
Abstract
Porcine circovirus-2 (PCV-2) is an economically important swine pathogen and causes PCV-associated disease (PCVAD) in pigs worldwide. Currently, 2 genotypes of PCV-2, PCV-2a and −2b, are circulating in U.S. swine herds. The objectives of the current study were to evaluate the amount of PCV-2 DNA present in semen over time, compare and correlate incidence and amount of PCV-2 present in semen samples to that present in serum samples and blood swabs, and determine if there are differences in shedding patterns between PCV-2a and −2b. Fifteen 7-month-old PCV-2-naïve Landrace boars (Sus scrofa) were randomly allocated to 3 treatment groups. The boars in group 1 (n = 3) served as negative controls, and those in groups 2 (n = 6) and 3 (n = 6) were intranasally and intramuscularly inoculated with PCV-2a and −2b, respectively. Semen, serum, and blood swab samples were collected up to 90 days postinoculation (DPI), and necropsies were performed on DPI 23,48, and 90. Larger quantities of both PCV-2a and − 2b DNA were detected earlier in serum and blood swab samples than in raw semen of experimentally inoculated boars. The incidence and duration of presence of PCV-2 DNA in semen varied among boars; however, intermittent shedding was not observed. In all sex glands, PCV-2 DNA was detected by polymerase chain reaction; however, PCV-2 antigen was not detected by immunohistochemistry, and PCV-2 had no effect on sperm morphology. Differences in shedding patterns between PCV-2a and −2b were not observed under the study conditions.
Introduction
Porcine circovirus-2 (PCV-2; family Circoviridae, genus Circovirus) is a small, nonenveloped, closed, circular, single-stranded DNA virus that has been associated with many disease entities including weight loss, pneumonia, diarrhea, and reproductive problems in swine. 27 Porcine circovirus-associated disease (PCVAD) was first reported in high-health herds in Canada in 1991 12 and has since been reported in swine worldwide. 39
Transmission of PCV-2 in swine populations is not well understood; however, the main route of infection is thought to be fecal-oral. 39 Detection of PCV-2 DNA has been reported in secretions and excretions including feces, urine, saliva, ocular fluid, nasal secretions, colostrum, and semen of infected pigs. 4 , 19,21 , There has been a marked increase in the incidence of PCVAD in North America, and it is thought by some to be associated with the introduction of PCV-2b, a distinct cluster of PCV-2. 9 , 14 Although previous epidemiologic investigations found no association with the use of artificial insemination (AI) as a risk factor for PCVAD (Lawton DE, Morris RS, King CM: 2004, PMWS in New Zealand part 2: epidemiological evidence for a novel agent. Proceedings of the 18th International Pig Veterinary Society Congress 1:128), 6 , 35 , 43 AI has not been ruled out as a way to quickly disseminate PCV-2 through populations.
Porcine circovirus-2 DNA in semen has been detected by nested 17 , 18 , 21 , 22 and real-time quantitative polymerase chain reaction (nPCR and real-time qPCR, respectively) 30 in both naturally and experimentally infected boars and, typically, is not associated with clinical disease in sexually mature boars. Previously, experimental inoculation of PCV-2 in naïve boars revealed that PCV-2 DNA was detected by nPCR as early as 5 days postinoculation (DPI) and intermittently thereafter up to DPI 47. 21 Furthermore, shedding of PCV-2 DNA in semen was detected up to 27.3 weeks in naturally infected boars; younger boars (<12 months of age) were more likely to shed PCV-2 DNA in semen, and PCV-2 infection had no effect on sperm morphology. 22 Previous reports have most consistently demonstrated PCV-2 DNA in the nonsperm fractions of raw semen in naturally infected boars. 17 In contrast, more recent reports have demonstrated that PCV-2 DNA is found in higher concentrations in the sperm-rich fraction when compared to seminal plasma fractions. 30 In addition, when raw semen was compared to sperm-rich fractions of experimentally inoculated boars, similar quantities of PCV-2 DNA were detected by real-time qPCR. 30
The prevalence of PCV-2 DNA in the semen of naturally infected boars as detected by nPCR is variable. 17,18,22 Previous reports indicate that PCV-2 DNA was detected in 13 of 43 Canadian boars, originating from a single boar stud, with an overall detection rate of 3.3% (30/903 semen samples). 22 In 30 Korean swine herds, PCV-2 DNA was detected in 28 of 60 (47%) semen samples tested. 17 The same group detected PCV-2 DNA in 26 of 98 (27%) semen samples obtained from 49 Korean swine herds. 18
Porcine circovirus-2 viral antigen detection by immunohistochemistry (IHC) has been documented in the reproductive organs of naturally and experimentally infected boars and was present in infiltrating macrophages of the tunic albuginea, interstitial macrophages, and germinal epithelial cells in the testes and in infiltrating macrophages of the epididymis in 4-week-old piglets 3 weeks after experimental coinoculation with Porcine parvovirus (PPV) and PCV-2. 16 Natural PCV-2 infection in a clinically affected mature boar was associated with the presence of abundant PCV-2 antigen in the interstitium of seminal vesicles. 25
The objectives of the current study were to evaluate the amount of PCV-2 present in semen over time, correlate incidence and amount of PCV-2 present in semen compared to incidence and amount present in serum and blood swab samples, and determine if there are differences in semen-shedding patterns between PCV-2a and −2b. This information could be used to formulate management decisions in boar studs and breeding herds.
Materials and methods
Animals and housing
Fifteen purebred, male, Landrace pigs (Sus scrofa) were segregated early, weaned from a single farm, and brought to the Iowa State University infectious disease isolation facility (Ames, IA) at approximately 3 weeks of age. The source farm was known to be free of Porcine reproductive and respiratory syndrome virus (PRRSV) and Swine influenza virus by frequent serologic surveillance of the herd. Animals were group housed until 6 months of age and then were individually penned. The pens were 2 m × 2 m × 1.5 m with solid partitions. Each room had a separate collection pen of similar dimensions with a mounting apparatus for semen collection. Boars were fed approximately 2.5 kg of a balanced corn-soybean base ration daily.
Experimental design
At 7 months of age, boars were randomly allocated into 3 groups. The boars in group 1 (n = 3) served as negative controls, and those in groups 2 (n = 6) and 3 (n = 6) were inoculated with PCV-2a or −2b, respectively. Randomly selected boars from each group were euthanized at 3 different time points during the study by intravenous pentobarbital. Necropsies were performed on 1 boar from group 1 and 2 boars each from groups 2 and 3 at DPI 23, 48, and 90. The experimental protocol was approved by the Iowa State University Institutional Animal Care and Use Committee.
Inoculation
Boars in group 2 were inoculated intranasally (3 ml) and intramuscularly (2 ml) with a stock of PCV-2a isolate ISU-40895 with an infectious titer of 104.5 TCID50 (50% tissue culture infective dose) per ml. The PCV-2a isolate was originally obtained from a clinical PCVAD-affected pig in a western Iowa herd in 1998 and has been shown to cause PCVAD in previous experimental studies using growing pigs. 24 , 28 Boars in group 3 were inoculated intranasally (3 ml) and intramuscularly (2 ml) with a stock of PCV-2b isolate NC-16845 with an infectious titer of 104.3 TCID50 per ml. The PCV-2b isolate was acquired from a finishing pig affected with severe PCVAD in North Carolina in 2006. Both PCV-2a and −2b infectious virus stocks were similarly generated by transfecting porcine kidney (PK)-15 cells with respective PCV-2a and −2b infectious DNA clones, as described previously. 8 The PCV-2a and −2b isolates shared 95.7% nucleic acid homology. The virus inocula were stored at −80°C until experimental inoculation.
Sample collection
Semen and serum samples were collected from all boars prior to PCV-2 inoculation on DPI 3. Semen, serum, and blood swabs 34 were collected on DPI 2, 4, 6, 9, 13, 16, 20, 23, 27, and every week thereafter until DPI 90. In addition, serum samples were collected on DPI 1 from all boars. For sample collection, individual animals were moved into the collection pen and, once mounted on the collection apparatus, gloved-hand semen collection was initiated using an insulated mug lined with a 2-liter disposable plastic bag with a tear-away filter. a Post ejaculation, the filter portion of the collection bag containing the gel fraction was torn off and discarded. Bags containing raw semen were sealed, immediately placed into a thermal container, and transported to the laboratory for processing.
Concurrently, during semen collection, serum and blood swabs were obtained while the boar was mounted on the collection apparatus. Whole blood was collected into a 300-μl serum gel capillary tube by venipuncture of an ear vein. b Similarly, blood swabs were obtained using individually packaged sterile swabs c and placed into a sterile 5-ml polystyrene round-bottom tube d containing 0.5 ml of sterile saline. After capillary tubes were centrifuged, the serum and blood swabs were stored at −80°C until tested.
Semen sample processing, morphology, and serology
Collection bags containing raw semen samples were gently agitated by inverting the sealed bag to equally resuspend spermatozoa after transportation. Raw semen was carefully allocated into a sterile 1.5-ml centrifuge vial using a sterile 10-ml graduated serology pipette and stored at −80°C until testing.
For morphology analysis, well-mixed raw semen from each ejaculate was initially diluted at a rate of 1:10 to 1:20 with prewarmed extender. e Subsequently, the diluted semen was mixed with an equal volume of buffered formal saline f for preservation. A full morphology differential was performed on each sample with wet mounts and a phase contrast microscope at 1,000X under oil. 3 , 15 , 20
Serum samples were tested for the presence of anti-PCV-2 immunoglobulin G (IgG) antibodies using an enzyme-linked immunosorbent assay (ELISA) based on PCV-2 recombinant major capsid protein (open reading frame 2 [ORF2]), as previously described. 23 Samples with a sample-to-positive ratio of ≥0.2 were considered positive.
Polymerase chain reaction
Serum, blood swab samples, and raw semen were tested for the presence and amount of PCV-2 DNA by real-time qPCR, as previously described. 29 , 30 An internal positive control composed of a PCV-2 genome clone with 25 nucleotide substitution beginning at position 1582 (mutated PCV-2) was added to the raw semen samples prior to DNA extraction for quality control, as previously described. 30 In addition, select tissue samples from each boar (lymph node, testes, epididymis, bulbourethral gland, seminal vesicle, and prostate) collected at necropsy were also tested for PCV-2 DNA. Briefly, DNA was extracted from 0.25 g of tissue using a commercially available isolation kit g following the manufacturer's protocol and recommendations. The PCR parameters used in the current study were essentially the same as previously described. 29
Necropsy
At necropsy, lungs were evaluated for the presence of pneumonia lesions (0-100% of the lung surface affected by lesions), 11 lymph nodes were examined for evidence of enlargement, and additional gross observations of other organ systems were recorded. Sections of all major organ systems were collected in 10% buffered formalin. Tissue sections collected included testes, epididymis, seminal vesicles, bulbourethral gland, prostate, penile tissue, 4 lung sections (cranial, middle, caudal, and accessory lobes), liver, spleen, kidney, tonsil, ileum, colon, thymus, lymph nodes (tracheobronchial, mediastinal, mesenteric, iliac, and superficial inguinal), myocardium, and urinary bladder.
Microscopic evaluation and immunohistochemistry
Formalin-fixed tissue samples were routinely processed, stained with hematoxylin and eosin, and evaluated in a blinded fashion by a veterinary pathologist. Microscopic lesions characteristic of PCV-2 infection (depletion of germinal center lymphocytes and histiocytic replacement or infiltration) in lymphoid tissues (lymph nodes, thymus, tonsil, and spleen) were scored, as previously described. 28 Immunohistochemistry staining using rabbit polyclonal antisera for PCV-2 antigen detection was performed on lymphoid tissues, testes, epididymis, seminal vesicles, bulbourethral gland, prostate, and penile tissue. 42 The presence and amount of PCV-2 antigen was scored, as previously described. 28
Statistical analysis
Data were assessed for overall quality, including normality. Differences in amount of PCV-2 DNA in serum, semen, and blood swab samples were analyzed using repeated multivariate analysis of variance, with DPI as a fixed factor, followed by nonparametric Kruskal-Wallis one-way analysis of variance (ANOVA). Data until DPI 48 was included for the analysis. Differences in amount of PCV-2 DNA in tissues (lymph node, testes, epididymis, seminal vesicles, bulbourethral gland, and prostate) and normal sperm morphology were assessed using nonparametric Kruskal-Wallis one-way ANOVA. Calculations were performed using statistical software. h Outliers were excluded from the analysis.
Results
Serology
All boars had passively acquired anti-PCV-2 antibodies at weaning, which decayed as expected. By 4 months of age, all 15 boars were negative for anti-PCV-2 antibodies. Group 1 boars (negative controls) remained negative for anti-PCV-2 antibodies for the duration of the study (Table 1). After experimental inoculation at approximately 7 months of age, anti-PCV-2-specific IgG antibodies were first detected on DPI 13 in 3 of 6 boars from group 2 (PCV-2a inoculated), and all boars in this group had detectable anti-PCV-2 antibodies by DPI 23. Five of 6 boars in group 3 (PCV-2b inoculated) developed anti-PCV-2-specific antibodies between DPI 16 and 27. The PCV-2b boar that did not develop anti-PCV-2 antibodies was euthanized at DPI 23 as predetermined by the experimental design.
Quantitative real-time PCR
Serum samples. Porcine circovirus-2 DNA was not detected in serum samples from the boars in group 1 during the study (Table 1), but was detected in serum samples from all boars in group 2 by DPI 6, with the earliest detection on DPI 2 in 3 of 6 boars. Length of viremia ranged from 18 to 43 days among individual boars in group 2, with the last detection of PCV-2 DNA in serum samples on DPI 48. Porcine circovirus-2 DNA in serum samples from boars in Group 3 was first detected on DPI 6 in 3 of 6 boars, and all boars in this group became viremic by DPI 16. Length of viremia ranged from 8 to 84 days among individual boars in Group 3, and PCV-2 DNA was still present in serum samples from one of the boars at the termination of the study on DPI 90. Figure 1 summarizes the mean log transformed amounts of PCV-2 DNA in positive serum samples during the study.
Summary of pigs that were Porcine circovirus-2 (PCV-2) DNA positive in serum, blood swab, and semen as determined by real-time quantitative polymerase chain reaction (PCR) and pigs that were anti-PCV-2 antibody positive as measured by enzyme-linked immunosorbent assay (ELISA) for noninoculated (group 1) boars and boars inoculated with PCV-2a (group 2) or PCV-2b (group 3) over 90 days postinoculation (DPI).*
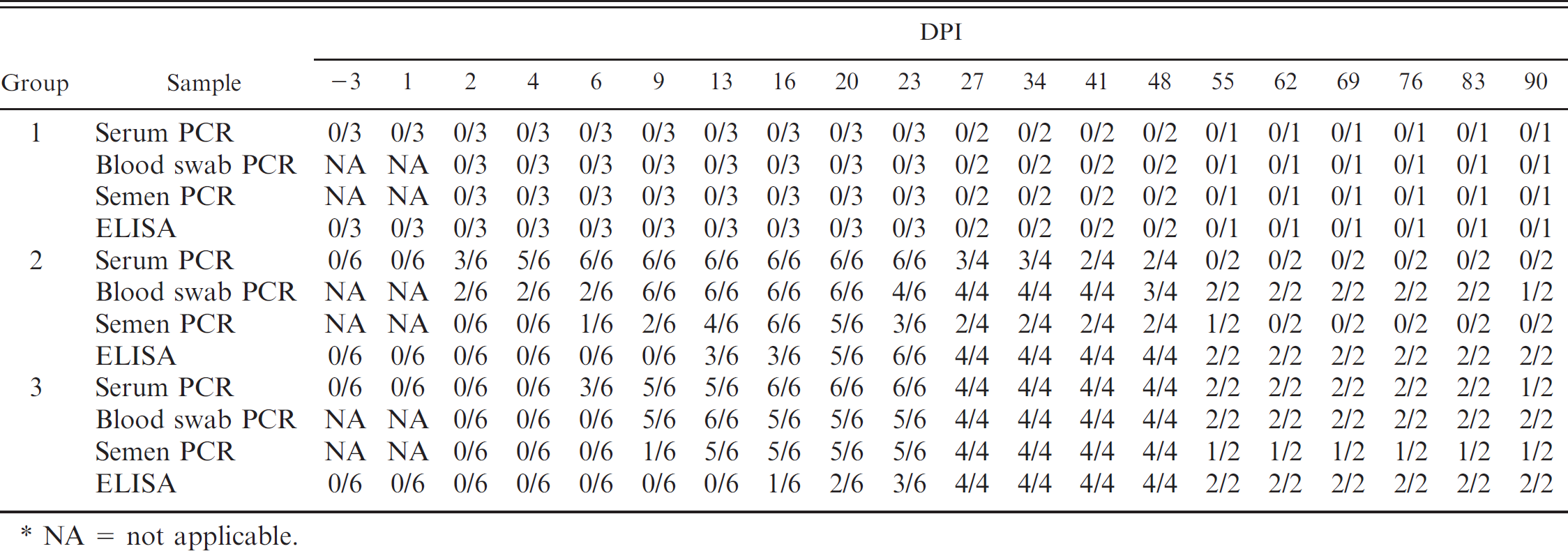
NA = not applicable.
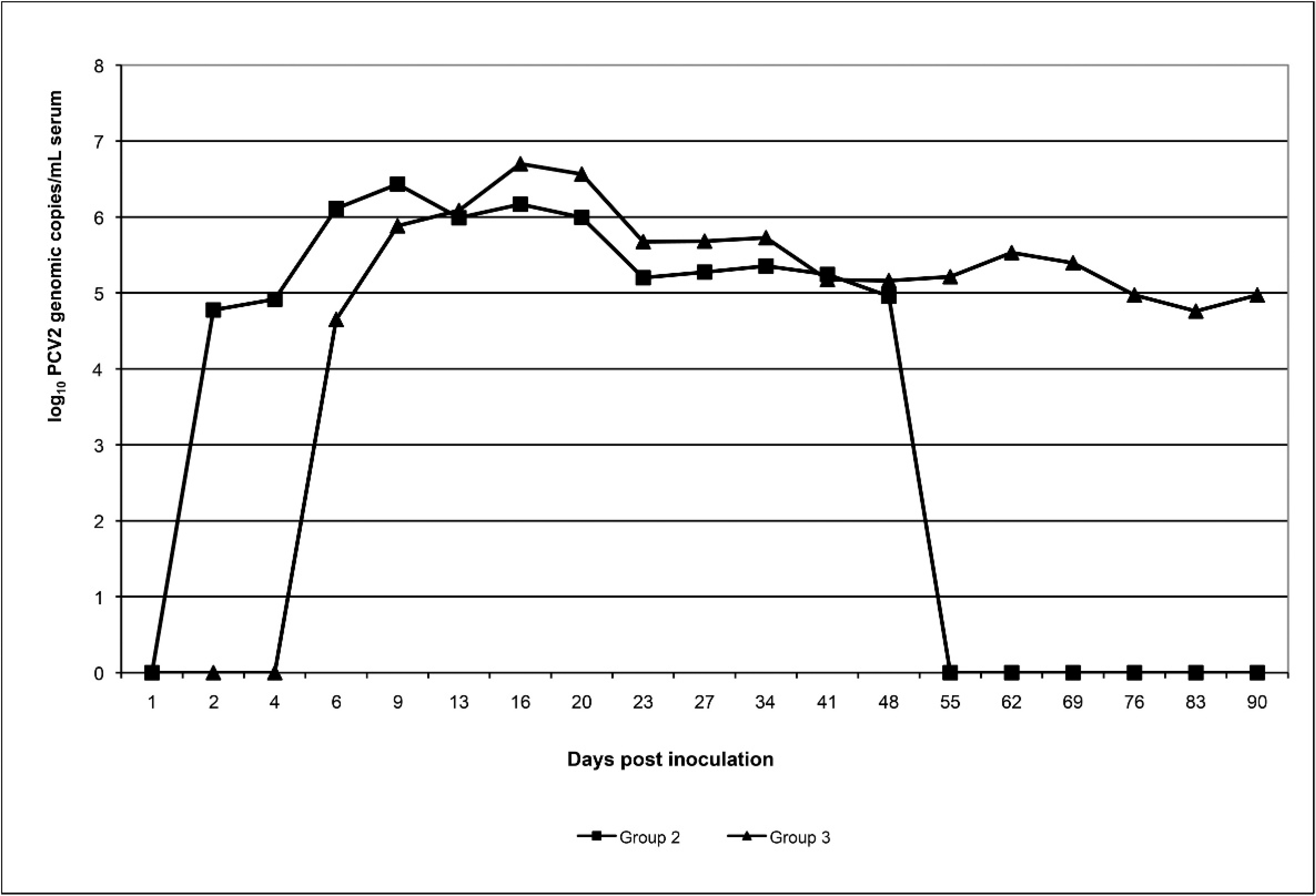
Mean Porcine circovirus-2 (PCV-2) genomic copies/ml (log transformed) in serum of boars inoculated with PCV-2a (group 2: n = 6, days postinoculation [DPI] 1–23; n = 4, DPI 27–48; n = 2, DPI 55–90) and boars inoculated with PCV-2b (group 3: n = 6, DPI 1–23; n = 4, DPI 27–18; n = 2, DPI 55–90). For all collection days, only positive serum samples were included in the group mean.
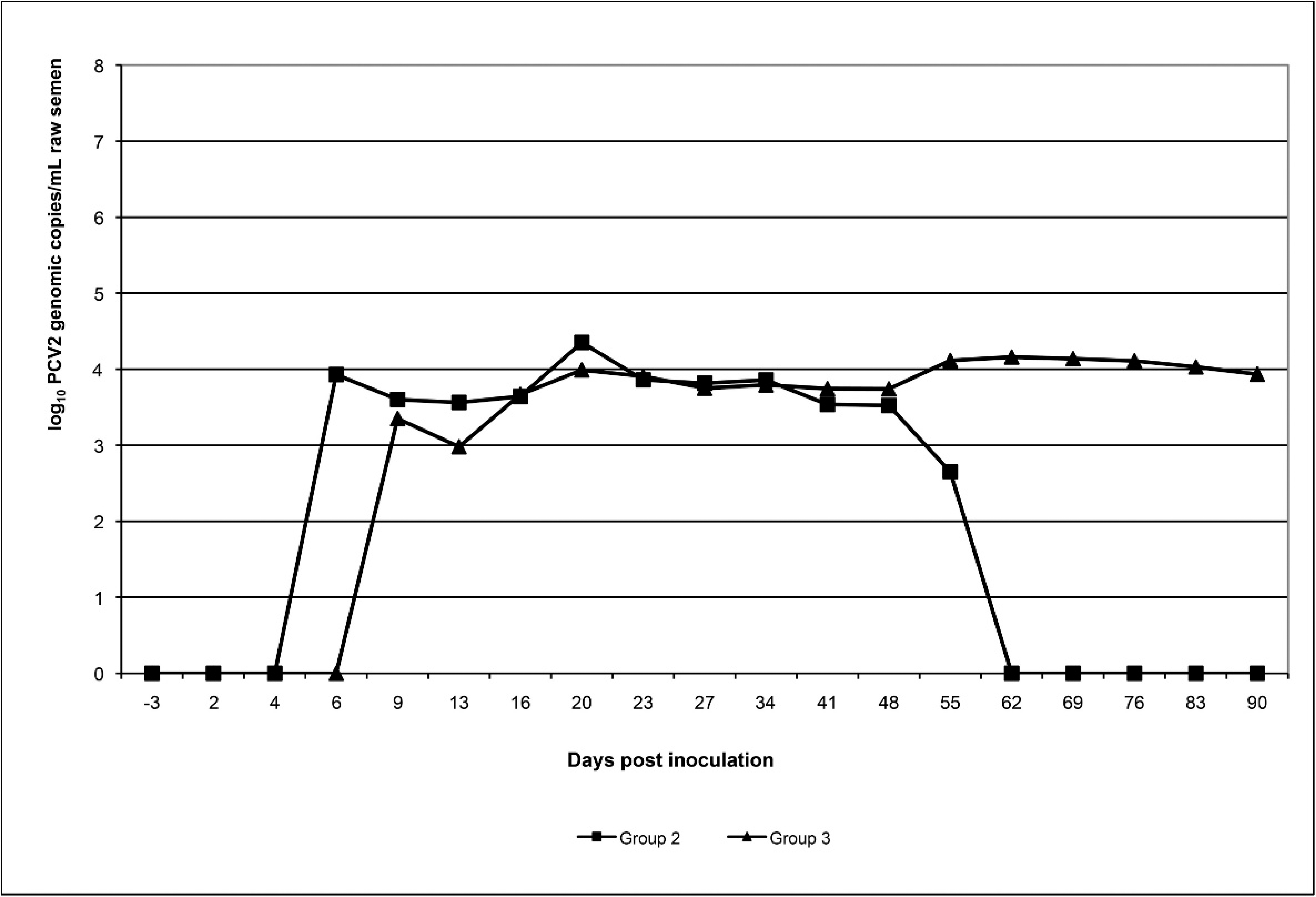
Mean Porcine circovirus-2 (PCV-2) genomic copies/ml (log transformed) in semen of boars inoculated with PCV-2a (group 2: n = 6, days postinoculation [DPI] 3–23; n = 4, DPI 27–48; n = 2, DPI 55–90) and boars inoculated with PCV-2b (group 3: n = 6, DPI 3–23; n = 4, DPI 27–48; n = 2, DPI 55–90). For all collection days, only positive semen samples were included in the group mean.
Semen samples. Inhibition of PCV-2 DNA detection in semen samples was not observed in any of the semen samples, as noted by consistent internal positive control levels. Porcine circovirus-2 DNA was not detected in raw semen samples collected from boars in group 1 during the study (Table 1). The amount of detectable PCV-2 DNA in semen varied among individual boars in group 2, from 221 to 103,336 genomic copies/ml of raw semen, with peak shedding between DPI 16 and 23. All boars in group 2 were PCR positive on raw semen, with shedding intervals ranging from 1 collection point to 11 consecutive collections (49 days). Intermittent shedding was not observed. In group 3, 5 of 6 boars shed PCV-2 DNA in raw semen, with first and last detection on DPI 9 and 90, respectively. The amount of detectable PCV-2 DNA in positive semen samples was similar to that of boars in group 2, and shedding intervals ranged from 0 to 77 days (Fig. 2).
Blood swab samples. Blood swab samples from group 1 were negative for PCV-2 DNA at all collection time points tested (Table 1). Porcine circovirus-2 DNA in blood swab samples from boars in group 2 was first detected on DPI 2, and all boars were positive for PCV-2 DNA by DPI 9 and remained positive until termination of the study. Earliest detection of PCV-2b DNA in boars from group 3 was on DPI 9; PCV-2 DNA was present in all blood swab samples by DPI 13, and PCV-2 DNA was still present in blood swab samples on DPI 90. The mean log transformed PCV-2 genomic copy numbers in positive blood swab samples over time are summarized in Figure 3.
Porcine circovirus-2 DNA was not detected in any semen samples on DPI 2 (0/12) and 4 (0/12). For DPI 6 and 9, 1 of 12 and 3 of 12 animals were PCV-2 PCR-positive on semen, and only data starting from DPI 13 were used for statistical analysis. Porcine circovirus-2 DNA was detected in 2 of 12 blood swab samples from 2 of 12 boars on DPI 6, and data prior to DPI 9 were excluded from the statistical analysis. The amount of PCV-2 DNA in serum samples was significantly higher (P < 0.05) than in semen from DPI 13 to 20. Similarly, the amount of PCV-2 DNA in serum samples was significantly higher (P < 0.05) than in blood swabs from DPI 13 to 20 (P < 0.05), and PCV-2 DNA was detected earlier in serum samples than in semen samples or blood swabs.
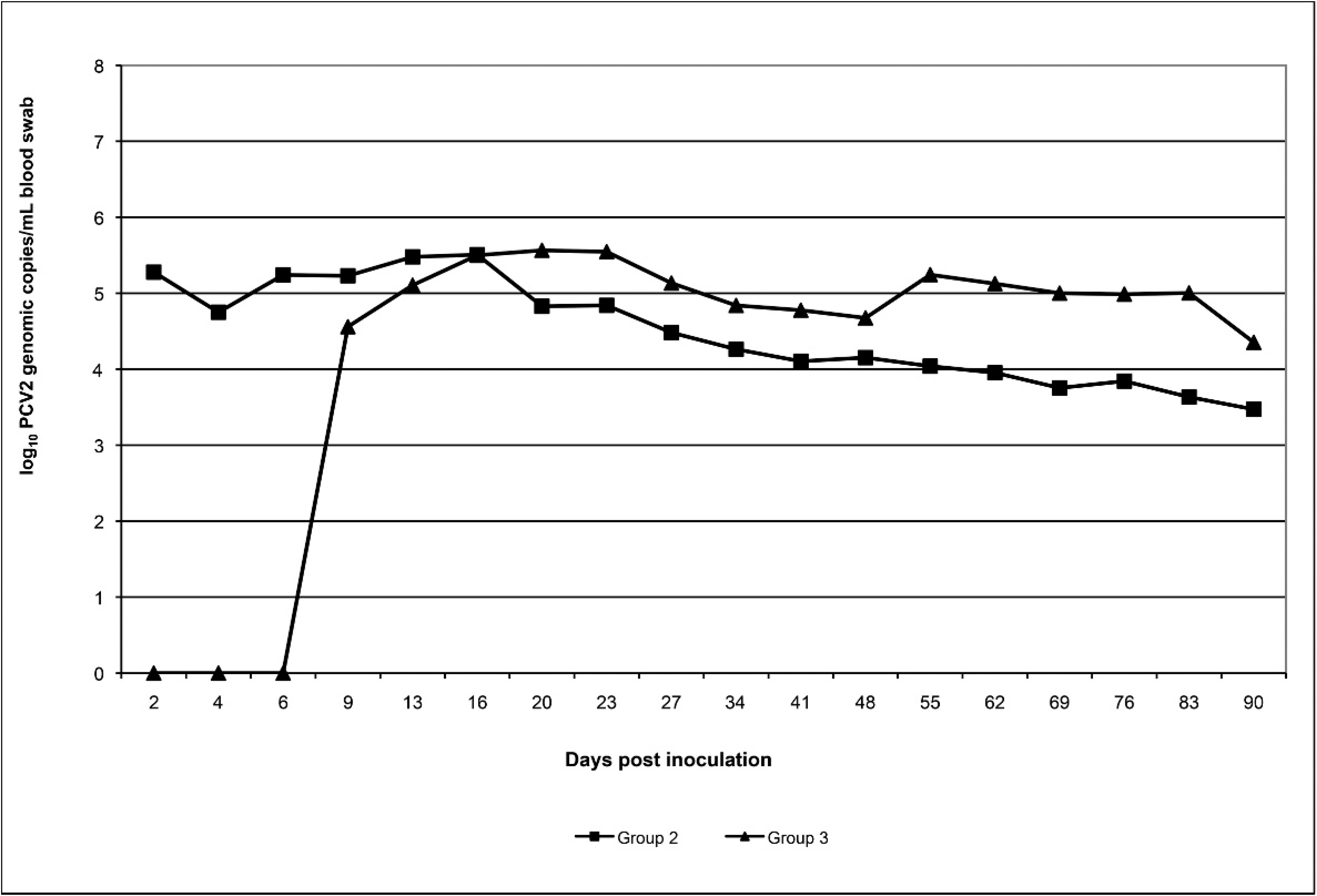
Mean Porcine circovirus-2 (PCV-2) genomic copies/ml (log transformed) present in blood swabs of boars inoculated with PCV-2a (group 2: n = 6, days postinoculation [DPI] 2–23; n = 4, DPI 27–48; n = 2, DPI 55–90) and boars inoculated with PCV-2b (group 3: n = 6, DPI 2–23; n = 4, DPI 27–48; n = 2, DPI 55–90). For all collection days, only positive blood swab samples were included in the group mean.
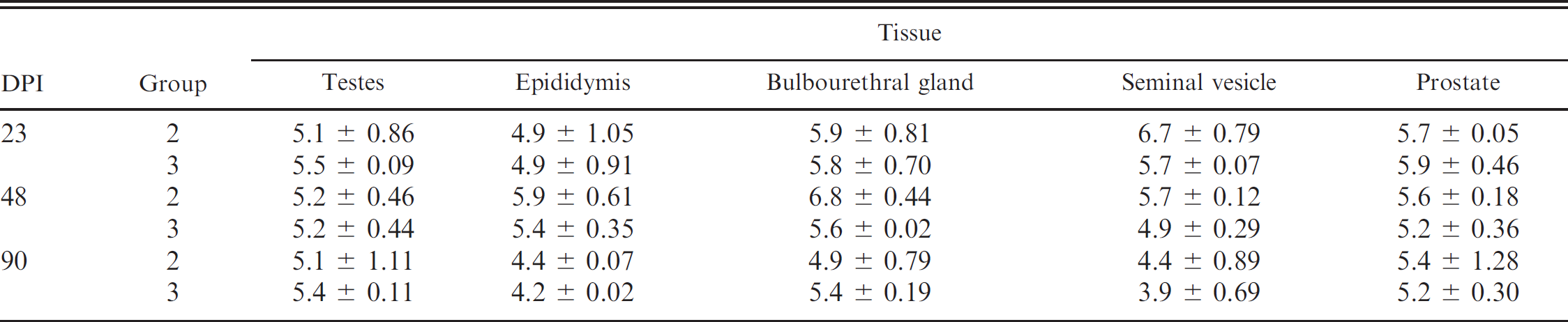
Tissue samples. Boar tissue samples (lymph nodes, testes, epididymis, bulbourethral gland, seminal vesicle, and prostate) from group 1 were negative for PCV-2 DNA on DPI 23, 48, and 90. In contrast, lymph nodes and sex glands tested from all boars in groups 2 and 3 were positive for PCV-2 DNA on each necropsy day. The group mean log transformed PCV-2 genomic copy numbers in 0.25 g of lymph node tissue for boars in groups 2 and 3 was 10 6 ± 0.15 and 106.1 ± 0.62, respectively (data not shown). Table 2 summarizes the amount of PCV-2 DNA (genomic copy numbers per 0.25 g of tissue) in the sex glands of PCV-2-inoculated boars. Significant differences in the amount of PCV-2 DNA present in tissues were not observed between groups 2 and 3 or when compared by DPI (P > 0.05).
Morphology
Mean percent normal sperm morphology prior to experimental inoculation was 79%,70%, and 68% for boars in groups 1, 2, and 3, respectively. Sperm morphology was not significantly different between PCV-2-inoculated and noninoculated boars or PCV-2a and −2b groups during weekly morphology evaluations throughout the study (data not shown).
Summary of log transformed group mean ± standard error of Porcine circovirus-2 (PCV-2) genomic copies in 0.25 g of tissue from the testes, epididymis, bulbourethral gland, seminal vesicle, and prostate of boars inoculated with PCV-2a (group 2) and PCV-2b (group 3) at necropsy (days postinoculation [DPI] 23, 48, and 90).
Macroscopic and microscopic evaluation
At DPI 23, no macroscopic or microscopic lesions were present in boars from group 1. Macroscopic lesions in boars from group 2 included moderately enlarged lymph nodes (2/2) and mottled, tan lungs (2/2), with 18–29% of the lungs grossly affected by lesions. Microscopically, 2 of 2 boars in group 2 had mild to moderate nonsuppurative interstitial pneumonia, 1 of 2 had mild lymphoid depletion, and 1 of 2 had mild, multifocal, lymphocytic interstitial nephritis. Group 3 macroscopic lesions were limited to mottled, tan lungs in 2 of 2 boars, with 4%-9% of the lungs grossly affected by lesions. Microscopic lesions in boars from group 3 included mild, multifocal, nonsuppurative interstitial pneumonia in 2 of 2 boars and mild, multifocal, lymphocytic interstitial nephritis in 1 of 2 boars.
At DPI 48, no macroscopic lesions were present in boars from groups 1 or 2, and the boars in group 1 had no microscopic lesions. Microscopically, 1 of 2 boars in group 2 had mild lymphoid depletion and multifocal mineralization in the prostate gland. In group 3, 1 of 2 boars had mild lymphadenopathy, 2 of 2 had moderate nonsuppurative interstitial pneumonia, 2 of 2 had mild lymphoid depletion, 2 of 2 had mild, multifocal, lymphocytic interstitial nephritis, and 1 of 2 had accumulation of low numbers of macrophages within the testicular interstitium.
On DPI 90, the euthanized boar from group 1 had mild, multifocal, bronchiolar-associated lymphoid hyperplasia. Microscopically, 2 of 2 boars in group 2 had mild to moderate nonsuppurative interstitial pneumonia, 1 of 2 had mild lymphoid depletion, and 1 of 2 had mild bronchiolar-associated lymphoid hyperplasia. Macroscopic lesions in boars from group 3 included mild lymphadenopathy and mild interstitial pneumonia, with 6% of the lungs grossly affected by lesions in 1 of 2 boars. Microscopically, 2 of 2 boars in group 3 had mild, multifocal, nonsuppurative interstitial pneumonia, and 1 of 2 had mild lymphoid depletion.
Immunohistochemistry
Porcine circovirus-2 antigen was not detected in any of the control boars. Low amounts of PCV-2 antigen were detected in lymphoid tissues (lymph nodes, tonsil, and spleen) in 2 of 2 boars from group 2 and 2 of 2 boars from group 3 on DPI 23. One of 2 boars in groups 2 and 3 were PCV-2 IHC positive in lymph nodes and tonsil on DPI 48. No IHC signal was observed in lymphoid tissues of boars in groups 2 and 3 on DPI 90. Porcine circovirus-2 antigen was not detected in the testes, epididymis, seminal vesicles, bulbourethral gland, prostate, or penile tissue of necropsied boars in the present study.
Discussion
Currently, there is little information available in the peer-reviewed literature on shedding of PCV-2 in semen. With other swine viruses such as PRRSV, extensive testing of serum, semen, or blood swabs is done on a routine basis to monitor and decrease the risk of transmission. 10 , 33,34 . The objectives of the current study were to determine if there is a difference in shedding pattern or quantity of virus in semen of distinct PCV-2 genotypes and correlate the incidence and amount of PCV-2 within serum and blood swabs to that in semen.
In a previous PCV-2 experimental boar inoculation model, detection of PCV-2 DNA in semen was intermittent throughout the 47 days postinfection after intranasal inoculation. 21 In the current study, the results indicate that shedding of PCV-2 DNA in boar semen was continuous rather than intermittent. Differences in PCV-2 DNA shedding between studies could be due to the low number of inoculated boars in both experiments, route of infection, differences in assays used for PCV-2 detection, and differences in isolates used for inoculation. In the current study, the boars were inoculated intranasally and intramuscularly to ensure that all boars became infected at the same time and with a similar dose. This is not always possible by the intranasal route alone, as pigs tend to sneeze or breathe hard during the inoculation process. In addition, it has been determined that real-time PCR has slightly better detection limits compared to nPCR, 30 and there are differences in virulence among PCV-2 isolates. 26
To the authors' knowledge, this is the first study to evaluate the amount of PCV-2 DNA shed in semen of infected boars, and the results indicate that both PCV-2a and −2b DNA are shed in low quantities in raw semen (221-103,336 PCV-2 genomic copy numbers/ml). Moreover, it was demonstrated that the amount of PCV-2 in semen is lower than in serum.
The inocula used in the present study had an infectious titer of 104.3 TCID50 for PCV-2b versus 104.5 TCID50 for PCV-2a. Porcine circovirus-2b-inoculated boars became viremic 3–7 days after viremia started in the PCV-2a boars. This difference in time until detection of viremia onset could be due to the difference in amount of virus given at challenge; nevertheless, the inocula titer differences were minimal, and both groups shed PCV-2 DNA in semen at similar low quantities.
Length of PCV-2 shedding in semen varied among inoculated boars in the current study. In PCV-2a-inoculated boars (group 2), the narrowest incidence of detectable PCV-2 DNA was 1 collection point, with the longest incidence of detection being 11 consecutive collections (49 days). It should be noted that 2 boars within group 2 were shedding PCV-2 DNA in semen, but were randomly selected for necropsies on DPI 23 and 48; thus, it is unknown how long shedding would have persisted in these boars. In the PCV-2b-inoculated group (group 3), 1 boar never shed detectable amounts of PCV-2 DNA in semen. However, PCV-2 DNA in serum was first detected on DPI 16, and the boar was then euthanized on DPI 23 as predetermined by the experimental design. In all other inoculated boars, serum viremia preceded semen PCV-2 DNA shedding. The range was 4–14 days post-serum detection. In addition, the boar that did not shed detectable amounts of PCV-2 DNA in semen did, however, have detectable levels of PCV-2 DNA in tissues (lymph nodes, testes, epididymis, prostate, seminal vesicles, and bulbourethral gland) at necropsy. Only 1 boar in group 3 stopped shedding prior to being euthanized. All other boars had detectable PCV-2 DNA in semen at necropsy or until termination of the study. Results are supportive of similar observations in naturally infected boars, where extended shedding of PCV-2 DNA was observed in the semen of young Landrace boars. 22 In addition, the presence of PCV-2 DNA in semen had no affect on morphology, 22 which was further confirmed in the present study.
In the field, PCVAD is commonly associated with coinfections from other swine pathogens. 7 , 31 In experimental PCV-2 models, it has been confirmed that coinfections with Mycoplasma hyopneumoniae 28 PPV, 1 and PRRSV 2 , 13 , 36 increase the incidence and severity of PCV-2-associated lesions and disease. In the current study, rooms were cleaned daily, and there were no new animal introductions. These conditions are considerably different from commercial boar studs and breeding herds. It can be hypothesized that environmental factors, introduction of naïve animals to the population, and coinfections may exacerbate the length of PCV-2 shedding in semen, which would further support observation of up to 27.3 weeks of PCV-2 shedding in the semen of naturally infected boars. 22
It has been shown that serum PCR is more sensitive than semen PCR for the detection of PRRSV in experimentally challenged boars early in the disease phase. 33 Moreover, there was agreement between blood swab samples and serum samples when used for the detection of PRRSV RNA in the first 6 days postchallenge in boars. 34 In the current study, PCV-2 DNA was detected in serum prior to, and in higher amounts than, semen or blood swab samples. In the later course of infection, blood swab samples remained positive for PCV-2 DNA longer than semen and serum samples. One boar in Group 2 did not have detectable PCV-2 in serum after DPI 34, yet real-time qPCR detected PCV-2 DNA in blood swab samples at all collection points thereafter, including at termination of the study on DPI 90. Previously, in experimentally PCV-2-inoculated gilts, plasma viremia was observed up to 21 days postinoculation. However, peripheral blood mononuclear cells (PBMC) were positive to DPI 49 in the same animals. 32 Because blood swab samples contain erythrocytes, plasma, and PBMC, it is reasonable to conclude that blood swab samples in the current study were positive due to cell-associated PCV-2. It is possible that the lack of PCR-positive blood swab samples in the early stages of infection was influenced by the low numbers of boars used. Cell-associated PCV-2 infection may not necessarily be associated with viremia or PCV-2 DNA shedding in semen.
Nine of 12 PCV-2a- and PCV-2b-inoculated boars had detectable amounts of PCV-2 DNA in serum at necropsy. Two PCV-2a- and 1 PCV-2b-inoculated boars necropsied at DPI 90 were not viremic; however, blood swab samples and tissues (lymph nodes and sex glands) were positive for PCV-2 DNA. These results indicate and support that PCV-2 DNA is cell associated in the later stages of infection. However, it is unknown how long PCV-2 remains in the cell-associated state in the postviremic phase of infection.
Porcine circovirus-2 antigen was detected in sex glands of a naturally infected diseased boar. 25 In contrast, in the current study no boar was clinically affected, and PCV-2 antigen was not observed in sex glands of inoculated boars. Porcine circovirus-2 DNA levels in all sex glands ranged from 103.3 to 107.3 PCV-2 copy numbers per 0.25 g of tissue. In contrast, IHC staining for PCV-2 antigen was observed in lymphoid tissues at different necropsy points, and this correlated with a minimum of 107.5 PCV-2 DNA copies in 0.25 g of tissue. Previously, it was estimated that 10 8 genomic copies of PCV-2 were needed to detect viral antigen in tissues by IHC. 5 It should to be taken into account that the previously published PCR and IHC assays 5 and the PCR and IHC assays used in the present study 29 are different, which might explain the slight discrepancies in detection limits between studies.
In summary, the results of the present study suggest that both PCV-2a and −2b DNA were present in the semen and all sex glands of experimentally inoculated boars. The incidence of PCV-2 DNA in semen and length of shedding may vary among individual boars. Furthermore, the current study suggests that PCV-2 DNA shedding in semen may be continuous rather than intermittent postinfection. It remains to be determined if infectious PCV-2 is shed in semen and is a possible route of transmission and dissemination of PCV-2 among swine populations. Furthermore, the effect of using PCV-2-positive semen on herd-reproductive performance should be evaluated to make appropriate decisions on the amount of testing that is justifiable in boar studs.
Acknowledgements
Funding for this project was supported by the National Pork Board. The authors would like to thank Dr. Phil Gauger, Dr. Abby Patterson, Matt Boogerd, and Paul Thomas for their assistance with animal care and sample collection throughout the study. The authors would also like to thank Newsham Choice Genetics for the contribution of the boars.
Footnotes
a.
US BAG™ System, Minitube of America Inc., Verona, WI.
b.
Safe-T-Fill®, RAM Scientific Inc., Yonkers, NY.
c.
QuickVue® In-Line Strep A swabs, Quidel Corp., San Diego, CA.
d.
Falcon®, BD Biosciences, San Jose, CA.
e.
TriXcell, IMV Technologies USA, Maple Grove, MN.
f.
10% buffered formalin, Fisher Scientific Co., Chicago, IL.
g.
QIAamp® DNeasy® Blood & Tissue Kit, Qiagen Inc., Valencia, CA.
h.
SAS® version 9.1.3, SAS Institute Inc., Cary, NC.