
Abstract
Cryptococcus gattii was isolated from a 1.5-year-old dog with systemic cryptococcosis in Oregon. The dog had no link to Vancouver Island or British Columbia, Canada. Samples from a nasal swab and from a granulomatous mass within the cranial cavity were pooled for culture. Colonies on Sabouraud dextrose agar were mucoid and exhibited bimorphic morphology, melanin-pigmented and unpigmented. Pigmented colonies were encapsulated budding spherical yeast, whereas unpigmented colonies were of unencapsulated ovoid budding yeast. In addition to defective melanin production, the unpigmented colony type exhibited defective mating. Genetic analysis by high-resolution multilocus sequence typing revealed that the 2 isolates are genetically identical at 8 unlinked loci tested and that the 2 isolates are both the VGIIa Vancouver Island major genotype. Findings are consistent with expansion of the Vancouver Island outbreak onto the mainland Pacific Northwest region of the United States.
Cryptococcus gattii (formerly Cryptococcus neoformans var. gattii) is a haploid yeast formerly thought to be restricted to tropical and subtropical climates. 1,9,11 Infection is by inhalation of airborne propagules. 10,11 The pathogenicity of C. gattii differs from that of Cryptococcus neoformans in that C. gattii infection occurs most often in immunocompetent hosts. 1,10,11 Genotypic analysis of C. gattii has revealed 4 major molecular groups: VGI–VGIV. 6 Cryptococcus gattii has recently emerged as a human and animal pathogen, causing an outbreak on Vancouver Island, Canada, that began in 1999 and was ongoing at the time of this writing. 1 3,12,13 Molecular studies indicate a previously rare genotype to be the primary cause of the outbreak. 6,9 Molecular analysis of the mating type specific genes SXI1α and SXI1a of this genotype suggest that same sex-mating has resulted in a highly pathogenic strain within this region. 6 More recently, C. gattii infection has caused clinical disease in animals and humans elsewhere in the Pacific Northwest. 3,4,10 A confirmed case of C. gattii caused by the Vancouver Island outbreak major genotype, designated VGIIa, was found in a man from Puget Sound Washington, USA, in January 2006. 13 Cryptococcus gattii infection with the VGIIa genotype has also been reported in cats in Washington State, with no apparent link to Vancouver Island. 10
Cryptococcus gattii and C. neoformans grow readily on Sabouraud dextrose agar. Colonies can be smooth, mucoid, or wrinkled, with increased virulence associated with a mucoid or wrinkled phenotype. 8 Colony phenotype is related to the composition of the polysaccharide capsule, and phenotypic switching has been documented in laboratory studies of C. neoformans 7 and C. gattii. 8 Phenotypic alteration of C. gattii also occurs in vivo, and alteration in the polysaccharide capsule is related to the ability to cross the blood brain barrier during infection of the central nervous system. 8 This report describes a case of primarily central nervous system infection by C. gattii with bimorphic colony types in a dog from Western Oregon with no history of travel to Canada. Both isolates were confirmed to be the Vancouver Island outbreak genotype.
A 1.5-year-old, intact, female Keeshond dog was presented for necropsy. The dog was born in Hillsboro, Oregon, and had been relocated to Salem, Oregon, at the age of 7 weeks. The dog had traveled to Washington State, including visits to Seattle and the Puget Sound area of Washington, but there was no travel history to Vancouver Island or Canada. The dog died after an approximately 1-week history of cervical pain, ataxia, anorexia, vomiting, depression, and collapse. Gait abnormalities were characterized by bending and staggering to the left. Additional history indicated occasional bouts of lethargy, diarrhea, and vomiting for approximately 3 months before the recent severe illness. Vaccinations and deworming were current.
The dog weighed 11.4 kg and was in poor body condition. The most significant finding was a yellow, soft, gelatinous mass (approximately 3 cm × 2.5 cm × 1.5 cm) within the left cranial vault, loosely attached to the underside of the left cerebrum at the area of the left olfactory bulb. A cavitation within the adjacent brain exuded clear yellow viscous fluid when sectioned. The mass was adhered out to the inner surface of the cranium and extended to the cribriform plate, but did not involve the nasal cavity. Cytologic evaluation revealed heavily encapsulated yeast with narrow-based budding consistent with Cryptococcus spp. A swab of the left caudal nasal cavity and a swab of the mass in the cranial vault were obtained and pooled for fungal isolation. The lungs contained patchy red areas and mesenteric lymph nodes were diffusely enlarged (approximately 2–3 times normal). No other postmortem findings were significant. The spinal cord was not examined. The brain and attached mass and tissue samples from lung, kidney, pancreas, liver, adrenal gland, spleen, thyroid gland, intestine, and mesenteric lymph node were fixed in 10% neutral buffered formalin and prepared for routine histopathologic examination. Selected sections of the granulomatous intracranial mass, kidney, and lung were also stained with Mayer's mucicarmine, Grocott's methenamine silver (GMS), and periodic acid–Schiff (PAS) stains.
Sections of the left cranial mass and cerebrum revealed severe granulomatous meningitis and encephalitis with numerous encapsulated spherical fungal organisms approximately 4–20 μm in diameter exhibiting narrow-based budding (Figs. 1, 2). Collagen was prominent within the granulomatous mass external to the brain, indicative of chronicity. Small multifocal to locally extensive zones of granulomatous inflammation with similar fungal organisms were detected in the renal cortices, with a very small number of similar foci within the pancreas and lung. Mesenteric lymph nodes were hyperplastic with no evidence of fungal infection, and fungal infection was not detected in any other organ section examined. Fungal organisms stained with PAS and GMS stains, and the capsule stained intensely with Mayer's mucicarmine stain (Fig. 2). No variation in fungal morphology was detected in tissue sections.
Tissue swabs were pooled and inoculated onto blood agar a incubated at 35°C and Sabouraud dextrose agar b incubated at room temperature. Growth started at 2 days and consisted of 2 mucoid colony types: gray and white. India ink preparations of the gray colonies indicated growth of spherical yeast up to approximately 20 μmin diameter—some encapsulated, with little evidence of budding. Similar preparations of the white colonies revealed nonencapsulated slightly oval yeast of similar size with more frequent budding. Isolates exhibited a similar bimorphic growth pattern at 3 days when subcultured onto Sabouraud dextrose agar b and incubated at room temperature and at 35°C. Isolates of both colony types were urease-positive at 24 hr when grown on Christensen's urea agar slants a incubated at room temperature. Only spherical yeast cells with no pseudohyphae or hyphae were detected in colonies grown on cornmeal agar b at 30°C. The 2 isolates were determined to be Cryptococcus spp. and were sent to the Center for Microbial Pathogenesis, Duke University Medical Center (Durham, NC), for further analysis. Both isolates exhibited growth and blue coloration on canavanine–glycine–bromthymol blue agar c after 24 hr of incubation. Melanin production after inoculation onto L-dopamine agar media as per a previous study 2 and incubated at 37°C was also drastically reduced in the unpigmented colony type (Fig. 3). The unpigmented colony type exhibited a severe fertility defect, characterized by lack of filamentous growth on Murashige and Skoog media with myo-inositol incubated at room temperature in the dark for approximately 7 days in dry conditions as per a previous study 14 (Fig. 4).
In addition to phenotypic analysis, multilocus sequence typing at 8 unlinked loci previously shown to characterize C. gattii outbreak strains was applied to the variant colony morphologies as previously described. 6 Both isolates harbored the SXI1α gene, and were therefore of the α mating type. 6 Furthermore, both isolates were identical across all 8 gene loci tested and were of the VGIIa molecular type, the major Vancouver Island genotype (Fig. 5).
The clinical signs in this case were primarily neurologic and were consistent with the pathologic findings. Central nervous system infection is common in dogs with C. gattii infection, and the presence of clinical signs of central nervous system disease has been shown to be a significant predictor of mortality. 3 No underlying disease process that might decrease immune function was identified in this dog, consistent with prior reports of C. gattii infection in immunocompetent hosts. 1,10,11
It is suggested that the bimorphic colony types might reflect differences in the nature of the organism in the intracranial granuloma versus that present in the nasal cavity. Perhaps 1 isolate infected the animal, and then an in vivo colony phenotypic switch either resulted in dissemination or occurred as a consequence of dissemination. The swabs from both areas were pooled for culture, however, and this remains a hypothesis. Whether phenotypic switching is genetically or epigenetically controlled is not yet known. This case illustrates that phenotypic variants isolated from a single patient can differ in melanin production and in mating ability while maintaining similar genetic profiles.
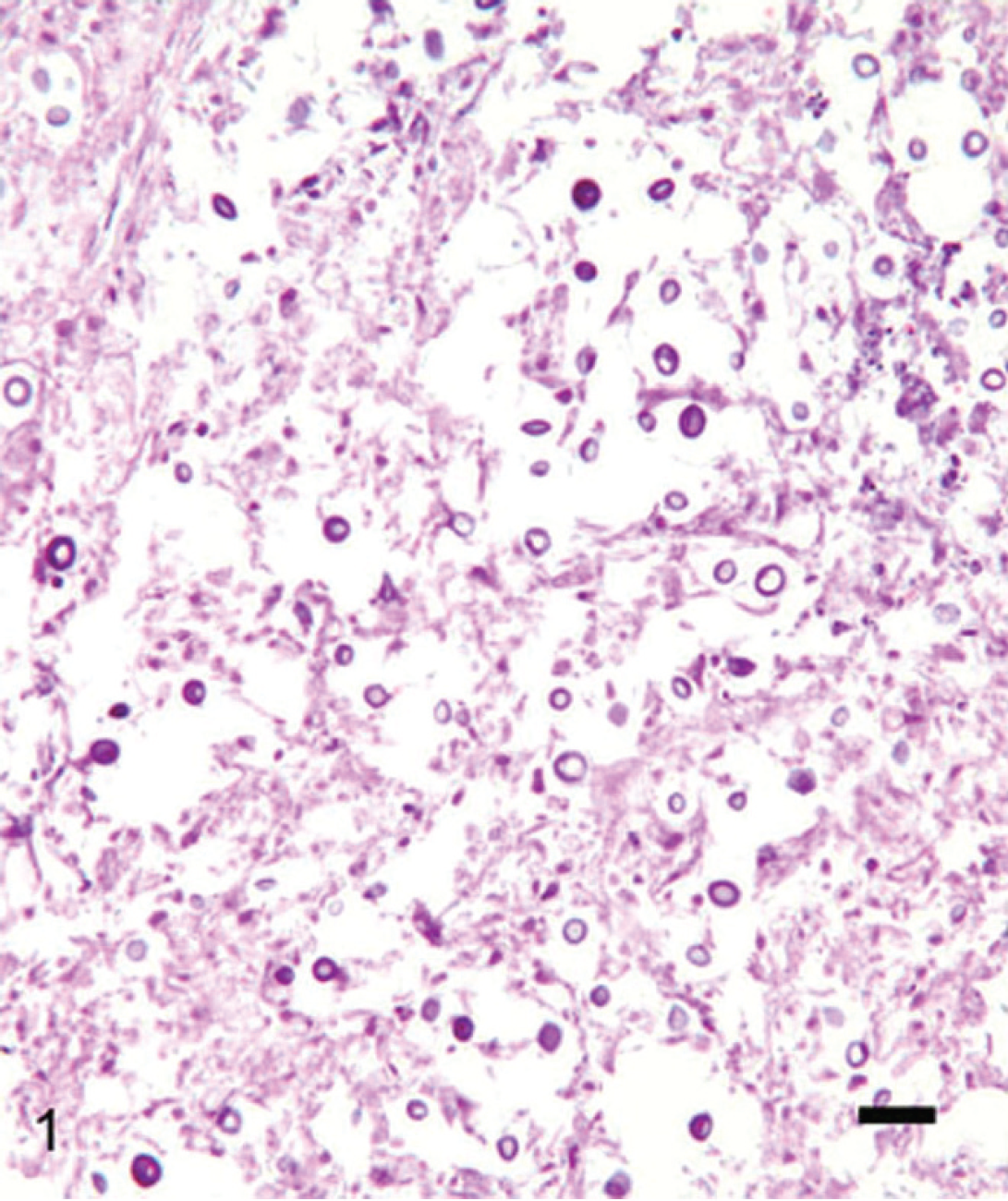
Granulomatous mass from the cranial cavity of a 1.5-year-old dog. There are many encapsulated spherical to oval yeast. Hematoxylin and eosin. Bar = 40 μm.

Yeast exhibiting narrow-based budding with intense capsular staining. The organism is surrounded by inflammation consisting of macrophages, lymphocytes, and plasma cells. Mayer's mucicarmine stain. Bar = 10 μm.

Colonies exhibit bimorphic morphology on L-dopamine agar. Both are mucoid, with melanin production in the colony on the right and lack of melanin production in the colony on the left.

Mating differences on Murashige and Skoog media with myo-inositol, incubated at room temperature in the dark. There is a lack of fertility in the yeast from the colony lacking melanin production, evidenced by lack of filamentous growth (

Multilocus sequence typing (MLST) analysis of the isolates with controls, performed for 8 unlinked loci. Numbers and color-coding represent different alleles designated by genetic sequence variation, with allele numbers and designations used in the current study, including GenBank accession numbers 6 (http://www.nature.com/nature/journal/v437/n7063/extref/nature04220-s7.xls). Isolate R272 represents the VGIIb minor outbreak genotype control; isolate R265 represents the VGIIa major outbreak genotype control; and isolates WM276, WA 861, NIH312, and MMRL2651 represent the VGI–VGIV controls for each molecular group, respectively. 6 Isolates EJB17a and EJB17b from the dog in this report each share identical genotypic profiles with R265 at these 8 loci, indicating they are VGIIa outbreak genotype isolates.
The majority of the infection in this case was adjacent to the cribriform plate in the cranium and in the left cranioventral cerebrum. Subclinical nasal infection by C. gattii in dogs and cats can progress to systemic infection, 4 which is the suspected pathogenesis in this case. This case illustrates the recent emergence of C. gattii in both veterinary and human patients in the Pacific Northwest. The genetic concordance of this isolate with the outbreak major genotype indicates that this case is an expansion of the geographic range of this virulent strain from Vancouver Island to the Pacific Northwest of the United States. An alternative hypothesis is that the Vancouver Island VGIIa major genotype was present in both Vancouver Island and the United States and emerged independently in the 2 locales. However, because no cases in animals or patients in Washington and Oregon were reported from 1999 until the first index case in 2006 in the San Juan Islands, during which time hundreds of cases occurred on Vancouver Island in animals and humans, 10 the more parsimonious explanation is favored that this represents an expansion from Vancouver Island, Canada, into the United States. Moreover, ample evidence indicates that the organism is spread via a variety of routes, including human traffic, 1 and the spate of several dozen recent cases in animals and humans in Washington and Oregon during 2007–2008 (Byrnes, unpublished observations) is in accord with models positing a recent expansion of geographic range by introduction from Vancouver Island, Canada, into the United States. This case also reflects an important need for genotypic analysis of veterinary isolates of Cryptococcus spp. infections to determine both the species causing infection and the molecular type of the isolate to establish modes and patterns of this emerging pathogenic fungus in the Pacific Northwest.
Acknowledgements. The authors thank Judi James for providing clinical information and history of the animal. This research was supported by NIH/NIAID R01 grant AI39115 to Joseph Heitman.