
Abstract
A significantly increased risk for a number of autoimmune and infectious diseases in purebred and mixed-breed dogs has been associated with certain alleles or allele combinations of the dog leukocyte antigen (DLA) class II complex containing the DRB1, DQA1, and DQB1 genes. The exact level of risk depends on the specific disease, the alleles in question, and whether alleles exist in a homozygous or heterozygous state. The gold standard for identifying high-risk alleles and their zygosity has involved direct sequencing of the exon 2 regions of each of the 3 genes. However, sequencing and identification of specific alleles at each of the 3 loci are relatively expensive and sequencing techniques are not ideal for additional parentage or identity determination. However, it is often possible to get the same information from sequencing only 1 gene given the small number of possible alleles at each locus in purebred dogs, extensive homozygosity, and tendency for disease-causing alleles at each of the 3 loci to be strongly linked to each other into haplotypes. Therefore, genetic testing in purebred dogs with immune diseases can be often simplified by sequencing alleles at 1 rather than 3 loci. Further simplification of genetic tests for canine immune diseases can be achieved by the use of alternative genetic markers in the DLA class II region that are also strongly linked with the disease genotype. These markers consist of either simple tandem repeats or single nucleotide polymorphisms that are also in strong linkage with specific DLA class II genotypes and/or haplotypes. The current study uses necrotizing meningoencephalitis of Pug dogs as a paradigm to assess simple alternative genetic tests for disease risk. It was possible to attain identical necrotizing meningoencephalitis risk assessments to 3-locus DLA class II sequencing by sequencing only the DQB1 gene, using 3 DLA class II–linked simple tandem repeat markers, or with a small single nucleotide polymorphism array designed to identify breed-specific DQB1 alleles.
Keywords
Introduction
Specific dog leukocyte antigen (DLA) class II genotypes and haplotypes have been associated with an increasing number of autoimmune disorders of purebred dogs. A Vogt–Koyanagi–Harada-like syndrome of Akita dogs has a strong association with certain alleles and allele combinations of DLA class II genes, 1 as does autoimmune hemolytic anemia, 20 immune arthritis, 28 hypothyroidism in several breeds, 25,26,38 type I diabetes mellitus in Samoyed dogs and Cairn and Tibetan Terriers, 23 anal furunculosis of German Shepherd Dogs, 5 and systemic lupus erythematosus–related complex of Nova Scotia Tolling Retrievers. 37 Necrotizing meningoencephalitis (NME) of Pug dogs 10 and Addison disease in Nova Scotia Duck Tolling Retrievers 12 have been recently added to this growing list. Dog leukocyte antigen class II associations have also been described for visceral leishmaniasis 30 in domestic dogs in Spain and for generalized demodicosis in Boxer, Argentinean Mastiff, and mixed-breed dogs in Argentina. 16 It appears, therefore, that immune disorders of dogs follow the pattern observed in humans (i.e., that most will occur in context of certain alleles within genes of the major histocompatibility complex [MHC]). 34 Unlike human beings, the incidence of immune disorders in dogs has increased with purebreeding, 29 which often involves inbreeding on both maternal and paternal sides. 3,36
Reports on the risk association of certain DLA class II types and autoimmune disorders and chronic infectious diseases in dogs have frequently mentioned the potential value of direct sequencing for allele assignment of the 3 DLA class II genes: DRB1, DQA1, and DQB1. However, direct sequencing of alleles at 3 different loci is time consuming, is expensive, and cannot easily be combined with genetic tests for parentage and/or identification. Therefore, the objective of the current study was to evaluate simpler and more versatile alternative test procedures to 3-locus DLA class II gene sequencing that might be applicable to immune disorders in dogs that are known to have DLA class II gene–associated disease risk. The tests should be applicable to buccal swab DNA and, if possible, easily combined with parentage and/or identification markers. The disease chosen for the current study was NME of Pugs, which, like a spectrum of immune disorders of other purebreds, takes advantage of limited DLA diversity, strong DLA class II allele linkages, and a high degree of genetic homozygosity.

Nucleotide alignment of exon 2 of the 6 dog leukocyte antigen DQB1 alleles detected in Pug dogs. The nucleotides in bold are locations of single-nucleotide polymorphisms identified by Sequenom MassArray Design software e that can be used to identify each of the 6 alleles.
Although many immune diseases of dogs in a number of purebreds have similar DLA class II gene–associated risks, NME of Pugs was chosen in the current study to test the feasibility of simplifying genetic risk assessment. Necrotizing meningoencephalitis in Pugs, formerly known as Pug dog encephalitis, is a progressive and fatal disease affecting 1–2% of Pugs, usually before 7 years of age. 9,27 There has been no laboratory evidence for an infectious cause, 27,32,35 and the etiology is now presumed to be immunologic. 9,35 Pug dog NME has been likened to fulminate atypical forms of human multiple sclerosis in its form and strong association to a specific human leukocyte antigen class II genotype. 10 A whole genome scan recently associated risk for the disease solely with a specific region on canine chromosome 12 coding for the genes of the DLA complex. 10 Fine mapping studies revealed a region of high homozygosity that included the DLA class II DRB1, DQA1, and DQB1 genes. 10 Sequencing of these 3 genes confirmed both a high degree of homozygosity and a strong linkage between certain alleles at each loci, yielding a high-risk haplotype DLA-DRB1*010011/DQA1*00201/DQB1*01501. Risk-protective and neutral allele combinations were also identified. These strong linkages also allow for several simple alternative approaches to direct sequences of alleles at all 3 DLA class II gene loci, including sequencing a single DLA class II locus (DQB1 in Pugs), the use of a threesome of DLA class II–linked simple tandem repeat (STR) markers, and a 4 single-nucleotide polymorphism (SNP)–based array capable of identifying breed-specific DLA class II alleles (DQB1 in Pugs).
Materials and methods
Case material
Purified DNA or DNA-containing tissue specimens were obtained from NME-affected and healthy Pugs. Some samples, in the form of liver or blood, were from Pugs previously reported. 10 Additional samples in the form of blood were from the collections of Dr. Karen Vernau, School of Veterinary Medicine, University of California, Davis. A final group of samples in the form of buccal swabs was also obtained from healthy Pugs sampled as part of a large collection of purebred dogs from the Veterinary Genetics Laboratory, University of California, Davis. The exact numbers of animals tested for each procedure are listed in the results and appropriate tables; exact numbers of samples and origins varied for each test procedure, depending on sample availability at the time. DNA was isolated from whole blood and liver samples using a standard proteinase K digestion procedure. 31 DNA isolation from the buccal swabs was performed as previously described. 1,15
DLA-DRB1/DQA1/DQB1 allele sequencing for NME risk
Exon 2 of DLA class II genes DRB1, DQA1, and DQB1 was sequenced using internationally recognized intronic primers according to a standardized protocol. 17,22,24 The 270–base pair polymorphic sequence in exon 2 of the DLA-DRB1 was amplified using primers DRBF (5′-GAT CCCCCCGTCCCCACAG-3′) and DRBR (5′-TGTGTCA CACACCTCAGCACCA-3′). The 307–base pair polymorphic sequence in exon 2 of DLA-DQA1 was amplified using primers DQAF (5′-TAAGGTTCTTTTCTCCCTCT-3′) and DQAR (5′-GGACAGATTCAGTGAAGAGA-3′). The 267–base pair polymorphic sequence in exon 2 of the DLA-DQB1 was amplified using primers DQBF (5′-TCACT GGCCCGGCTGTCTCC-3′) and DQBR (5′-GGTGC GCTCACCTCGCCGCT-3′). Polymerase chain reaction (PCR) was carried out in 20-μl reaction volumes containing 20–50 ng of DNA, 0.2 mM of deoxyribonucleotide triphosphate, 0.25 μM of forward and reverse primers in 1× PCR buffer, and 1 unit of DNA polymerase. a The mixture was then incubated for 5 min at 93°C followed by 30 cycles of 45 sec at 93°C, 45 sec at 61°C for DRB1, 57°C for DQA1, and 66°C for DQB1; for 1 min at 68°C; and then a final extension of 10 min at 68°C. The PCR products were electrophoresed on 2% agarose gel and purified with a gel purification kit a according to the manufacturer's instructions.
Sequencing of exon 2 regions of the DLA-DRB, -DQA, and -DQB genes was performed in both directions for the NME-affected Pugs and in one direction for healthy control dogs. For one-direction sequencing, the reverse primers for DRB1 and DQB1 and forward primer for DQA1 were used. The purified PCR products were sequenced with a cycle sequencing kit b in a 15-μl reaction containing 1 μl of terminator mix, b 2.2 μl of reaction buffer (5×), 35 ng of sequencing primer, and 3 μl of purified PCR product. The sequencing reaction was incubated at 93°C for 2 min and amplified for 40 cycles at 93°C for 20 sec, 50°C for 20 sec, and 60°C for 4 min. Unincorporated dye terminators and deoxyribonucleotide triphosphates were removed by gel filtration, c and the amplified products were analyzed by capillary electrophoresis on a genetic analyzer. Sequence analysis software d was used for alignment of sequence data. Homozygous and heterozygous (mixed) sequences were resolved, and allele nomenclature was assigned according to an international standard. 18 All DLA-DRB1, -DQA1, and -DQB1 alleles identified among Pugs10 were previously known from this and other breeds.
Polymerase chain reaction (PCR) and extension primer sequences used for single-nucleotide polymorphism (SNP) testing e for specific Pug dog leukocyte antigen DQB1 alleles.
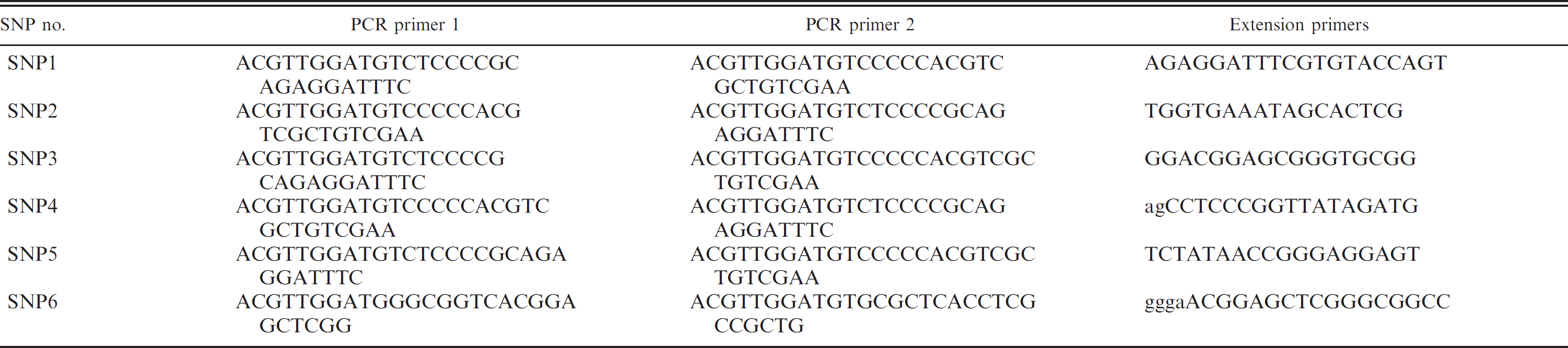
Dog leukocyte antigen class II alleles and their haplotype frequency in 51 necrotizing meningoencephalitis–affected and 57 healthy control Pug dogs.
Proportion of haplotypes in affected dogs: (102/216) = 47.22%. Binomial test used to assess probability.
Linked STR marker test for NME risk
Three STRs (12-005A_CA, 12_005A_CT, 12_005B_CA) on Canis familiaris autosome (CFA12) were mined from the canine genome (nucleotide 5,018,175; 5,361,083; and 5,677,936). These 3 markers were closely linked to the DLA class II region and certain alleles strongly associated with risk for developing NME in Pugs. 10 The original primer sequences used for the present study were derived from a set of 3,000 STR markers 40 and listed in DOGSET 39 (http://www.vgl.ucdavis.edu/dogset). The primer sequences and fluorescent tags used in the present study were as follows: 5ACA-F (FAM) TATTGCACCCTGGTGTCT GC, 5ACA-R TTGCCCTGGGTGGTAAAATC; 5ACT-F (NED) GGAACCCCCTGTAAAATTTCTT, 5ACT-R CAGCCAAGACCTTAGGAGCAA; 5BCA-F CCCTGG ATATGTGGCAGTCA, 5BCA-R (NED) TGCCCTCTT CCACTTCACCT. The PCR-amplified products were analyzed using a capillary instrument. b
Single-nucleotide polymorphism testing for DQB1 genotype
Six SNPs based on the sequences of 6 known Pug DLA-DQB1 alleles were identified using assay design software e (Fig. 1). The PCR and extension primer sequences of the 6 SNPs are given in Table 1. The SNPs were assayed using an automated system. e Although all 6 SNPs were used to test each sample, SNPs 1, 2, 5, and 6; 1, 2, 4, and 6; 2, 3, 4, and 6; and 2, 3, 5, and 6 were equally useful in allele identification. Results from SNPS 1, 2, 5, and 6 were used primarily for this study. The additional SNPs were used to confirm the genotype and to allow for occasional allele dropout in the primary SNP panel.
Results
Three-locus DLA class II haplotypes and relationship to NME risk
Exon 2 of DLA-DRB1, -DQA1, and -DQB1 genes were sequenced from a total of 51 NME-affected and 57 healthy Pugs. Results from 35 normal and 38 NME-affected Pugs from this group were previously reported, 10 and 13 new affected and 22 new normal animals were added for this study. Five DRB1 (010011, 01501, 01502, 00601, and 01702), 3 DQA1 (00201, 00601, and 005011), and 6 DQB1 (01501, 02301, 02601, 00701, 01304, and 05701) alleles were identified (Table 2). These findings were identical to those previously reported, 10 with the exception of DQB1*05701, which was not observed among the original group of Pugs. This allele differs from DQB1*02301 by 2 nucleotides, each of which causes a change in amino acid (Fig. 1). It has been observed previously in a small number of Pugs and Husky dogs and has now been recognized as a new allele (L. J. Kennedy, University of Manchester, personal communication, 2010). The DQB1*05701 allele was interchangeable with DQB1*02301 in the DRB1*01501/DQA1*00601/DQB1*02301 haplotype.
The 5 DRB1, 3 DQA1, and 6 DQB1 alleles formed 7 3-locus haplotypes, the frequency of which is given in Table 2. These 7 haplotypes conferred high, neutral, or decreased risk for the development of NME, depending on zygosity (Tables 2, 3). A significant risk for disease was apparent when the predominant 010011/00201/01501 haplotype was present in a homozygous (odds ratio [OR] 15.58, relative risk [RR] 5.9) but not heterozygous state (OR 0.7, RR 0.78; Table 3). The 01502/00601/02301 haplotype was highly protective in both homozygous (OR 0, RR 0) and heterozygous (OR 0.14, RR 0.2) forms. The 01501/00601/02301 haplotype was protective in heterozygotes (OR 0.13, RR 0.16). The 01501/00601/02601 haplotype was protective in homozygotes (OR 0, RR 0) but had no effect in heterozygotes (OR 0.83, RR 0.85). The 00601/005011/00701 haplotype had no effect on disease in heterozygotes (OR 1.13, RR 1.1).
Odds ratio (OR) and relative risk (RR) for affected and normal Pug dogs that were either homozygous (homo) or heterozygous (hetero) for the alleles across DRB1/DQA1/DQB1 haplotype.
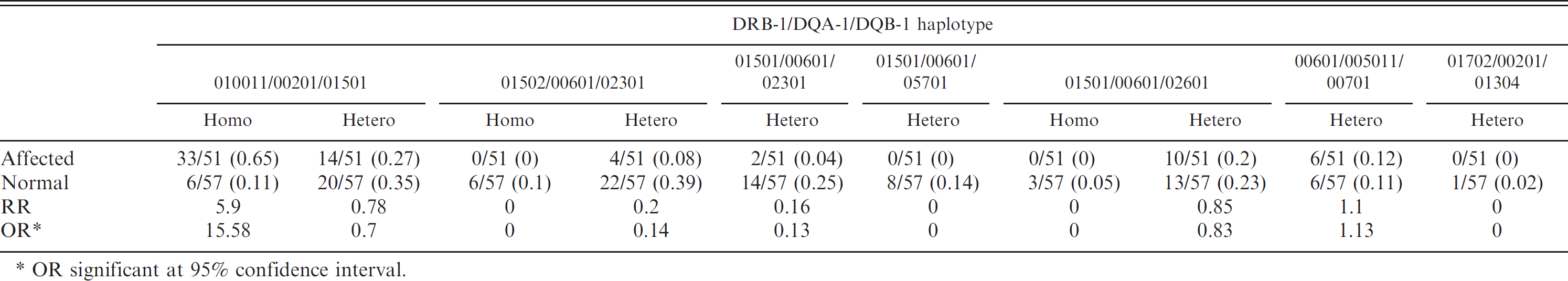
OR significant at 95% confidence interval.
Single-locus DLA class II genotype and relationship to NME risk
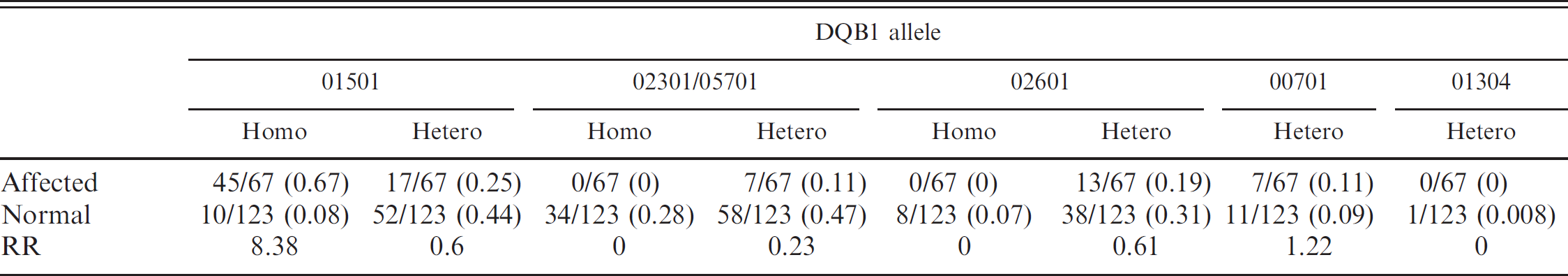
Fifty-one affected and 57 healthy Pugs were tested in the current study. Linkage between DRB1/DQA1/DQB1 alleles was such that NME risk could be defined solely by specific DQB1 alleles and their zygosity (Table 3). DQB1*01501 was associated with the 010011/00201/01501 haplotype, DQB1*02301 with the 01501/00601/02301 or 01502/00601/02301 haplotypes, DQB1*05701 with the 01501/00601/05701 haplotype, DQB1*02601 with the 01501/00601/02601 haplotype, DQB1*00701 with the 00601/005011/00701 haplotype, and DQB1*01304 with the 01702/00201/01304 haplotype. DQB1*01501 was also associated with high risk when in a homozygous state and neutral when heterozygous, whereas DQB1*02301 and *05701 were protective in both heterozygous and homozygous states. DQB*02601 was protective when homozygous and neutral in a heterozygous state; DQB*00701 was neutral in a heterozygous state, whereas DQB*01304 was too infrequent to assess risk. To validate the concept that the risk for NME can be determined by the status of DQB1 alone, the data set of Table 3 was expanded by sequencing only the DQB1 genes from an additional 16 affected and 66 healthy appearing Pugs (Table 4). These data indicate that specific DQB1 alleles and their zygosity had the same predictive risk value as the DRB1/DQA1/DQB1 haplotypes that they defined (compare Tables 3 and 4).
Correlation between high-risk STR testing and NME risk
Pugs from among the total sample collection were tested using 3 STR markers specific for the DLA class II region; 54 had NME and 229 were healthy at the time of sampling. Thirty-eight to 39 of the 54 NME-affected Pugs were homozygous for the predominant STR alleles 341, 327, and 280, but only 18 of the 229 healthy Pugs were homozygous for these same alleles (RR 6.6–9.0, OR 22.2–30.5; Table 5).
There was a strong, although not across-the-board, linkage between STR and DLA class II gene haplotypes (Table 6). The 341-327-280 STR haplotype defined the high-risk 3-locus DLA class II haplotype (010011/00201/01501), and Pugs homozygous for this STR haplotype were also homozygous for the high-risk 3-locus DLA class II haplotype and by inference for the high-risk DQB1*1501 genotype. The 01502/00601/02301 and 01501/00601/02301 haplotypes, which were protective against NME in both homozygous and heterozygous states, were not defined by a single STR haplotype. Rather, they were correlated with several different STR haplotypes, including 339/325/276, 343/324/284, and 343/324/282. The risk-neutral 00601/005011/00701 haplotype was defined by the 339/323/284 STR haplotype and was only found in a heterozygous state.
Relative risk (RR) for affected and normal Pug dogs that were either homozygous (homo) or heterozygous (hetero) for the alleles at DQB1 locus with expanded data.
Single-nucleotide polymorphism testing for DLA-DQB1 allele and NME risk
A 6-SNP panel, employing an automated system, e was used to identify DQB1 alleles in 99 healthy and 41 NME-affected Pugs from among the group that had been previously genotyped for DQB1 (Table 7). DQB1 allele assignments were based on SNPs 1, 2, 5, and 6, with SNPs 3 and 4 used for confirmation and backups for allele dropout. Five of the 6 known DQB1 alleles were automatically called by the software. The exception was DQB1*00701, which was automatically read as C at SNP6 and therefore erroneously designated DQB1*1304. Sequence similarities between DQB1 and DRB1 in this region resulted in a triple amplification of the C SNP and a single amplification of the G SNP, resulting in a large unanticipated C peak and a smaller anticipated G peak (Fig. 2B). This duplication was not evident with the other 5 DQB1 alleles because they all possessed C at SNP6 and were therefore called correctly on automatic read. There was concordance between DQB1*00701 sequence and SNP assignments when SNP6 peaks were manually examined and the C peak ignored (Table 7). Therefore, a simple SNP test e can be used to determine the risk for NME with the same accuracy as 1- or 3-locus DLA class II gene sequencing or a threesome of linked STR markers.
Correlation of homozygosity of the simple tandem repeat (STR) markers with necrotizing meningoencephalitis (NME) in Pug dogs. *
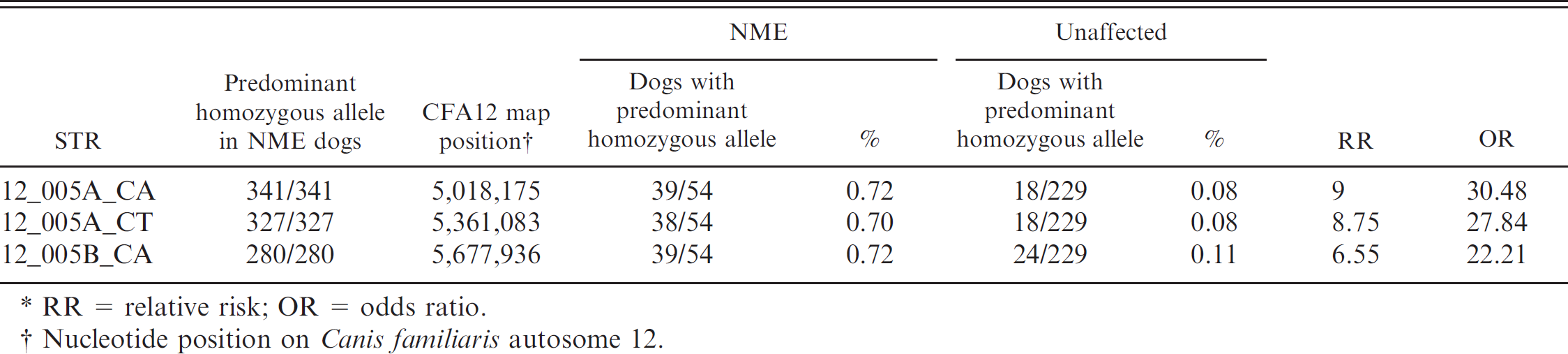
RR = relative risk; OR = odds ratio.
Nucleotide position on Canis familiaris autosome 12.
Discussion
Several genetic test approaches were used to assess DLA class II–associated disease risk using NME of Pugs as a model. The tests took advantage of limited genetic diversity and high-linkage disequilibrium in the DLA and a disease association with specific DLA class II alleles. Three-locus exon 2 sequencing provided exact identification of the alleles involved and their zygosity, allele haplotypes, and disease risk. However, because of strong allele linkages (haplotyping) between loci, sequencing of only the DQB1 alleles provided identical information on zygosity and disease risk as 3-locus sequencing. Given similar conditions in the DLA class II region among all purebreds, single DLA class II gene sequencing should also be applicable to other breeds with different DLA class II–associated immune disorders. The number of DLA class II alleles and haplotypes among dogs in general and purebreds in particular is very limited. 2,19,24 At least 143 DRB1, 26 DQA1, and 66 DQB1 alleles have been formally designated to date in all dogs, with a limited number still to be identified and/or named (L. J. Kennedy, personal communication, 2010). 17,18,21 Although the potential for allelic variation within the DLA class II region is great, this potential is limited by natural linkage disequilibrium in the DLA and by more recent inbreeding. Thus, DLA class II diversity among 6,434 purebred and mixed-breed dogs tested to date has been limited to 189 3-locus haplotypes (L. J. Kennedy, personal communication, 2010). Furthermore, each purebred dog possesses only a small number of specific 3-locus haplotypes, 2,19 similar to Pugs, with only 7 haplotypes identified to date.
Correlation of simple tandem repeat (STR) and DRB1/DQA1/DQB1 haplotypes and the relative risk (RR) and odds ratio (OR) for necrotizing meningoencephalitis in Pug dogs.
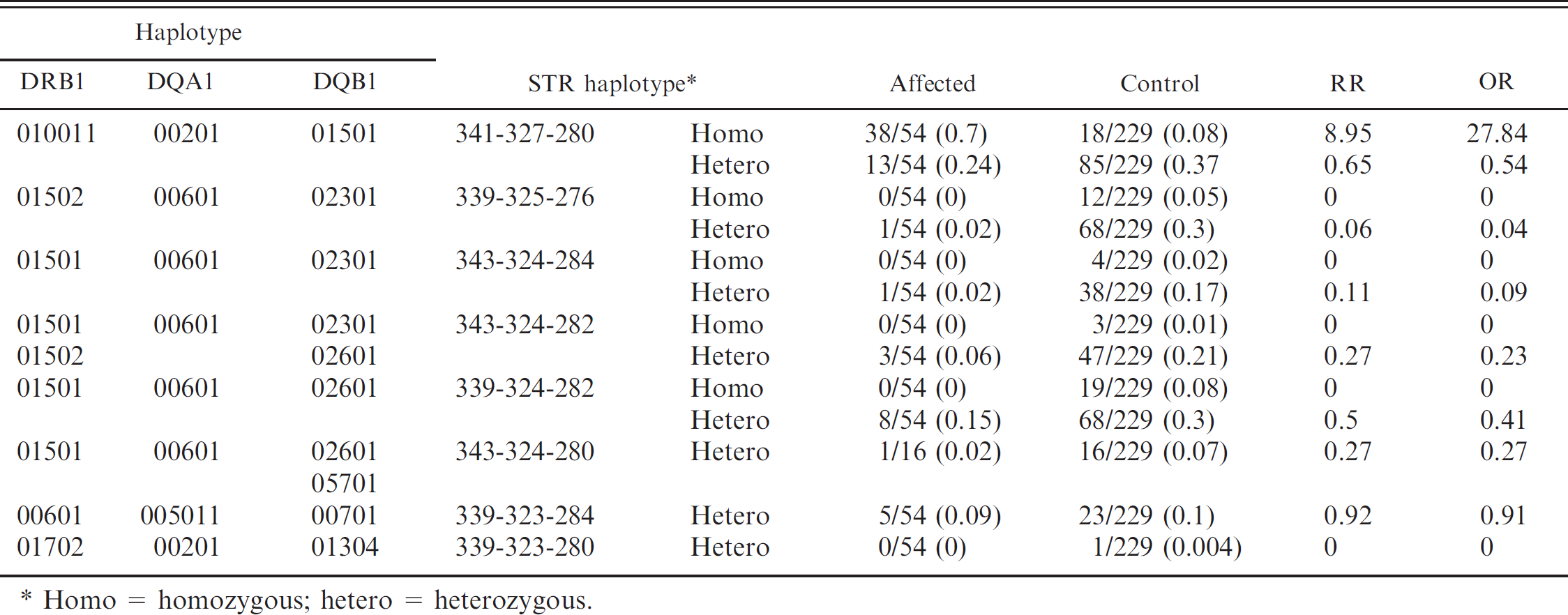
Homo = homozygous; hetero = heterozygous.
A linked STR marker test also worked well for determining DLA class II–associated risk for NME in Pugs. The linked STR marker test for Pug dog NME was quick, was relatively inexpensive, and could be easily incorporated into a larger panel including parentage and/or identification markers. 6–8 Nevertheless, association studies would have to be repeated with the present linked STR markers for other reported autoimmune and infectious diseases in dogs to ascertain their utility. The use of such a STR panel is not novel; a less DLA class II–specific linked STR marker test has proved effective in assessing risk for generalized demodicosis in Boxers, Argentinean Mastiffs, and mixed-breed dogs. 16 The STRs also undergo more genetic drift than coding genes (calculated at 1.1 × 10−2 for dogs 8 ), although such events are uncommon in the time periods in question and are easily recognized when parentage testing is included.
Single-nucleotide polymorphism testing e proved to be relatively simple in the Pug model and is applicable to DLA class II–associated immune disorders in other breeds. A small battery of SNPs was derived using the known exon 2 sequences of the 6 DLA-DQB1 alleles in Pugs. The small SNP panels used in the current study were able to predict the DQB1 genotypes and, by extension, the DRB1/DQA1/DQB1 haplotypes. The SNP-based testing may prove to be a simple and highly cost-effective method for determining DLA class II–associated genotypes and/or haplotypes in autoimmune and infectious disorders in purebreds. However, as would be the case for linked STR markers, each SNP panel would have to be customized for breed based on the disease and risk-associated DLA class II alleles. Care would also be required to read around occasional sequence similarities between DRB1 and DQB1 sequence homologies. The SNP testing, like STR assays, can also be easily incorporated with identification and parentage panels using a larger number (20–40) of SNPs identified for this purpose. 33
All of the assays described herein worked extremely well with relatively crude sodium hydroxide–extracted, buccal swab–derived DNA. This significantly reduces the cost of testing by eliminating the need for the collection of whole blood and more expensive extraction procedures. Buccal swabs are also easily stored in a dry state, and the DNA lasts for many years.
It is important that tests such as this, which measure disease risk, not be used for diagnosing a specific immune disease. This is because immune disorders often involve multiple genes as well as nongenetic factors. 11,13 Autoimmunity, as is now known from human genetics, involves a number of genes related to the immune system, both within and outside of the MHC region, which work in concert with certain MHC class I and II gene variations. 4,14,41 Epigenetic factors are also important in disease risk. This is best exemplified by comparing the disease phenotype and associated genotype in NME of Pugs. The incidence of NME in Pugs based on previous studies is approximately 1.2%, whereas the incidence of the high-risk haplotype in a homozygous state is 6–11%. 9 When these figures are used, only 1 in 5–10 Pugs at genetic risk for NME will develop the disease in their lifetime. Therefore, tests for susceptibility and protective genotypes and haplotypes described herein should be used only for assessing disease risk and not for diagnosing a specific disorder. Genetic testing can also be used to select breeding pairs that will produce puppies at low or negligible risk. The first inclination is to reduce risk by breeding only dogs possessing protective haplotypes. Elimination of the high-risk haplotype and increasing the incidence of protective haplotypes would lead to a significant loss of genetic diversity. The susceptibility haplotype is present in a heterozygous or homozygous state in approximately 50% of the Pug population. However, it is fortunate that the susceptibility haplotype in Pugs only increases risk for NME when in the homozygous state. This allows breeders to maintain the susceptibility haplotype, albeit in a heterozygous state. The ability to maintain a susceptibility haplotype while reducing the incidence of a specific autoimmune disease will vary from breed to breed and from one disorder to another. If homozygosity for a specific allele or haplotype greatly increases risk, than selective mating against homozygosity may be preferred.
DQB1 genotype determined by single-nucleotide polymorphism (SNP) testing. e
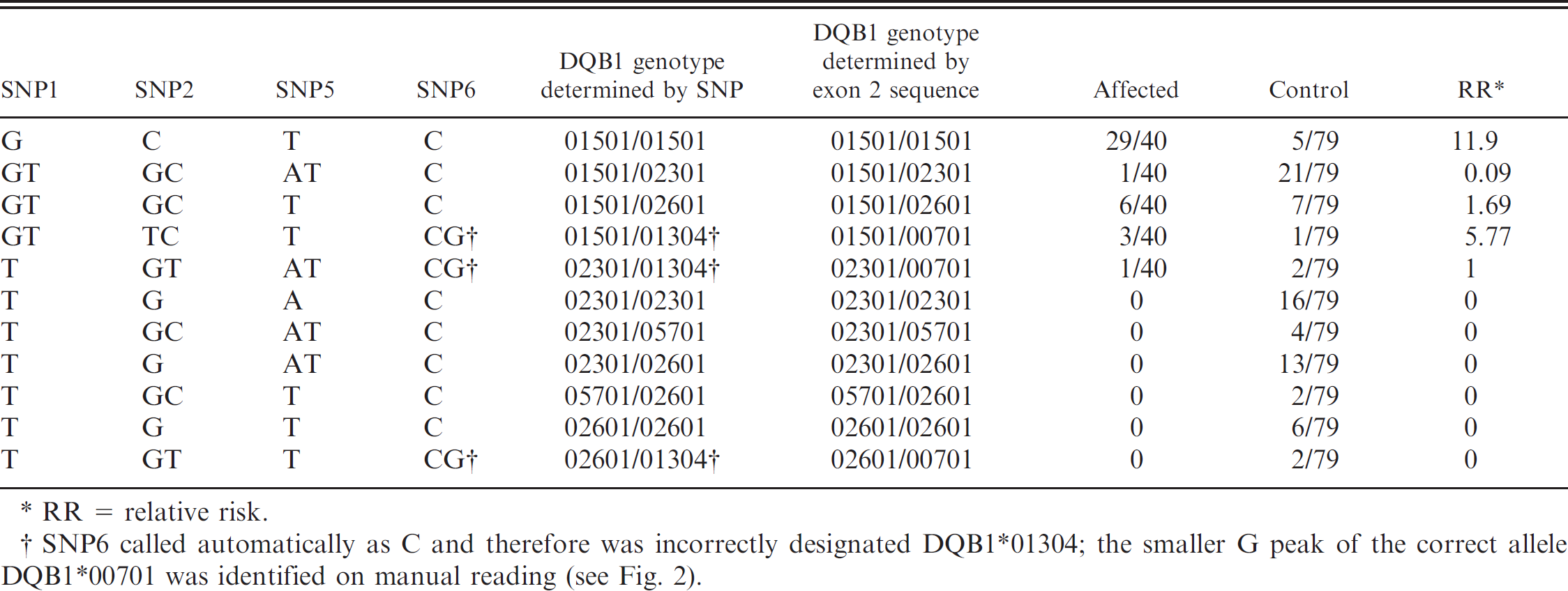
RR = relative risk.
SNP6 called automatically as C and therefore was incorrectly designated DQB1*01304; the smaller G peak of the correct allele DQB1*00701 was identified on manual reading (see Fig. 2).
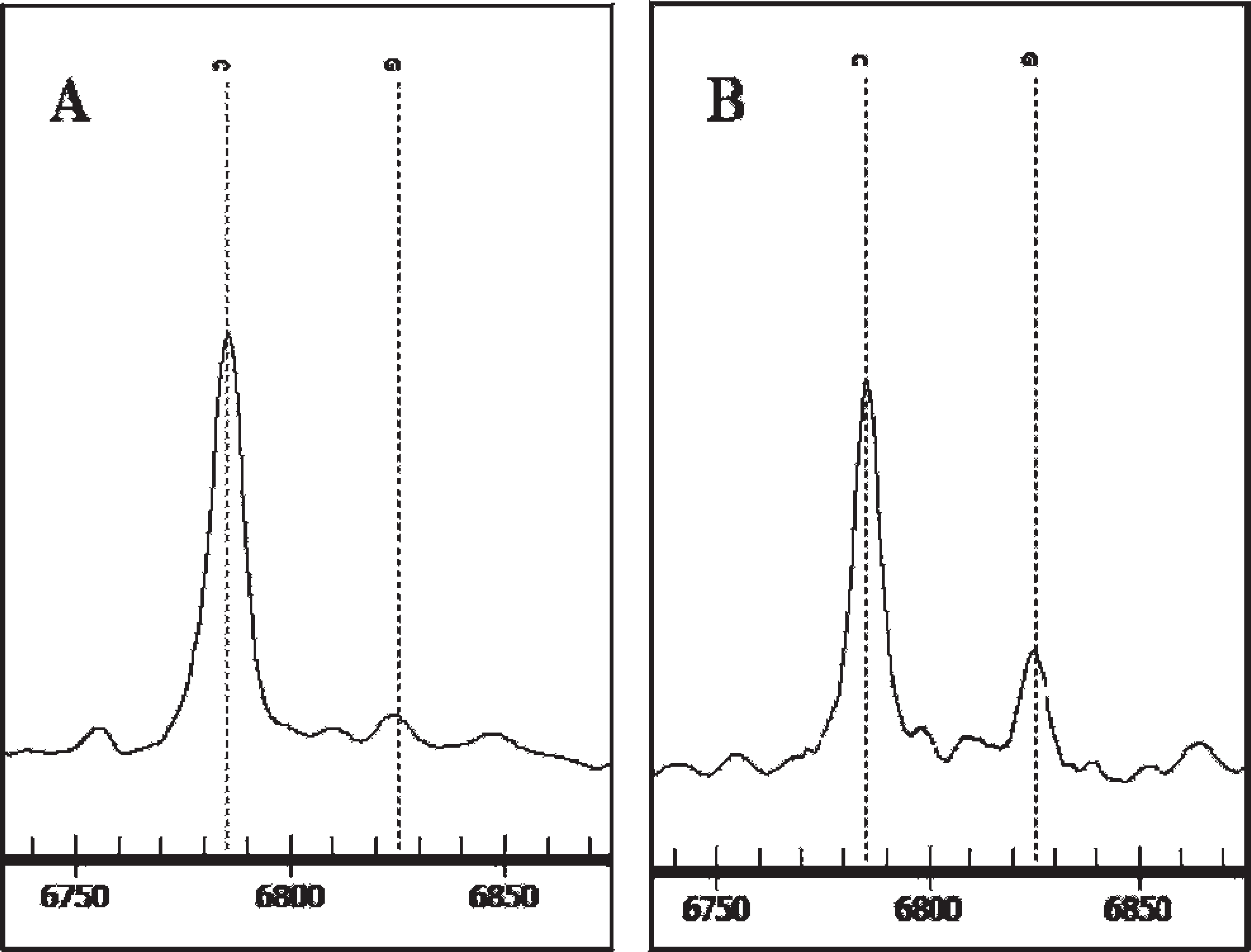
A, all DQB1 genotypes are homozygous (C/C) at single-nucleotide polymorphism 6 except for those containing DQB1*00701 (see panel B);
Acknowledgements
Funding for this study was provided by the Center for Companion Animal Health and the Veterinary Genetics Laboratory, School of Veterinary Medicine, University of California, Davis. The authors are grateful for the assistance of Dr. Phil Kass, University of California, Davis, with some of the statistical analyses and for the many Pug dog owners and breeders who contributed DNA samples from affected and healthy dogs.
Footnotes
a.
HotStarTaq® Plus, QIAquick®; Qiagen Inc., Valencia, CA.
b.
Big Dye® Terminator, ABI 3730; Applied Biosystems, Foster City, CA.
c.
Performa DTR Ultra, Edge BioSystems, Gaithersburg, MD.
d.
Vector NTI Advance™, Invitrogen Corp., Carlsbad, CA.
e.
MassARRAY® version 4.0.0.2, MassARRAY® Compact 96 System featuring iPLEX® Gold technology; Sequenom Inc., San Diego, CA.