
Abstract
Intoxication with Solanum bonariense in cattle causes cerebellar cortical degeneration with perikaryal vacuolation, axonal swelling, and death primarily of Purkinje cells, with accumulation of electron-dense residual storage bodies in membrane-bound vesicles. The pathogenesis of this disease is not fully understood. Previously, we proposed that inhibition of protein synthesis in Purkinje cells among other altered metabolic pathways could lead to cytoskeletal alterations, subsequently altering cell-specific axonal transport. In the present study, immunohistochemical and histochemical methods were used to identify neuronal cytoskeletal alterations and axonal loss, demyelination, and astrogliosis in the cerebellum of intoxicated bovines. Samples of cerebellum from 3 natural and 4 experimental cases and 2 control bovines were studied. Immunoreactivity against neurofilament (NF)-200KDa confirmed marked loss of Purkinje neurons, and phospho-NF protein, β-tubulin, and affinity reaction against phalloidin revealed an altered perikaryal distribution of neuronal cytoskeletal proteins in the remaining Purkinje cells in intoxicated cattle. Reactive astrogliosis in every layer of the cerebellar cortex was also observed with anti–glial fibrillary acidic protein immunohistochemistry. In affected cattle, demyelination and axonal loss in the cerebellar white matter, as well as basket cell loss were demonstrated with Klüver–Barrera and Bielschowsky stains, respectively. Based on these results, we propose that neuronal cytoskeletal alterations with subsequent interference of the axonal transport in Purkinje cells may play a relevant role in the pathogenesis of this neurodegenerative disorder, and also that demyelination and axonal loss in the cerebellar white matter, as well as astrogliosis in the gray matter, likely occur secondarily to Purkinje cell degeneration and death.
Introduction
Solanum bonariense L. (family Solanaceae) is a perennial shrub endemic in Uruguay, Argentina, Brazil, and Paraguay. Spontaneous cases of S. bonariense intoxication have been reported in dairy cattle in Uruguay, 30 and beef cattle in Argentina, 20 causing economic losses in the agricultural sector. The disease has been experimentally reproduced in cattle, and the clinical signs and pathology have been characterized. 30 Affected cattle develop a syndrome characterized clinically by cerebellar neurological signs (ataxia, hypermetria, hyperesthesia, opisthotonus, and falling), and histologically by progressive Purkinje cell vacuolation, degeneration, and, eventually, death and loss. 30 Similar cerebellar cortical degeneration caused by genus Solanum has been described in Brazil in cattle intoxicated with S. fastigiatum1,26,27 (which is now considered a synonym of S. bonariense 32 ), S. paniculatum, 6 and S. subinerme, 14 and in South Africa in cattle fed S. kwebense. 24 Ultrastructural alterations in the cerebellar Purkinje cells of affected cattle consist of perikaryal and axonal accumulation of electron-dense residual storage bodies in membrane-bound vesicles (organelle accumulation). 30 A similarity with the pathogenesis of acquired lysosomal storage diseases has been proposed 27 ; however, the precise mechanisms leading to vesicular dilation and accumulation are unknown. The nature of the perikaryal vesicles and the biochemical composition of the intravesicular accumulations are not yet completely characterized,27,29 although the lectin-binding pattern is consistent with a glycolipid storage disease, 22 and the vacuolation is suggestive of a lysosomal disease. Ultrastructurally, the membranous cytoplasmic inclusions are similar to those seen in sphingolipidoses. 24 In plant-induced lysosomal storage diseases affecting the cerebellar Purkinje cells such as those caused by Astragalus spp., Oxytropis spp., Swainsona spp., and Sida spp., plant alkaloids (e.g., locoine, swainsonine) inhibit lysosomal enzymes (e.g., α-mannosidase) leading to the accumulation of their substrates (e.g., mannose). 36 However, such enzyme inhibitors have not been identified in S. bonariense, and a different pathogenic mechanism might be involved in the neurodegeneration induced by this plant. 32
Axonal transport systems require a well-organized cytoskeleton to allow movements of molecules and cellular organelles from the cell body through the axon to the synapses, and of degradation products back to the cell body. Alterations in these anterograde and retrograde transport systems can lead to severe neuronal dysfunction. 36 In the present study, we characterize cerebellar Purkinje cell cytoskeletal alterations and propose that these changes may be important in determining Purkinje cell death and loss and other sequelae (astrogliosis, axonal swelling, myelin loss).
Material and methods
Three S. bonariense naturally intoxicated cattle (2 cows and a heifer), 4 experimentally intoxicated steers, and 2 control adult bovines (a cow and a steer) were used. Details of the experimental induction of the disease, the animals’ history, severity of clinical signs, and microscopic cerebellar lesions were previously reported. 30 Briefly, nervous signs, including periodic episodes of recumbency with inability to rise without loss of consciousness, ataxia, hypermetria, hyperesthesia, staggering gait, muscle tremors, head and thoracic limbs extension, opisthotonus, nystagmus, and in those animals most severely affected, falling to the side or backward are shown. These signs occurred spontaneously or are induced when affected animals become excited or intentionally stressed. 30 Tissue samples from selected areas of the cerebellum (3 from the rostral, medial, and caudal areas of cerebellar vermis, 1 from each lateral hemisphere, and 1 from the flocculonodular lobe) were obtained. Samples were immersion fixed in neutral buffered formalin (pH 7.4) for at least 48 hr, processed by standard histological methods, and embedded in paraffin for the production of 5-µm–thick tissue sections. Some section were stained with hematoxylin and eosin (HE) for light microscopic observation, while other were stained with Klüver–Barrera for myelin and neurons and Bielschowsky silver impregnation stain to asses myelin loss, and the integrity of basket cells (inhibitory GABAergic interneurons) and Purkinje cell axonal loss in the gray and white matter, respectively.
Immunohistochemical staining was performed to evaluate neuronal cytoskeletal alterations or reactive astrogliosis using mouse monoclonal antibodies. A summary of techniques, antibodies, and regents used is presented in Table 1.
Antibodies and reagents used to evaluate neuronal cytoskeletal alteration and reactive astrogliosis.*

NF = neurofilament; GFAP = glial fibrillary acidic protein; FA = fluorescent assay; FAT = fluorescent antibody test; HRP = horseradish peroxidase.
Sigma-Aldrich, St. Louis, MO; Covance, Princeton, NJ; Invitrogen, Life Technologies, Grand Island, NY; Dako Denmark A/S, Glostrup, Denmark.
The complex neuronal cytoskeleton is composed of 3 interconnected protein structures: actin microfilaments, intermediate filaments (neurofilaments), and microtubules (α and β-tubulin).9,23 The integrity of the neuronal cytoskeleton was evaluated using antibodies against total neurofilament (NF)-200KDa, the heavy molecular weight (MW) and highly phosphorylated neurofilament subunit, a and to specifically evaluate phosphorylation of neurofilaments, phospho-NF protein, b and β-tubulin for microtubules, c as well as Alexa Fluor 647–phalloidin for actin microfilaments, d while astrogliosis was assessed using an antibody against glial fibrillary acidic protein (GFAP). e For NF-200KDa, phospho-NF protein, β-tubulin, and GFAP, sections were antibody blocked with a solution of 50 mM glycine plus 10% normal goat serum and 0.1% bovine serum albumin (BSA) in phosphate buffered saline (PBS; pH 7.4) for 60 min and incubated with the primary antibody at 4°C overnight. Following 3-fold washing of the slides with PBS, samples were incubated with a peroxidase-labeled polymer system, f and diaminobenzidine as a chromogen (for NF-200KDa and GFAP) or with a fluorescein conjugated secondary antibody, dilution of 1:1,000 (for phospho-NF protein and β-tubulin) for 45 min. g Finally, the slides were washed 3 times with PBS, dried at room temperature, and mounted conventionally (for NF-200KDa and GFAP) or with an antifade reagent (for phospho-NF protein and β-tubulin). h For phalloidin, tissue sections were incubated for 45 min at room temperature, and then washed, dried, and mounted with an antifade reagent. Sections stained with HE and Klüver–Barrera or Bielschowsky stains, and immunolabeled with anti–NF-200KDa and GFAP were observed under a transmitted light microscope, i and sections with fluorescent dyes (phospho-NF protein, β-tubulin, and phalloidin) under a confocal microscope. j
Results
Histology, Klüver–Barrera, and Bielschowsky staining methods
Microscopic lesions seen with HE stain in the cerebella of spontaneous and experimental cases included in this study were published elsewhere. 30 The same neuroanatomic areas were subjectively compared with the cerebella of control animals, as previously reported.26,27,30,31 Briefly, the following changes were observed in the cerebellar Purkinje cells: swollen pale eosinophilic cell bodies with fine cytoplasmic vacuolation of the perikarya and/or 1 or more large clear cytoplasmic vacuoles, either nuclear distension with occasional nuclear displacement to the periphery of the neuronal body or nuclear pyknosis, neuronal loss, and formation of proximal axonal swellings (torpedoes) in the granular layer and cerebellar white matter. In all cases, all examined sections obtained from the rostral, medial, and caudal areas of cerebellar vermis, lateral hemispheres, and flocculonodular lobe were similarly affected.
To determine standard intensity values and establish a reference frame for comparison for Klüver–Barrera and Bielschowsky staining methods, we stained the same cerebellar areas in normal control and affected cases. In experimental and natural cases, dose-dependent cerebellar demyelination and Purkinje cell axonal loss, characterized by an increased paleness of the white matter, were identified by both staining methods (Figs. 1, 2). Bielschowsky stain confirmed that in affected cases, when cerebellar Purkinje cells had undergone advanced degeneration and death, the cellular processes of the neighboring basket cells had also disappeared, suggesting concomitant basket cell death (Figs. 3, 4). In less affected cases, no significant changes were noted in the density of basket cell processes.
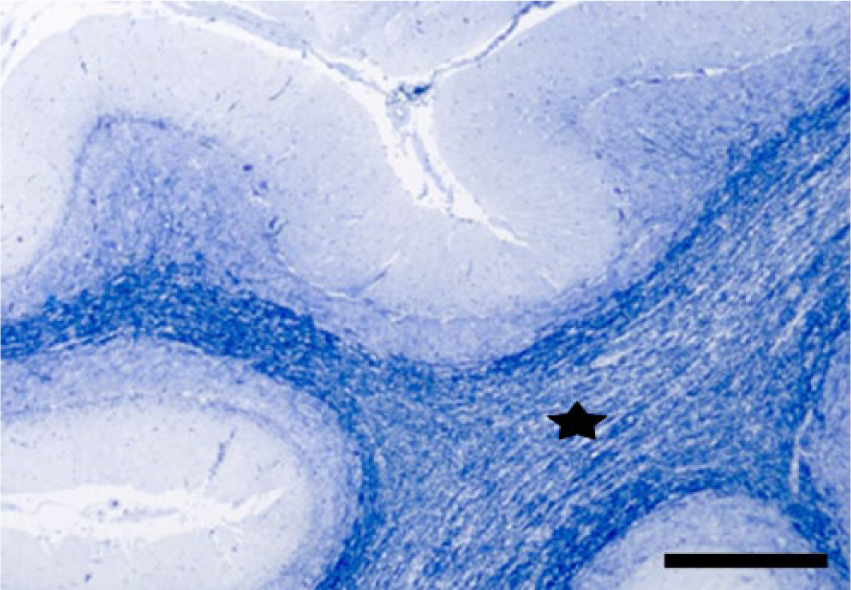
Cerebellum. Steer experimentally intoxicated with Solanum bonariense. Demyelination in the white matter of cerebellar folia (star). Klüver–Barrera stain. Bar = 500 µm.
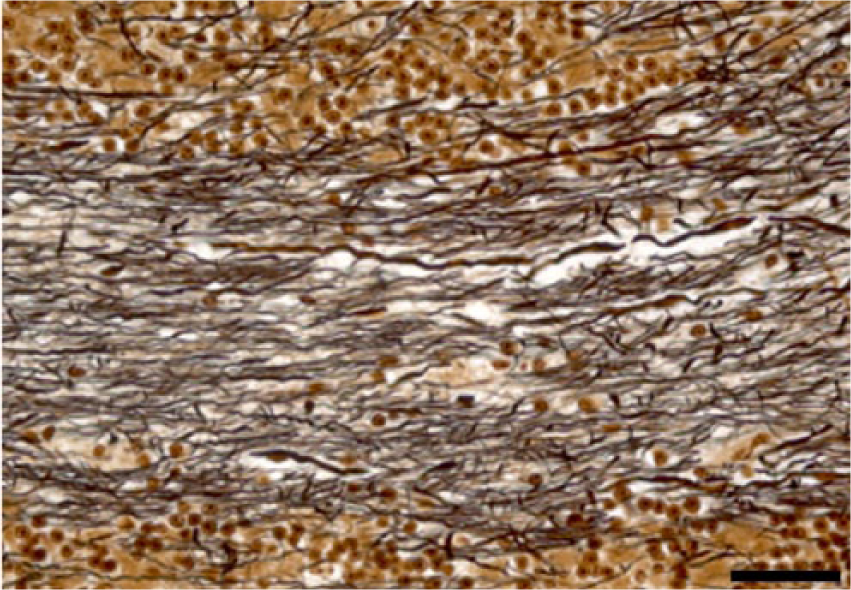
Cerebellum. Steer experimentally intoxicated with Solanum bonariense. Reduction in the axonal density (axonal loss) in the white matter of cerebellar folia. Bielschowsky stain. Bar = 40 µm.
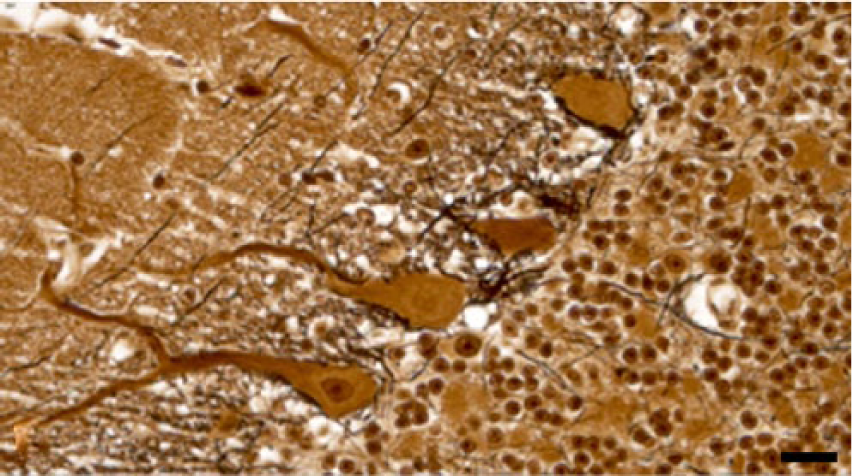
Cerebellar cortex; control. In a normal steer, Purkinje cells are surrounded by basket cell processes that stain black with Bielschowsky stain. Bar = 20 µm.
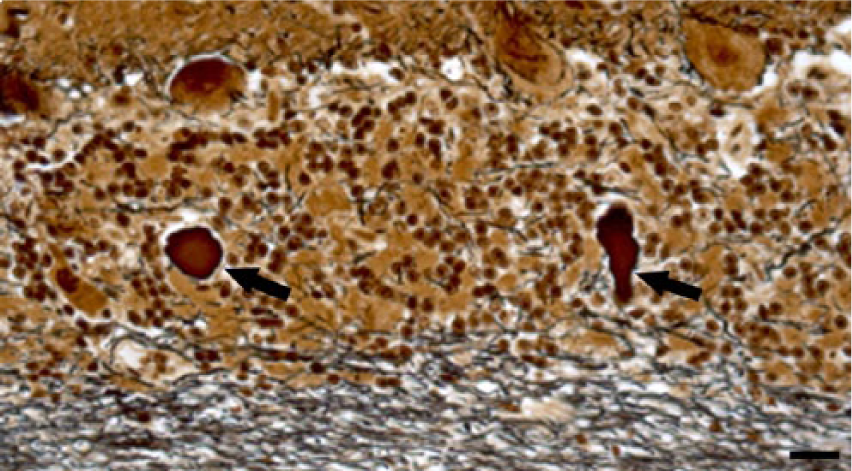
Cerebellar cortex. Steer experimentally intoxicated with Solanum bonariense. Degenerate Purkinje cells loss with reduced staining of adjacent basket cell processes and 2 axonal torpedoes in the granular layer (arrows). Bielschowsky stain. Bar = 20 µm.
Immunohistochemical staining
NF-200KDa and GFAP
Simultaneous cerebellar neuronal loss and reactive astrogliosis were evaluated comparing consecutive tissue sections immunolabeled with the neuronal marker NF-200KDa and anti-GFAP antibody. In cases with more severe lesions, marked reactive astrogliosis was present diffusely in the cerebellar cortex and white matter, as demonstrated with anti-GFAP. The immunoreactivity was particularly prominent in the Purkinje cell layer (Bergmann astroglia), with an enhanced expression of their radial processes in the molecular layer; and to a lesser extent in reactive astrocytes in the granular layer (Figs. 5, 6). In the more severely affected animals, cerebellar folia with low or no immunoreactivity against NF-200KDa in the Purkinje cell layer (neuronal loss) correlated with higher GFAP expression in the corresponding overlying molecular layer (Figs. 7, 8), demonstrating concomitant Purkinje cell loss and astrogliosis.
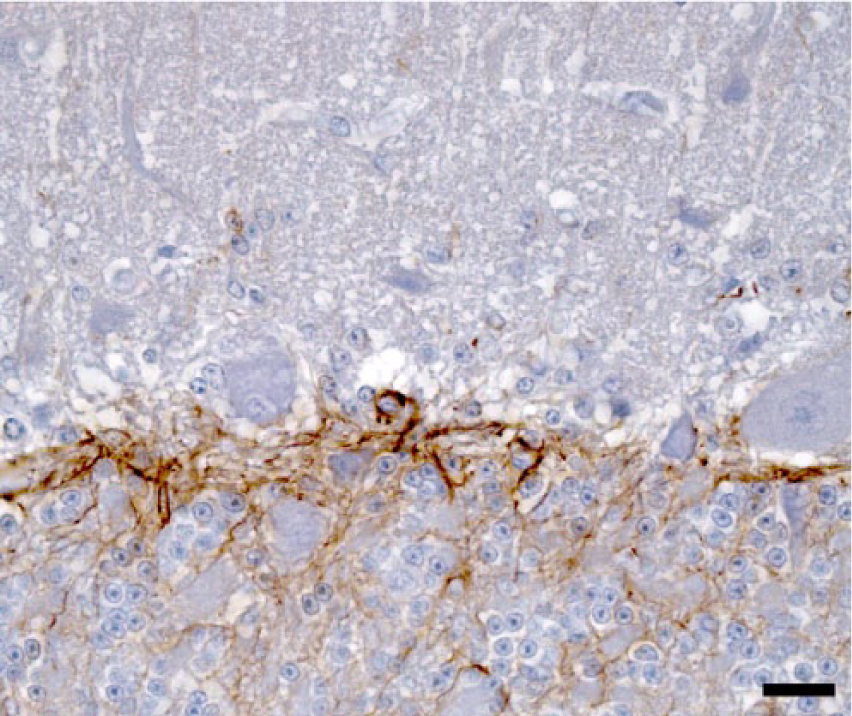
Cerebellar cortex; control. Immunoreaction against glial fibrillary acidic protein (GFAP) in the Purkinje and granular cell layers of a normal steer. Note the lack of immunoreactivity in the molecular layer (top). Anti-GFAP immunohistochemistry and hematoxylin counterstain. Bar = 20 µm.
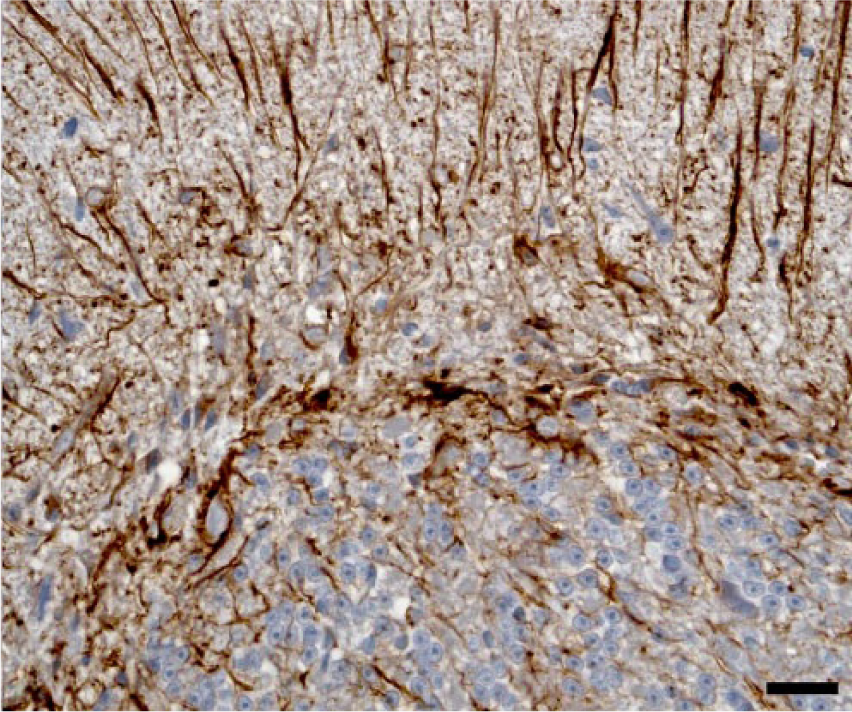
Cerebellar cortex. Steer experimentally intoxicated with Solanum bonariense. Intense immunoreaction of Bergmann astroglia and their processes in the molecular layer (astrogliosis) and mild astrogliosis in the granular layer. Anti–glial fibrillary acidic protein immunohistochemistry and hematoxylin counterstain. Bar = 20 µm.
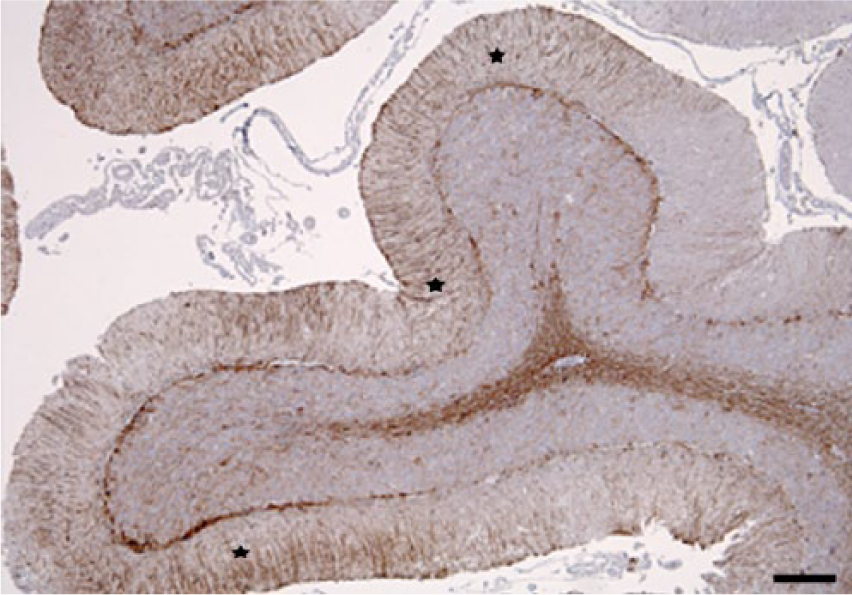
Cerebellum. Steer experimentally intoxicated with Solanum bonariense. Areas with intense glial fibrillary acidic protein (GFAP) immunoreaction indicating severe reactive astrogliosis (stars) in molecular layer coincide with areas of Purkinje cells loss (see Fig. 8). Anti-GFAP immunohistochemistry and hematoxylin counterstain. Bar = 200 µm.
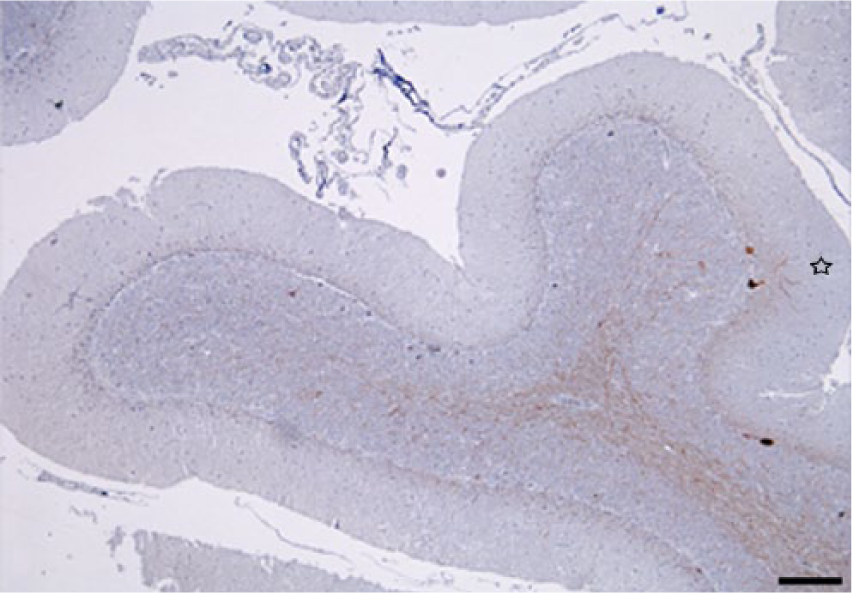
Cerebellum. Steer experimentally intoxicated with Solanum bonariense. Reduction in the number of Purkinje cell bodies and dendritic processes (empty star) in molecular layer, suggesting that remaining Purkinje cells limit astroglial reaction of the same cortical area (see Fig. 7). Anti–neurofilament (NF)-200KDa immunohistochemistry and hematoxylin counterstain. Bar = 200 µm.
Phospho-NF protein
In control animals, phospho-NF protein immunohistochemical staining showed a fluorescent signal distributed homogeneously throughout the cytoplasm of Purkinje cells, with a clear delineation of the central nuclear area. Immunoreactivity for phospho-NF protein was also concentrated in basket cell processes in the Purkinje cell layer and axonal processes in the lower molecular layer confirming that this protein was highly expressed in the most distant peripheral processes of neurons, without a significant increase in the neuronal perikarya. In both spontaneous and experimental cases, degenerate Purkinje cells showed a displaced nuclear area surrounded by immunoreactive cytoplasm, and a nonimmunoreactive dark area toward the axonal pole of the cell body (presumably intracytoplasmic vesicles [organelle accumulation area]; Figs. 9, 10).
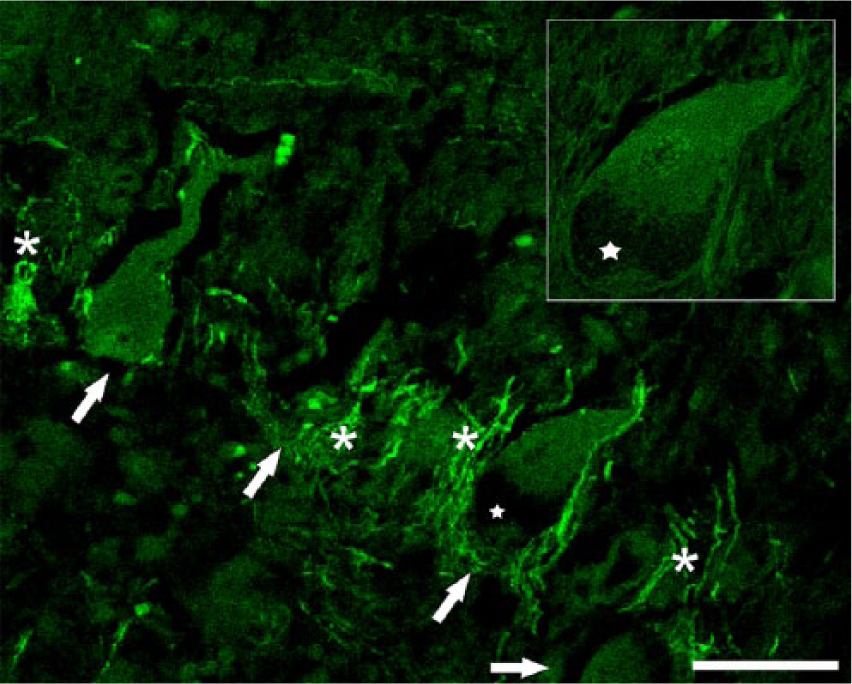
Cerebellar cortex. Cow naturally intoxicated with Solanum bonariense. Purkinje neuron perikarya (arrows) show different degrees of degeneration. The perikaryon of one of the Purkinje cells (star and inset) has a typical nuclear displacement and a peripheral region without labeling consistent with the organelle storage area. Neighboring basket cell axonal process show intense labeling around Purkinje cell perikarya (asterisks). Fluorescent immunohistochemistry for phospho-neurofilament protein (clone SMI-31). Bar = 50 µm.
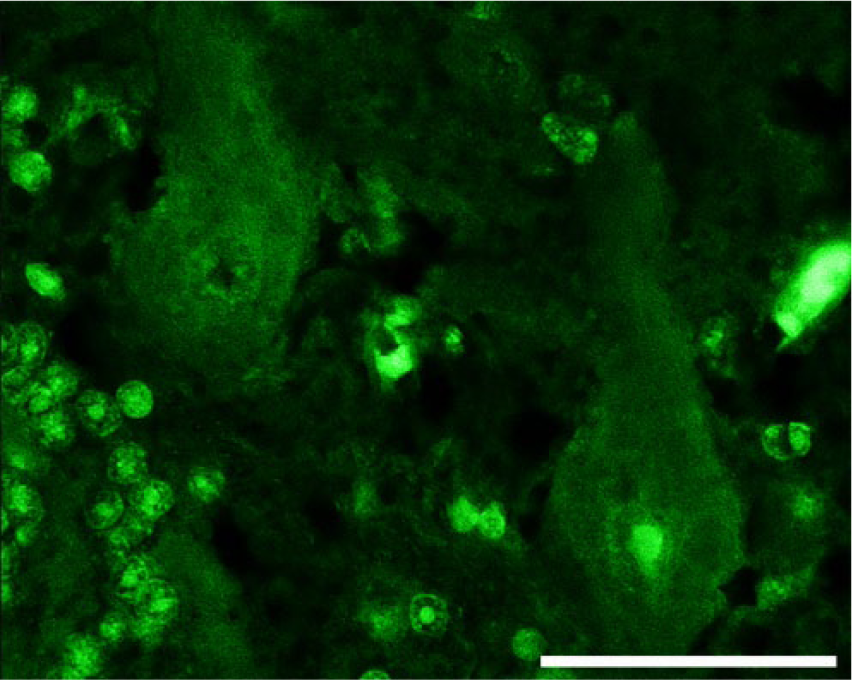
Cerebellar cortex; control. Purkinje cells from a normal cow show diffuse homogeneous immunofluorescent signal throughout the perikarya. Fluorescent immunohistochemistry for phospho-neurofilament protein (clone SMI-31). Bar = 50 µm.
β-tubulin
Immunohistochemical staining for β-tubulin produced in all cases an intense fluorescent signal within the perikarya of Purkinje cells and cerebellar granular neurons, and their neuronal processes. This signal allowed ready identification of specific organelles (such as nuclei), different neuronal types and layers in the cerebellar cortex of both intoxicated and control animals (Figs. 11, 12). Furthermore, this marker allowed the identification of specific intracellular changes in degenerate Purkinje cells of affected cattle, which showed a displacement of fluorescent immunoreaction toward the dendritic pole of the neuronal perikaryon, surrounded by a peripheral dark nonimmunolabeled area (Fig. 12).
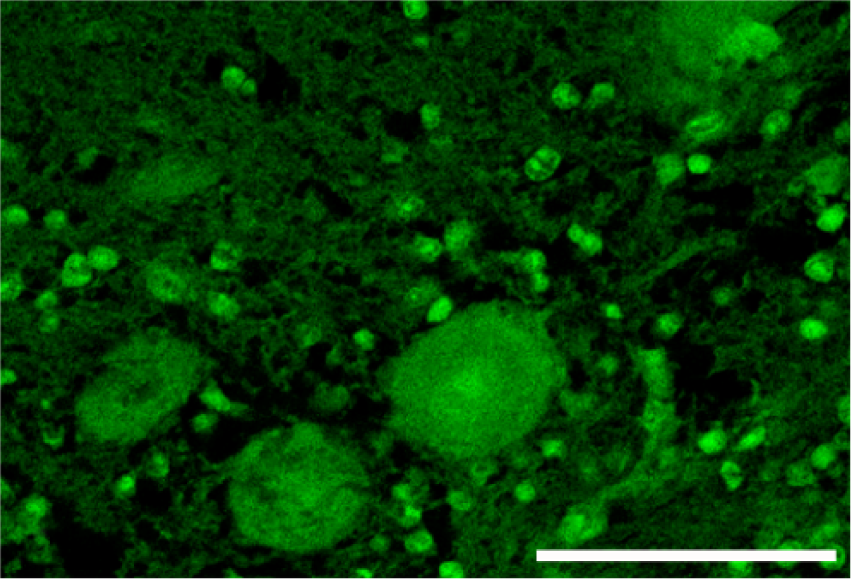
Cerebellar cortex; control. Normal distribution of β-tubulin in Purkinje cells and granular neurons of a normal cow. Fluorescent immunohistochemistry for β-tubulin. Bar = 50 μm.
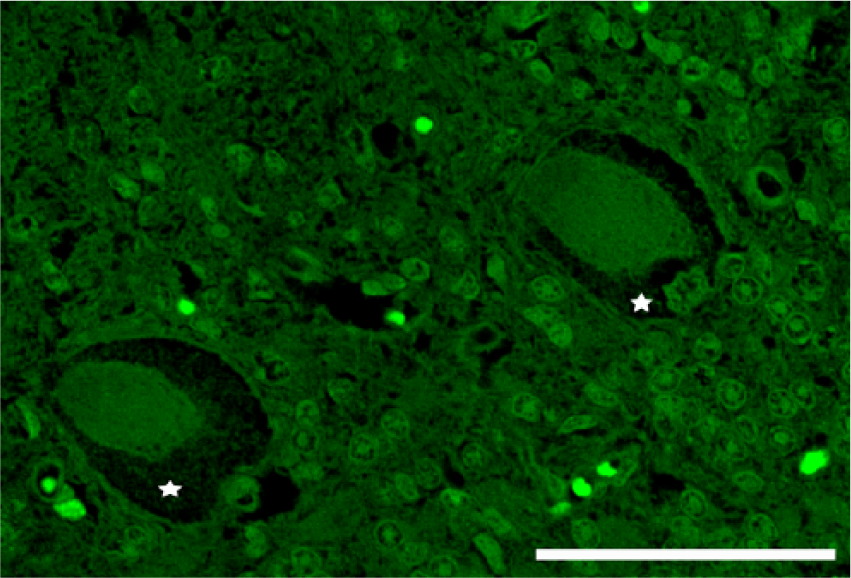
Cerebellar cortex. Cow naturally intoxicated with Solanum bonariense. The perikarya of 2 adjacent Purkinje neurons show peripheral nonlabeled regions (organelle accumulation area; stars) indicating lack of β-tubulin. Fluorescent immunohistochemistry for β-tubulin. Bar = 50 μm.
Phalloidin
Affinity reaction to actin microfilaments using the phalloidin probe showed a wide and diffuse fluorescence distribution in the perikarya of Purkinje cells and other neurons of the cerebellar cortex in unaffected control animals. In the case of degenerate Purkinje cells in affected animals, those specific areas that had been previously identified as intracytoplasmic organelle accumulation areas using the phase contrast lens, presented peripheral displacement of the nucleus and the perikaryal fluorescent signal. These areas were surrounded by marginal dark regions, also noted with phase contrast in the same neurons.
Discussion
Solanum bonariense intoxication in cattle causes pronounced microvesicular perikaryal vacuolation in cerebellar Purkinje cells.20,30 This same pathologic change has been described in intoxications with other plant species in the Solanum genus (S. fastigiatum [now considered S. bonariense]),1,26,27,30,32 S. paniculatum,1,6 S. kwebense,24,29 and S. subinerme. 14 Transmission electron microscopic examination of degenerate Purkinje cells revealed significant ultrastructural abnormalities in most of the perikaryal organelles and axonal processes, with accumulation of electron-dense residual storage bodies in membrane-bound vesicles, which eventually leads to lethal neuronal damage. 30
The pathogenesis of the cerebellar degeneration induced by S. bonariense has been compared to that of acquired neuronal lysosomal storage diseases such as those produced by species of Astragalus, Oxytropis, Swainsona, and Sida 27 ; however, the mechanisms leading to Purkinje cell degeneration and death are largely unknown. In an attempt to better characterize the lesions in this neurodegenerative disease and establish the bases to further understand its pathogenesis, we used histochemical and immunohistochemical methods to assess: 1) alteration in the distribution of cytoskeletal proteins in the perikarya of Purkinje cells; 2) cerebellar demyelination; 3) Purkinje cell loss; 4) the integrity of the basket cells in the cerebellar cortex; and 5) reactive astrogliosis. In the following paragraphs, we briefly discuss each of these categories.
Cytoskeletal proteins in the perikarya of Purkinje cells
The neuronal cytoskeleton plays a central role in the maintenance of neuronal shape, and provides structural organization for the intracellular media to establish metabolic compartments and intracellular transport. 25 Axonal transport systems allow the movement of molecules and cellular organelles from the cell body through the axon to the synapses (fast and slow anterograde transport), and the degradation products back to the cell body (fast retrograde transport). Fast anterograde axonal transport moves materials not intended for use in the neuronal body; these materials formed from the Golgi apparatus are principally membrane-bound vesicles containing neurotransmitters and enzymes. 36 These materials (as well as mitochondria, etc.) are moved down the axon on microtubules, and alterations in this transport system can lead to severe neuronal dysfunction and perikaryal accumulation of membrane-bound vesicles. 36 Cytoskeletal proteins normally have a characteristic immunohistochemical distribution within normal neurons, 4 which is altered in several neuropathies (such as human and animal motor neuron disease, hereditary canine spinal muscular atrophy, shaker calf syndrome, swayback, and cerebellar degeneration induced by diphenyl ditelluride in preweaning rats) characterized by a disrupted or aberrant cytoskeleton.3,4,7,8,15–17,25 Neurofilaments are the most abundant proteins in the nervous system and high MW NF proteins (NF-H) are among the most highly phosphorylated proteins in the brain. In normal neurons, NFs are phosphorylated in the axonal compartment but, in human neurodegenerative diseases, NF proteins are aberrantly hyperphosphorylated in neuronal cell bodies, and these perikaryal NF aggregations are pathological hallmarks of many of these neurodegenerative disorders. An increase in NF phosphorylation in the neuronal cell body results in blockage of axonal transport and neuronal cell death.3,4,7,8,15–17,25 NF proteins are synthesized in the neuronal body and are assembled and transported into axons, where they primarily provide structural support for the axon. The main recognized role of NFs is to increase the caliber of myelinated axons and consequently their conduction velocity.19,23
In a previous report, a marked reduction in the number of microtubules and NFs in Purkinje cell perikarya was demonstrated by transmission electron microscopy in S. fastigiatum intoxication in cattle, 1 and similar ultrastructural alterations were seen in S. bonariense intoxication in this species. 30 Our current results provide immunohistochemical evidence of a progressive loss of NFs in Purkinje cells in the cerebellar cortex, with an altered distribution of the other evaluated cytoskeletal components (β-tubulin, actin). Interestingly, this disruption of the cytoskeletal homeostasis was not characterized by abnormal accumulation of phospho-NF proteins or microtubules in neuronal perikarya, as seen in other neurodegenerative diseases.3,4,7,8,15–17,25 In contrast, the cytoskeletal disruption in S. bonariense–intoxicated cattle in our series was characterized by abnormal distribution of cytoskeletal components in the cell soma, suggesting that there might be a mechanic displacement of cytoskeletal proteins and/or the activation of different and perhaps unique metabolic pathways in S. bonariense intoxication. A previous study suggested that the progressive Purkinje cell death induced by S. bonariense is not mediated by excitotoxicity or apoptotic activation. 31 In another report, we suggested that mitochondrial damage might alter protein synthesis and lead to cytoskeletal disruption altering the axonal transport, playing a potential role in the pathogenesis of S. bonariense intoxication. 30 This hypothesis agrees with the concept that defects in the synthesis and transport of cytoskeletal components may provoke neuronal degenerative changes,13,15,34 but in our cases, we do not determine a primary or a “downstream” origin. Nervous tissue appears to be particularly sensitive to perturbations in the protein synthesis, perhaps because central nervous system (CNS) neurons have a very demanding metabolism, and shortcomings in translation that are tolerated in other cell types become limiting in this tissue. 12 Purkinje cells have an exceptionally high metabolic demand, and for that reason are particularly vulnerable to metabolic injury.11,33
Cerebellar demyelination and Purkinje cell loss
Our results further confirm that cerebellar demyelination is a feature of S. bonariense intoxication and that this lesion is more severe in animals with more severe clinical disease (positive clinicopathologic correlation). Furthermore, demyelination occurs with concurrent loss of Purkinje cells and their axonal processes. These findings suggest that cerebellar demyelination induced by S. bonariense is most likely secondary to neuronal damage (rather than being a primary event), as has been suggested in other neurodegenerative diseases, including in this group several infectious, metabolic, and toxic diseases of humans and domestic animals.18,28
Integrity of basket cells in the cerebellar cortex
Bielschowsky stain showed a positive correlation between degree and extent of Purkinje cell degeneration and loss and basket cell loss and a concomitant proliferation of Bergmann’s astrocytes (see next section) in the cerebellar cortex. This loss of both basket and Purkinje cells is probably the reason for the decreased thickness of the cerebellar molecular layer reported in cattle intoxicated with S. fastigiatum, 26 and also suggests that basket cell depletion likely occurs secondarily to Purkinje cell degeneration and death. The demonstration of empty baskets corresponding to sites of Purkinje cell loss has been used as a confirmatory evidence of cerebellar cortical degeneration in a variety of diseases in dogs,2,8,10,21,35 and also in cattle intoxicated with S. fastigiatum. 27 As expected, in our series, Purkinje cell loss was more pronounced in animals with more severe clinical signs, suggesting that degeneration and loss of Purkinje cells is directly associated with the development and progression of clinical signs. The most likely pathophysiologic mechanism involved is the lack of inhibition of neurons of the cerebellar nuclei by dysfunctional Purkinje cells, 11 which is likely responsible for the abnormal rate, range, and force of voluntary movements, as well as the loss of balance, menace deficit, and nystagmus observed in affected animals. 30
Reactive astrogliosis
The loss of Purkinje cells is typically attended by hypertrophy and hyperplasia of Bergmann glia, whose prominent processes extend radially upward through the thinned molecular layer to the pial surface. Our results demonstrate that Purkinje cell loss is accompanied by an increase of GFAP-immunoreactive astrocytes in the same cerebellar cortical areas, showing an intense immunoreaction of Bergmann astroglia and their processes in the molecular layer and mild astrogliosis in the granular layer. This supports previous results reported in several neurodegenerative conditions, in which astrocytes react promptly after CNS injury such as AIDS dementia, a variety of other viral infections, prion-associated spongiform encephalopathies, inflammatory demyelinating diseases, acute traumatic brain injury, neurodegenerative diseases such as Alzheimer’s disease, and Solanum intoxication of cattle.5,27,37
In conclusion, based on the results of the present work, we propose that cytoskeletal disruption in Purkinje cells likely plays a role in the S. bonariense–induced cerebellar cortical degeneration in cattle. Additionally, we demonstrate that Purkinje cell loss is associated with diffuse astrocytosis and precedes loss of basket cells in the cerebellar gray matter. Concurrent demyelination and axonal loss, demonstrated in the cerebellar white matter, are presumed to be secondary to Purkinje cell death. The severity of cerebellar lesions is positively correlated with the severity of the clinical signs. Furthermore, the combined techniques used herein could become an interesting tool to improve the diagnosis of cerebellar degeneration induced by S. bonariense, and establishes data with which other bovine cerebellar cortical degenerations could be compared in the future. Although this study broadens our knowledge of the CNS lesions induced by S. bonariense in cattle, further investigations are needed in order to better understand the pathogenesis of this neurodegenerative disorder.
Footnotes
Acknowledgements
Our gratitude to Lic. Valentín Cóppola and Dr. Alejandra Kun (IIBCE, Uruguay), Dr. Alba Román (Universidad de Santiago de Compostela, Spain), Ester Blasco and Lola Pérez (Universitat Autònoma de Barcelona, Spain) for technical assistance, Dr. James Pfister (USDA, United States) and Dr. Claudio Borteiro (Universidad de la República, Uruguay) for manuscript proofreading, and especially to Emeritus Prof. Dr. Kosuke Okada (Iwate University, Japan) for his constructive criticism and review of this article.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by “Comisión Sectorial de Investigación Científica de la Universidad de la República” (CSIC, UdelaR), “Comisión de Investigación y Desarrollo Científico” (Veterinary School, UdelaR) and “Programa de Desarrollo de las Ciencias Básicas” (PEDECIBA), Uruguay; and PICT 2011/1379 (FONCyT), Argentina. JMV and AC are Research Career Members of the “Sistema Nacional de Investigadores” (SNI-ANII), Uruguay. EJG is a Research Career Member of the “Consejo Nacional de Investigaciones Científicas y Técnicas,” Argentina.
a.
Clone N-0142; Sigma-Aldrich, St. Louis, MO.
b.
SMI-31; Covance, Princeton, NJ.
c.
Clone 2-28-33; Sigma-Aldrich, St. Louis, MO.
d.
Invitrogen; Life Technologies, Grand Island, NY.
e.
Clone Z0334; Dako Denmark A/S, Glostrup, Denmark.
f.
EnVision; Dako Denmark A/S, Glostrup, Denmark.
g.
Fluorescein conjugated goat anti-mouse IgG; Sigma-Aldrich, St. Louis, MO.
h.
Invitrogen ProLong Gold antifade reagent; Life Technologies, Grand Island, NY.
i.
BX41 microscope; Olympus Corp., Tokyo, Japan.
j.
BX61 microscope with a confocal module Fluoview FV 300 equipped with He-Ne lasers emitting at 488 and 543 nm; Olympus Corp., Tokyo, Japan.