 d after bison were vaccinated twice, 21 d apart, with an autogenous M. bovis vaccine (Gallant Custom Laboratories, Cambridge, ON, Canada) on arrival at an operation in Alberta. Although not an ideal reference sample, sera from the vaccinated animals was the closest to a positive reference sample available at the time.
Statistical analyses
The procedure used to determine the cutoff values was adapted from the World Organization for Animal Health (OIE) methods of validation of assays.13,30 In our study, overlap between some sera in the negative reference sample with some in the positive reference sample led to the selection of multiple cutoff values, thus defining a seronegative, suspicious, and seropositive serologic status for Mycoplasma sp. of the sampled bison. The cutoffs were defined as percentiles of the reference samples antibody titers corresponding to 93% diagnostic specificity (DSp) and 99.9% diagnostic sensitivity (DSe). The small sample size of the reference samples led to DSp and DSe with ±4% acceptable error and 97% confidence interval (CI). 30
Descriptive analyses of predictor variables were conducted, and chi-square tests were employed to investigate the relationship between each predictor variable and the outcome variable of Mycoplasma sp. serologic status of bison with 3 ordered levels: seronegative, suspicious, and seropositive. Univariable ordered logistic regression model building was conducted while accounting for the clustering of serum samples by herd with robust standard errors. 6 The Brant test was used to assess the proportional odds assumption (equal relationship between each pair of outcome levels). Independent variables with p < 0.10 were considered for a multivariable ordered logistic regression model, and backward elimination was used to remove variables with p > 0.05. Data were analyzed using STATA v. 13.0 (StataCorp, College Station, TX), and graphs were drawn with open source software R v. 3.2.2 (https://www.r-project.org/). A p < 0.05 was considered statistically significant.
Results
Immunoblot analysis of selected bison sera
Immunoblot analysis was performed to evaluate the antigens used for coating the ELISA plates, including whole-cell extracts and membrane fractions prepared from M. bovis bison isolates (Fig. 1). These antigens were tested for their specific interaction with selected high-titer bison serum pool and low-titer bison serum pool. A qualitative analysis of immunoblots revealed that the majority of proteins were strongly recognized by the high-titer serum pool (Fig. 1A), whereas a low background interaction was observed in the immunoblot using the low-titer serum pool (Fig. 1B).
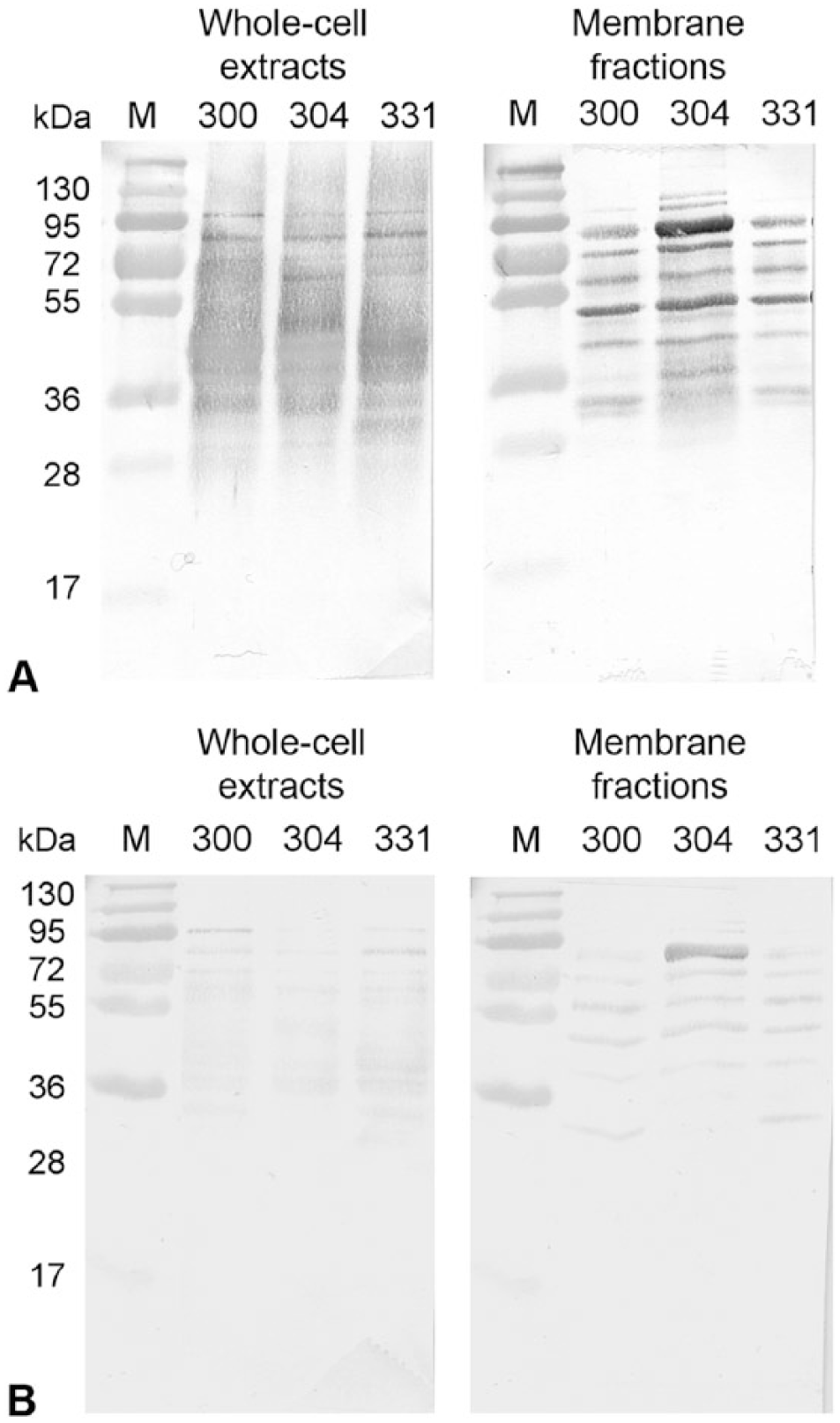
Immunoblotting of selected bison sera with ELISA antigens prepared from selected Mycoplasma bovis bison isolates (Mb300, Mb304, and Mb331). Membranes containing either whole-cell extracts or membrane fractions were incubated with a 1:500 dilution of bison sera having either a high titer against M. bovis
Selection of cutoff values
Visual assessment of the frequency distribution of Mycoplasma sp. antibody titers of the reference samples (Fig. 2A) indicated a substantial overlap between values from presumed uninfected and vaccinated animals. Thus, 2 cutoff values were selected: 1) at titers >42,001, which included 99.9% of the vaccinated reference sera; and 2) at titers <266,283, which included 93% of the presumed uninfected reference sera. Serum titers <42,001 were considered negative, those between 42,001 and 266,283 were considered suspicious, and those >266,283 were considered positive. The multiple cutoff values were assessed by visual inspection of the frequency distribution of Mycoplasma spp. antibody titers from farmed bison in western Canada, which had a trimodal distribution (Fig. 2B).
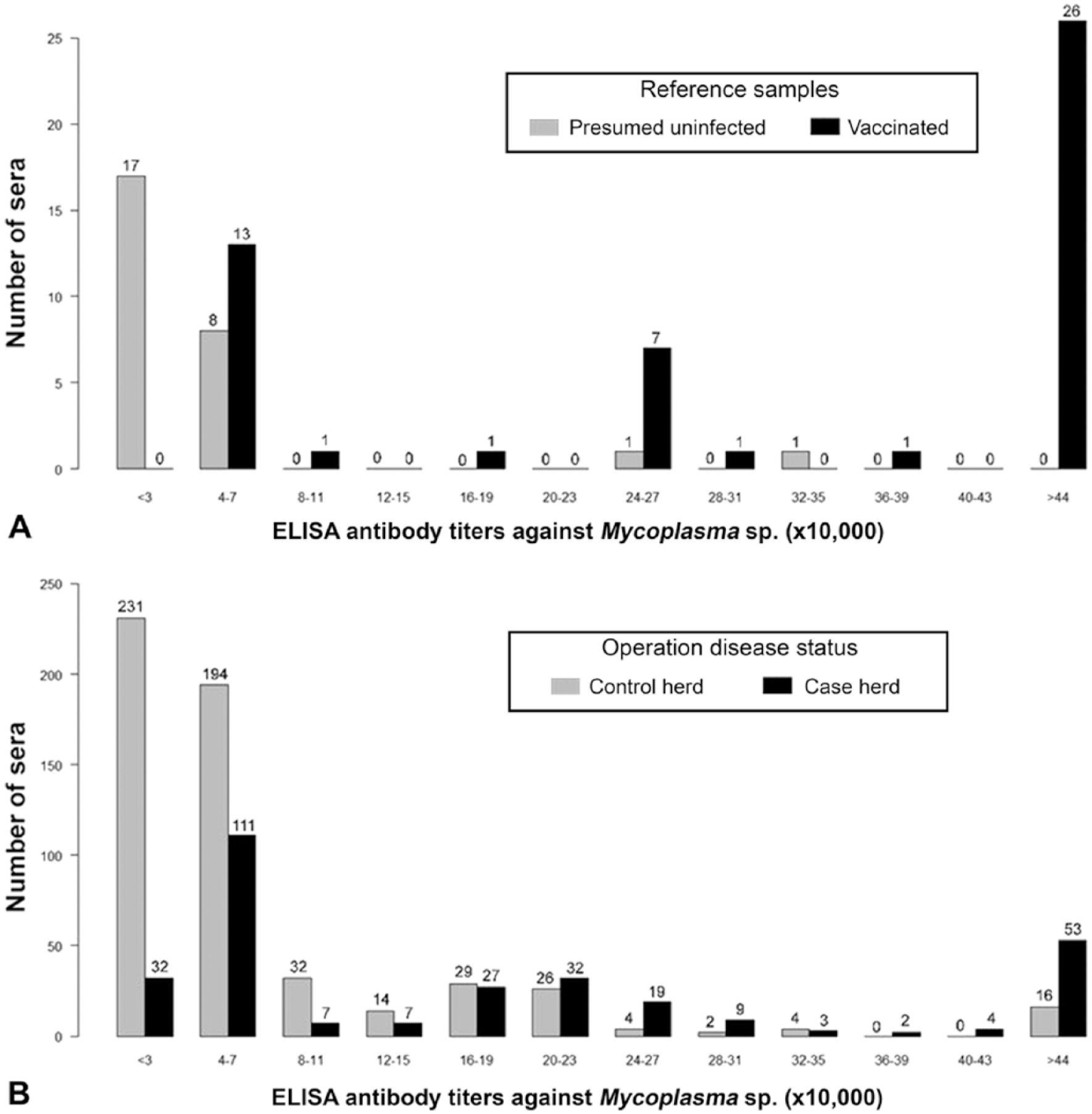
Distribution of antibody titers against Mycoplasma spp. of farmed bison in western Canada collected from October 2014 to May 2015.
Seroprevalence of Mycoplasma spp. and relationship with herd-level M. bovis–associated disease status
Of the 858 samples, 101 (12%) were seropositive, 485 (57%) were suspicious, and 272 (32%) were seronegative to Mycoplasma spp. The percentages of seropositive, suspicious, and seronegative bison within the herds were 0–41%, 9–82%, and 0–91%, respectively. If a herd had at least 1 seropositive bison, it was considered positive; thus, 15 (79%) of the 19 sampled herds were classified as Mycoplasma-positive. However, herds classified as positive or negative based on the serologic results were not concordant with known M. bovis–associated disease status for 1 of the 8 case herds and 8 of the 11 control herds (Fig. 3). If a threshold of 3% seropositive animals was used instead to classify a herd as positive, only 1 herd changed classification (data not shown). Because interpretation of the data did not change with the higher herd-level threshold, the more stringent threshold of 1 positive animal was reported. Visual inspection of the frequency distribution of Mycoplasma spp. antibody titers of bison by M. bovis–associated disease status of the herd (Fig. 2B) shows a wide range of titers within both case and control herds. The proportion of seropositive animals within case herds was 0–41%, and within control herds the proportion of seropositive animals was 0–9%. The odds of bison having a higher serologic status (seronegative < suspicious < seropositive) were 2.0 times (95% CI: 1.0–3.0) greater in bison from case herds than those from control herds (p < 0.01; Table 1).
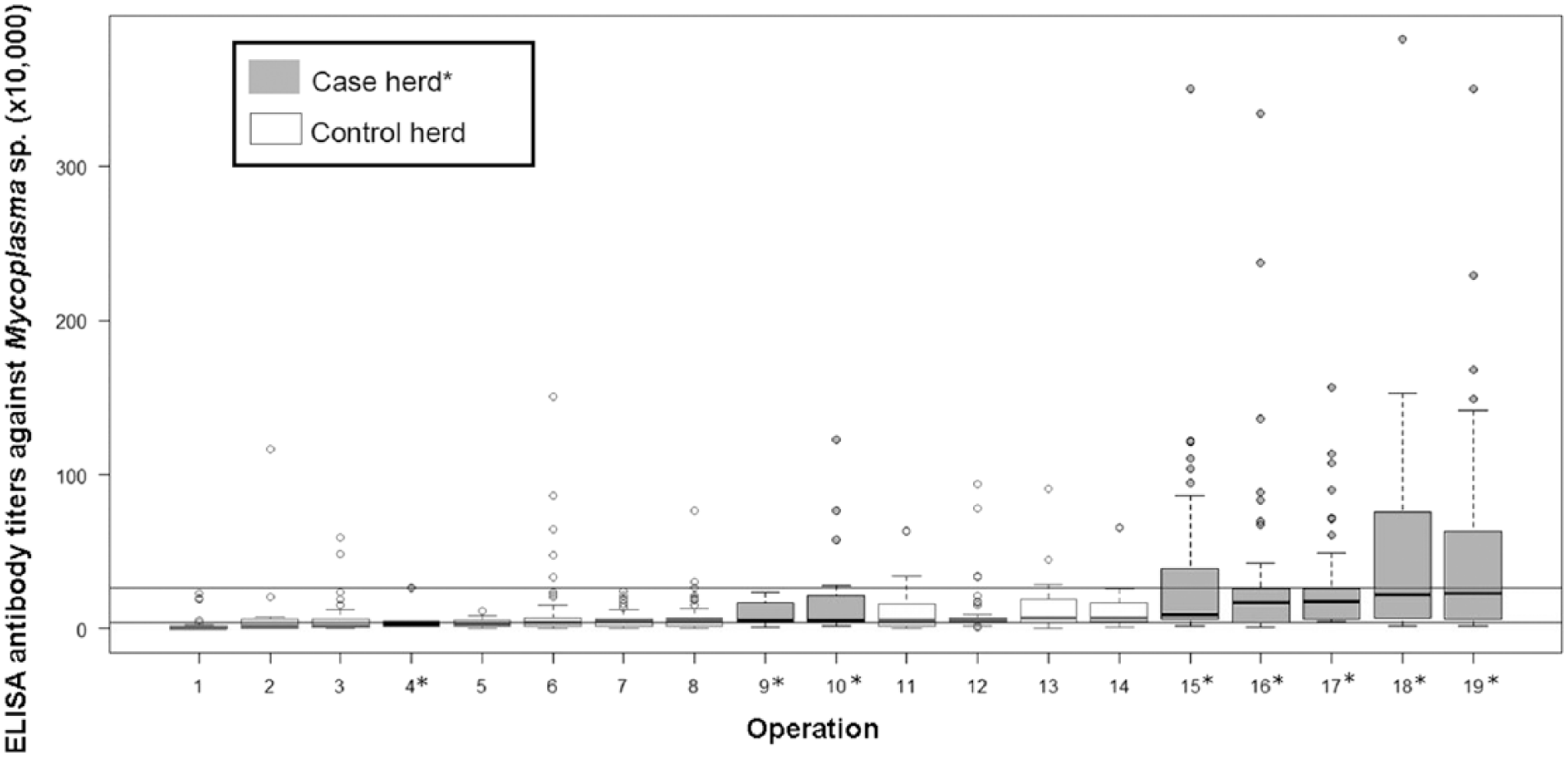
Distribution of antibody titers against Mycoplasma sp. within farmed bison operations in western Canada, in ascending order of median titers within a herd, by M. bovis–associated disease status of the operation in the preceding 5 y. Horizontal lines separate seronegative, suspicious, and seropositive titers, indicated by cutoffs at titers of 42,001 and 266,283.
Associations between herd-level characteristics and classification of bison antibody titers against Mycoplasma spp. assessed using univariable ordered logistic regression models with robust standard errors (adjusted for clustering by herd) in sera collected between October 2014 and May 2015 from farmed bison in western Canada.*
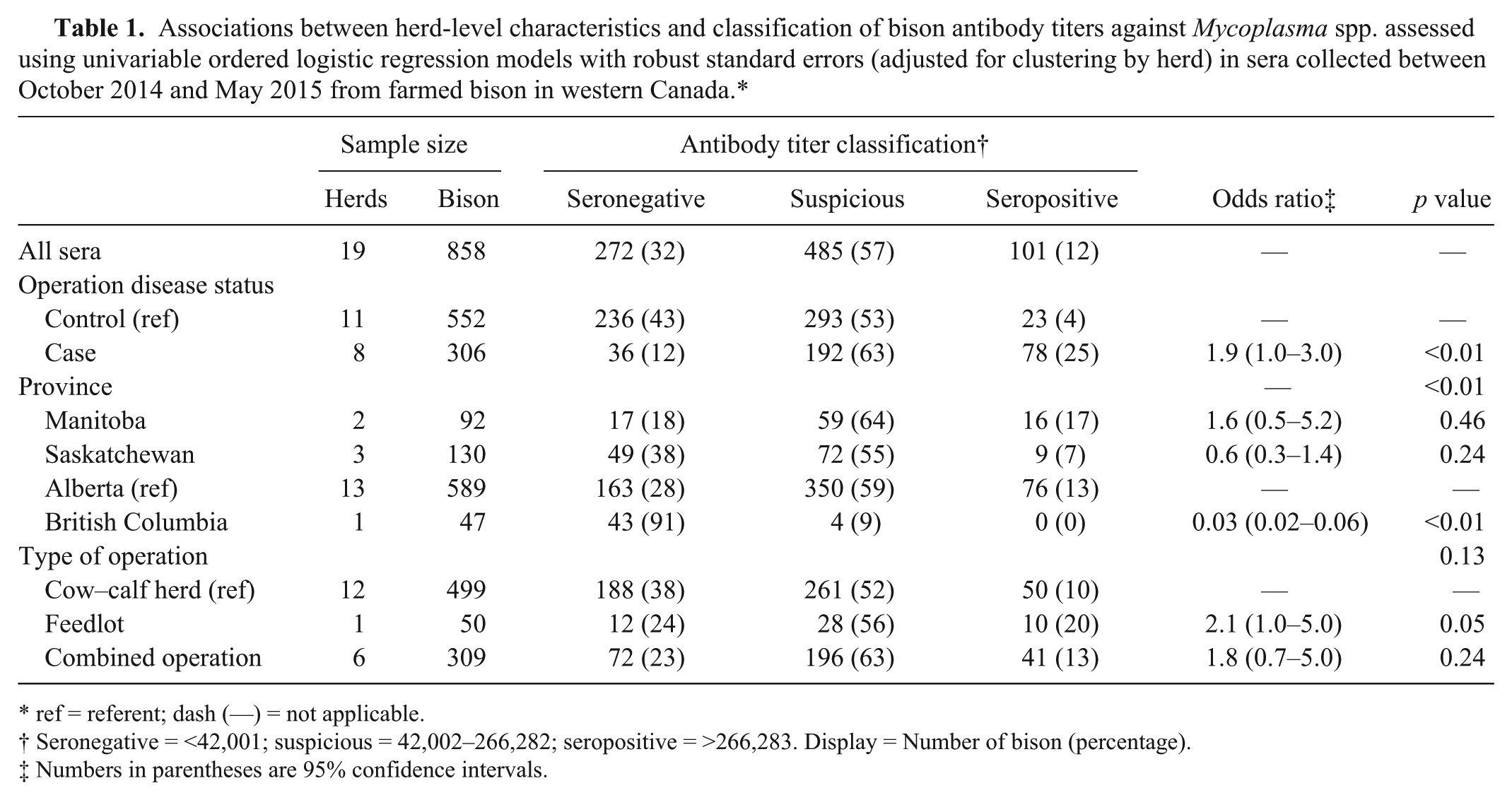
ref = referent; dash (—) = not applicable.
Seronegative = <42,001; suspicious = 42,002–266,282; seropositive = >266,283. Display = Number of bison (percentage).
Numbers in parentheses are 95% confidence intervals.
Individual and herd-level risk factors
The association between Mycoplasma spp. serologic status of bison and herd-level factors was explored with univariable ordered logistic regression models, adjusted for clustering by herd (Table 1). The one herd sampled in British Columbia had no seropositive bison. Thus, the odds of bison having a higher serologic status were lower for bison raised in British Columbia than in Alberta (odds ratio [OR] = 0.03, p < 0.01).
Information regarding age and sex of the bison was not available for all samples collected given constraints of time, labor, or both during sample collection. For 320 samples from 10 herds, the sex and age of the bison was recorded, with the majority of the samples (83%) being collected from females over 30 mo. One male over 30 mo was excluded from further analysis because it was the only animal sampled from this age and sex category. A multivariable ordered logistic regression model, adjusted for clustering by herd, was determined (Table 2). The odds of having a higher Mycoplasma sp. serologic status (seronegative < suspicious < seropositive) was 7.0 times (95% CI: 2.5–20.2) higher in bison belonging to a case herd than a control herd (p < 0.01), after accounting for sex and age of bison. There was a significant difference in Mycoplasma sp. serologic status of sera by sex and age of the bison after accounting for herd disease status (p < 0.01). The odds of having a higher serologic status were lower in female bison <30 mo of age than females >30 mo (OR = 0.3, 95% CI: 0.2–0.6, p < 0.01). Also, the odds of having a higher serologic status were 3.0 times (95% CI: 1.6–5.3) higher in male bison <30 mo of age than in females >30 mo (p < 0.01).
Associations between individual and herd-level characteristics and classification of bison antibody titers against Mycoplasma spp. assessed using multivariable ordered logistic regression models with robust standard errors (adjusted for clustering by herd) in sera collected between October 2014 and May 2015 from farmed bison in western Canada.*
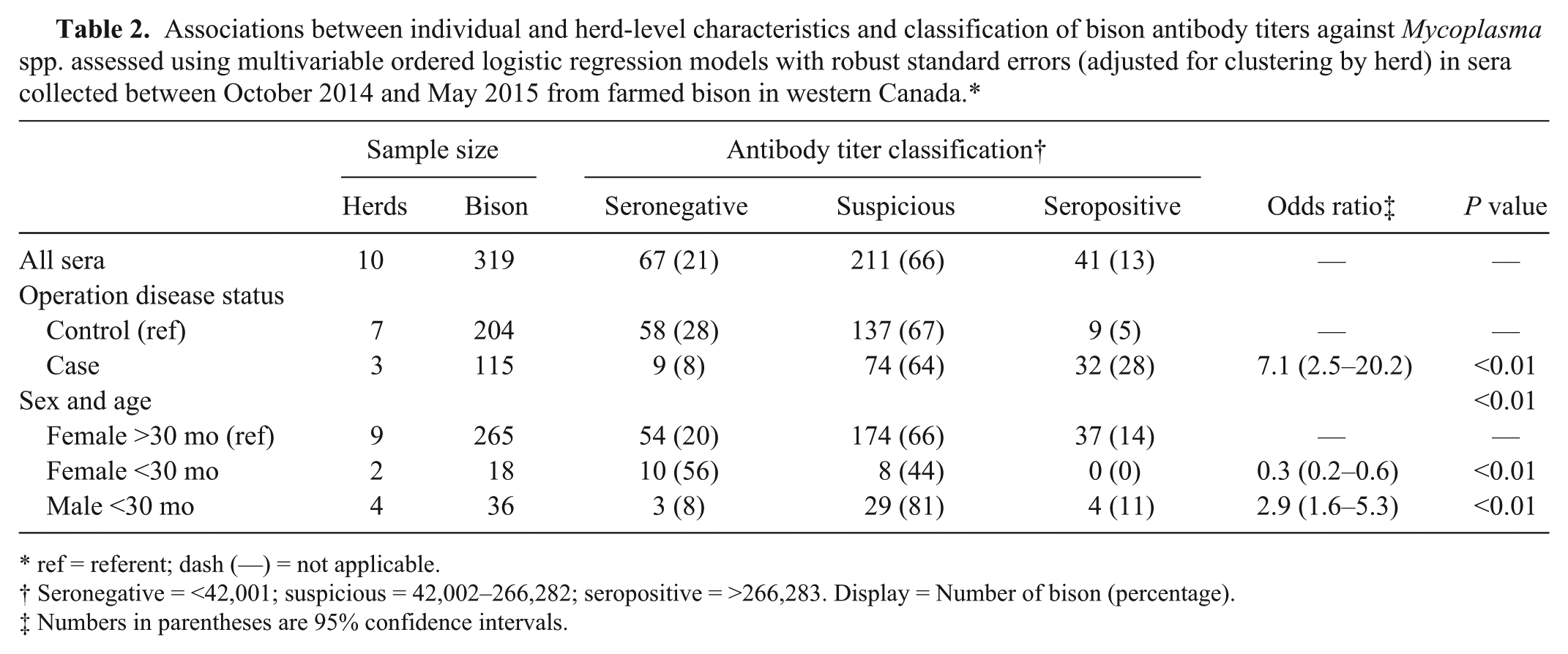
ref = referent; dash (—) = not applicable.
Seronegative = <42,001; suspicious = 42,002–266,282; seropositive = >266,283. Display = Number of bison (percentage).
Numbers in parentheses are 95% confidence intervals.
Discussion
Currently, the most accurate means of diagnosing M. bovis infection in bison is with postmortem sampling and subsequent laboratory testing using Mycoplasma culture and PCR.16,19 Detection tools that can be used for live animals, such as serology, would be invaluable for efficient screening of M. bovis infection in bison herds.15,23 Our newly developed ELISA was not specific for M. bovis because sera against M. agalactiae, M. bovigenitalium, or M. bovirhinis obtained from the Mollicutes Collection of Cultures and Antisera (World Federation of Culture Collections’ World Data Centre for Microorganisms, collection 858) reacted weakly with the protein extract and membrane fractions (data not shown). Despite this limitation, it is likely that the high serum titers observed in some animals are the result of recent M. bovis exposure or infection. This conclusion is based on the results obtained with a similar ELISA performed with sera from cattle that were experimentally challenged with M. bovis.18,22 In those studies, pre-challenge serum titers against M. bovis were similar to the titers considered to be seronegative in our study. After challenge, those titers18,22 significantly increased to values equal to or higher than the cutoff for positive animals used in our study.
An important step during the validation of an assay is the selection of reference samples to establish cutoff value(s). Ideally, a larger number of reference samples would be selected based on known history and M. bovis infection status.13,30 The negative reference samples in our study were selected based on historical freedom from M. bovis–associated infection and prolonged closed-herd status. However, no clinically diseased bison were available during the study period in the target population, so a vaccinated reference sample was used. Furthermore, the stress associated with the necessary handling and restraint for sample collection would have been prohibitive in sick bison. Usually, reference samples are collected from vaccinated animals consecutively, over months, in order to determine the kinetics of the antibody response and the point at which antibodies are first detectable by the assay. 13 However, in our study, the goal of the vaccinated reference sample was to provide a framework of antibody titers as close to what would be seen in a natural M. bovis infection as possible. This approach has inherent limitations given the unknown efficacy of the autogenous vaccine and factors that affect antibody titers, such as strain used, dose, and method of administration.30,27 Thus, the DSe determined with the vaccinated reference sample may be a poor estimate of the true DSe of the assay. However, an alternative positive reference sample population was not feasible.
A limitation of the validation process of the present ELISA was the low number of presumed uninfected and vaccinated bison available. Guidelines for the validation of an assay recommend a reference sample of at least 100 sera.10,13 A smaller sample size may lead to inaccuracy of the assay because the sample does not appropriately represent variables that influence the production of antibodies (e.g., sex, age, stage of infection, cross-reactions with other infectious agents) in the target population.13,20 Thus, as proposed by the OIE manual for validation of diagnostic assays, 30 the validation assessment conducted in our study produced provisional estimates of the DSe and DSp of the assay, with further evaluation of serum samples needed in the future to update these estimates.
Two cutoff values were established because of overlap of the frequency distribution of the uninfected and vaccinated reference samples, so that the lower cutoff titer of 42,001 defined a 99.9% DSe and the higher cutoff titer of 266,283 defined a 93% DSp. The use of multiple cutoff values was assessed through the frequency distribution of antibody titers from the study sample and was visually determined to be sensible in order to optimize sensitivity and specificity.
The serologic study results for the 858 serum samples indicate that the individual-level seroprevalence of Mycoplasma spp. was 12% and the herd-level seroprevalence was 79%. These findings were very similar to results from studies carried out in cattle,1,2 particularly a seroprevalence study conducted in France involving suckling beef cattle 11 where the proportion of seropositive animals was low (2–13% depending on the county) and the herd-level seroprevalence of M. bovis was high (28–90% depending on the county). Such findings suggest that a few seropositive animals were present in a relatively high number of herds. However, none of the sampled bison in our study showed evidence of clinical disease at the time of sampling, which may have contributed to the low number of seropositive bison. An M. bovis seroprevalence study of Polish cattle found that clinically healthy cattle had a considerably lower individual seroprevalence than cattle with respiratory disease (16% and 43%, respectively). 2 Moreover, a study of European bison (Bison bonasus) with clinical signs associated with M. bovis found 21% of sampled animals to be seropositive. 7
Although no other laboratory tests were performed to determine the true Mycoplasma spp. infection status of the sampled bison, we expected that herd-level seroprevalence would reflect the known M. bovis outbreak history of the herd, showing evidence of past exposure to infection with M. bovis. 2 Historical outbreaks of M. bovis–associated disease occurred within the previous 5 y in 8 of the herds sampled, and all but 1 were classified as seropositive. However, 8 of 15 herds with no history of M. bovis outbreaks in the past 5 y were considered seropositive with at least 1 seropositive animal. This disparity between disease and herd immune status may be the result of the under-diagnosis of M. bovis–associated disease, the presence of bison subclinically infected with M. bovis or other cross-reactive species, or previous exposure resulting in prolonged serologic immunity in control herds. 4 Regardless, bison with a higher serologic status were more likely to belong to a case herd than a control herd.
In 2011, the Canadian Agricultural Census reported a total of 1,102 bison operations in Canada, 91% of which were located in western Canada (https://goo.gl/p4OGD5). Thus, our study collected sera from bison in the western provinces. However, bison from only 19 operations were sampled, which may not be representative of the entire Canadian farmed bison population. There were no case herds in British Columbia, which may in part explain why animals from British Columbia were less likely to have higher serologic status compared with animals from Alberta. However, because only one herd was sampled in British Columbia, conclusions are limited.
Our study targeted bison >1 y of age because previously published reports in bison suggest that mature bison cows are more likely to be affected, followed by yearlings.4,8,9,14,24 However, the mechanism(s) by which mature bison cows are seemingly more predisposed to M. bovis–associated disease are, as yet, unclear. Our study found that, compared with adult cows, females <30 mo of age were more likely to have a low serologic status. This may be because young females had fewer opportunities to be exposed to Mycoplasma spp. and thus to develop detectable antibodies than adult cows. However, males <30 mo of age were more likely to have a higher serologic status than adult cows. This may be because males are more likely to be transported to a feedlot to be fed for slaughter rather than being kept on ranch as breeding stock. The introduction of new bison from different sources and a higher density of animals may lead feedlots to be at higher risk of disease introduction and spread of infectious agents compared to cow–calf operations.5,17 Moreover, a serology study in cattle found that the seroprevalence of M. bovis increased from 27% at arrival of bovine calves at a feedlot to 98% 1 mo later. 12 A study of European bison found seropositive male bison (4%), but none of the females sampled had detectable antibodies against M. bovis. 29
Footnotes
Acknowledgements
We thank the bison producers for participating on this study. We thank our collaborators Drs. Pat Burrage, Roy Lewis, Ted Clark, Rob Tremblay, Eugene Janzen, and Jennifer Davies; students from Utrecht University, Thom Oostenbrug and Dirk van Aaken; Dr. Melissa Moggy from the University of Calgary Faculty of Veterinary Medicine; Dr. Shelagh Copeland from Manitoba Agriculture Food and Rural Development; Drs. Sandy Stafford, Gordon Earnshaw, and Dennis Schonheiter from Alberta Agriculture and Forestry; Dr. Jesus Palomino Cano from Western College of Veterinary Medicine, Saskatoon; Ms. Hailey Marwood from the University of Saskatchewan; and the Canadian Bison Association for their support of the project.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors received financial support for this project from the Alberta Livestock and Meat Agency Ltd.