
Abstract
Organophosphorous pesticides (OPs), with high efficiency, broad-spectrum and low residue, are widely used in China. Omethoate is a broad category of organophosphorous pesticides and is more domestically utilized which has chronic toxic effect on human health caused by long-term, low-dose exposure to Ops, recently its potential genotoxicity has attracted wide attention which can cause chromosomal DNA damage. Thus, the aim of this study is screen susceptible biomarkers and explore the mechanism of canceration induced by omethoate. 180 long-term organophosphorus pesticide–exposed workers and 115 healthy controls were recruited. Quantitative polymerase chain reaction method was applied to determine the relative telomere length in peripheral lymphocyte DNA as well as p53 and p21 gene expression levels. Genetic polymorphisms were determined by the polymerase chain reaction–restriction fragment length polymorphism method. Multiple linear regression was conducted to explore the effects of exposure, expression levels, and polymorphisms in genes on the telomere length. The results showed the relative telomere lengths in the exposure group were significantly longer than that in the control group. The messenger RNA expression levels of p53 and p21 in exposure group were significantly lower than that in the control group; telomere lengths of the CA genotype individuals of p21 rs1801270 polymorphism locus were significantly longer than that of the CC genotype in the control group that were estimated using the Bonferroni method; and bivariate correlation analysis showed that the messenger RNA expression level of gene p53 was negatively correlated with telomere length, and the messenger RNA expression level of gene p21 was positively correlated with telomere length. Multivariate analysis found that p53 messenger RNA and p21 messenger RNA had an impact on telomere length. These results demonstrated that the messenger RNA expression levels of p53 and p21 may have a relationship with the changes in telomere length induced by omethoate and provided strong evidence for the mechanism of canceration induced by poison.
Introduction
Omethoate belongs to a broad category of organophosphorus pesticides (OPs) and is more domestically utilized; it is an organophosphorus insecticide and acaricide which has highly toxic, highly effective, and broad-spectrum features. The acute toxicity mechanisms of OPs are mainly by inhibiting cholinesterase (ChE) activity which results in the accumulation of acetylcholine in vivo, thus stimulating muscarinic and nicotinic receptors to cause neurological disorders.1,2
In recent years, acute poisoning incidents caused by OPs have been reduced, while the chronic toxic effect on human health caused by long-term, low-dose exposure to OPs attracted people’s attention gradually. More and more studies have concluded that long-term, low-dose exposure to OPs (mainly methyl parathion, methamidophos, omethoate, and diazinon) had a close relationship with human tumorigenesis, adverse reproductive outcomes, and neurological and neurobehavioral function abnormalities.3–5 Previous studies on genetic damage caused by OPs mainly included comet assay, micronucleus (MN), and sister chromatid exchange (SCE).6–11 The research involving damage to telomeres was rarely reported. DNA damage is an important molecular biological event which results in malignant transformation, because telomere DNA is located at the end of the chromosome and is more susceptible to be attacked by exogenous compounds. Telomeres are specialized DNA–protein functional complexes which are constituted of non-coding DNA repeating sequences and telomere-binding proteins (TBPs) at the ends of the linear chromosomes of the eukaryotic cell, its dysfunction caused by the telomere excessive consumption or structural disruption may result in cell carcinogenesis. 12 When DNA is damaged, cell division will stagnate in the G1 phase of mitosis, so the checkpoint is extremely important in this phase. p53 is a tumor suppressor gene which is involved in the G1–S checkpoint, has the function of gene guards. The p21 gene is located in the downstream position of p53 gene in the signaling pathway and is a cyclin-dependent kinase inhibitor (CDKI), p21 and p53 constitute the G1–S checkpoints, so damaged cells stagnate in the G1 phase to reduce the formation of mutations, and thus play a tumor suppressor effect. At the same time, the p53 expression caused by DNA damage is regulated negatively by the MDM2 gene; MDM2 is an important regulator of p53, has the function of degrading p53, and also participates in tumor growth and metastasis. 13 Currently, there are a large number of polymorphic loci within these genes. The polymorphism can cause the changes in the encoded products conformation, the enzyme activity, and the expression levels of encoded products. These changes may lead to different abilities of repairing damaged DNA, thus causing different susceptibility to diseases among individuals. Studies have shown that the p53 gene rs1042522, rs17878362, and rs1625895 and the p21 gene rs1801270 and rs1059234 have a close relationship with cancer risk.14–20 MDM2 gene rs3730485 was also intimately related to tumor. 21 So far, the correlation between the polymorphisms and the messenger RNA (mRNA) expression levels of the p53, p21, and MDM2 genes and telomere length has not been explored.
Therefore, we studied the genetic polymorphisms and mRNA expression of the above three cell cycle regulation genes and explored the relationships with telomere length to screen susceptible biomarkers and explore the mechanism of canceration induced by poison. This study provides the basis for OP toxicity research and biological monitoring of occupational exposure and protects the health of the occupational population.
Materials and methods
Study population
In total, 180 workers exposed to omethoate for longer than 8 years were recruited as the exposure group. In addition, 115 unexposed controls were recruited who lived in the same city but without a history of exposure to OPs or other toxicants. Written informed consent was obtained from each subject, in order to collect occupational history, basic situation, and biological samples.
Research methods
The detection of relative telomere length
The relative telomere length was determined using genomic DNA isolated from peripheral blood lymphocytes and real-time polymerase chain reaction (PCR) method provided by Cawthon. 22 Real-time PCR was performed for each sample using the reference primers and telomere primers, and each sample was run using three parallels; qPCR SYBR Green Master Mix was purchased from Vazyme Biotech, and the primers were synthesized by Beijing Invitrogen Corporation. The software for ABI 7500 Fast Real-Time Quantitative PCR instrument can calculate Ct values of the samples directly. Relative telomere length was calculated using the formula of 2−ΔΔCt (ΔCt = CtTelomere − CtReference; ΔΔCt = ΔCt − ΔCtaverage normal controls).
The detection of genetic polymorphisms
Six polymorphic loci of p53 rs1042522, p53 rs17878362, p53 rs1625895, p21 rs1801270, p21 rs1059234, and MDM2 rs3730485 from three genes were detected using genomic DNA isolated from peripheral blood lymphocytes. Among them, the deletion in p53 rs17878362 and MDM2 rs3730485 can be detected by PCR electrophoresis directly; the PCR–restriction fragment length polymorphism (PCR-RFLP) method detected other loci for genotyping, primer sequences, and restriction endonucleases as described previously.23–27
The PCR was carried out in a volume of 15 μL containing 50 ng of DNA, 7.5 μL of 2× PCR master Mix (Shanghai Lifefeng Biotech Co., Ltd), and 3 pmol of each primer. PCR reaction conditions were as follows: first, incubated at 94°C for 5 min, then entered into the cycling program of denaturation–annealing–extension to select a different annealing temperature depending on the Tm value and pre-experimental results, followed by a final extension at 72°C for 10 min. PCR products were electrophoresed on 2% to 4% agarose gels and visualized with ethidium bromide staining and ultraviolet illumination to determine the success of PCR.
Enzyme digestion was conducted in a final volume of 20 μL using 5U enzyme (Thermo Fisher Scientific) and 10 μL of PCR product was obtained. Enzyme reaction was carried out according to the enzyme reaction temperature for more than 4 h or overnight. The digested products were visualized using gel imaging analyzer after electrophoresis.
Detection of gene expression level
Total RNA in peripheral blood leukocytes was extracted using chloroform and isopropyl alcohol extraction method and quantified by measurement of the absorbance at 260 nm; purity was assessed from the 260/280 nm absorbance ratio; using reverse transcription kit reversed mRNA to complementary DNA (cDNA) for frozen spare (TaKaRa). Real-time PCR was used to determine the relative expression levels of p53 and p21 genes. 13
Double-distilled water (ddH2O) was used as a negative control and three parallel samples were done for each sample. The difference among the Ct value of three parallels should be less than or equal to 0.5; if more than 0.5, the sample should be redone. Negative control was used in each round of PCR. The PCR was carried out in a volume of 20 μL containing 1.0 µL of cDNA, 10 µL of 2× AceQ qPCR SYBR Green Master Mix (Vazyme Biotech), and 3 pmol of each primer (Sangon Biotech). Reaction conditions were the following: predegeneration at 95°C for 5 min, and then temperature cycles were set at 95°C for 30 s, 60°C for 30 s for 30 cycles. After the end of each reaction, the amplified products were analyzed according to the melting curves, and the parameters set were as follows: 95°C for 15 s, 60°C for 1 min, and 95°C for 15 s.
The software for Stratagene Mx3000P real-time quantitative PCR instrument can calculate Ct values of the samples directly. The relative gene expression was calculated using the formula of 2−ΔΔCt (ΔCt = CtGene − CtGAPDH; ΔΔCt = ΔCt − ΔCtaverage normal controls).
Statistical analysis
The data were analyzed using SPSS 21.0 software (SPSS Inc., Chicago, IL, USA). Rank sum test was performed to compare the relative telomere length and mRNA expression between omethoate-exposed workers and healthy controls. Genotypes and allele frequencies were calculated by direct counting and were tested for Hardy–Weinberg equilibrium. Effects of genetic polymorphisms on the relative telomere length were analyzed using Rank sum test, and the Bonferroni method was used to do the comparisons between the two groups. The spearman rank correlation was used to analyze the correlation for non-normal distribution data. Multiple linear regression was conducted to analyze the influencing factors of telomere length. All statistical tests were two sided, and the level of statistical significance was set at α = 0.05.
Results
Population-based data
The compared results of gender, age, smoking, and drinking history between the two groups are shown in Table 1. The differences had statistical significance in the gender, age, smoking, and drinking characteristics between the two groups.
General characteristics of exposure and control groups.

The detecting results of external exposure
The assessment of the occupational exposure level in the research workers was from the results of the detecting reports of a pesticide plant from 2011 to 2013. The 8 h time-weighted average concentration and short-term exposure concentrations were not exceeded in the omethoate packaging workshops of the pesticide plant, which is lower than occupational exposure limits in the national occupational health standards. The concentration of each type of work was not beyond the occupational exposure limit prescribed by the state.
Relative telomere length in two groups
After statistical analysis, we found that relative telomere lengths in the exposed group (1.50 (1.11, 2.57)) were significantly longer than that of the normal controls (0.94 (0.76, 1.32)), and the difference was statistically significant (Z = 7.910, p < 0.001).
Effects of gender, age, smoking, drinking, and working duration on telomere length
Some studies suggested that age had an impact on the telomere length;28,29 therefore, we analyzed the correlation between age and telomere length and found that there was no correlation in the exposure group and control group (rs = 0.049, p = 0.517; rs = 0.116, p = 0.218).The results in Table 2 show that gender, age, smoking, drinking, and working duration had no effect on the relative telomere length in the exposure or control group (p > 0.05). In the same stratification of gender, age, smoking, and drinking, the relative telomere length had a statistically significant difference between the two groups (p < 0.05). According to the segments of Chinese age, we regarded 40 years old as the boundary and divided age into the low-age group and the high-age group.
The effects of sex, age, smoking, drinking, and working duration on telomere length.

p* indicates the comparison of telomere length between exposure group and control group after stratifying.
p# represents the comparison among the layers after stratifying.
Effects of genetic polymorphisms on telomere length
The genotype distribution for each genetic polymorphism locus did not deviate from the Hardy–Weinberg balance (p > 0.05), suggesting that the control samples had representativeness. We regarded the wild homozygous genotype as a reference to analyze the differences of telomere length between heterozygous or mutant homozygous genotype and wild homozygous genotype. Among them, the wild homozygous genotype of rs1625895 locus and mutation homozygous genotype of rs17878362 locus for cell cycle regulation gene p53 were only one case in the exposure group; therefore, they would merge with the heterozygous genotype directly. The relationships between genetic polymorphisms and telomere length are shown in Table 3. The results show that telomere length of the CA genotype for p21 rs1801270 polymorphism locus was significantly longer than that of the CC genotype in the control group, the difference was statistically significant (p = 0.008); however, there were no statistically significant differences among the genotypes of other loci (p > 0.05).
The relationships between genetic polymorphisms and telomere lengths.

SNP: single nucleotide polymorphism.
p*: To compare telomere lengths among genotypes using a rank test of k independent samples.
p**: The compared results between the two groups, the Bonferroni method was adopted.
Ref.*: The reference group of comparing different genotypes.
Gene mRNA expression levels in different groups
After the statistical analysis, we found that the mR
The relationship between mRNA expression levels and telomere length
After the bivariate correlation analysis, we found that the mRNA expression of gene p53 was negatively correlated with telomere length (rs = −0.735, p < 0.001), and the mRNA expression of gene p21 was positively correlated with telomere length (rs = 0.181, p = 0.002).
The relationship between genetic polymorphism and mRNA expression level
Table 4 shows the difference in mRNA expression levels classified as per the genotype of single nucleotide polymorphism (SNPs) and exposure. We merged the wild homozygous genotype of p53 rs1625895 locus and mutation homozygous genotype of p53 rs17878362 locus and then compared it with the heterozygous genotype for only one case in the exposure group. The results were that the mRNA expression levels of each genotype individuals of p53 rs1042522, p53 rs17878362, p53 rs1625895, p21 rs1801270, and p21 rs1059234 loci were lower than those in the control group, and the differences were significant (p < 0.05). The mRNA expression levels had no statistically significant differences among the different genotypes in the exposure group or control group.
The relationship between p53 and p21 genetic polymorphisms and mRNA expression levels.
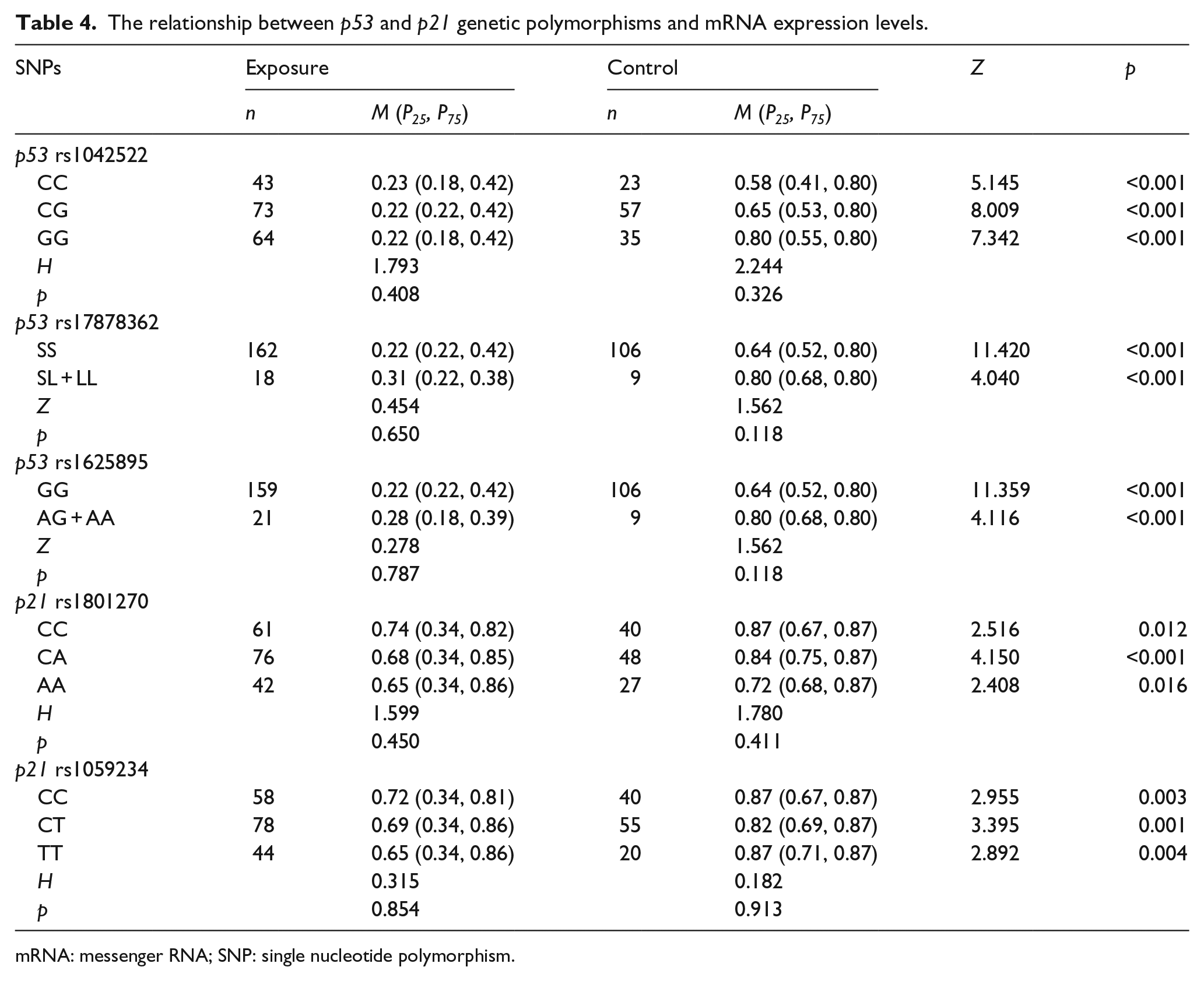
mRNA: messenger RNA; SNP: single nucleotide polymorphism.
Influencing factors of telomere length in multiple linear regression model
A multiple linear regression equation was fitted to screen the influencing factors. Because the data type of telomere length was an abnormal distribution, the data were logarithmically transformed to meet the conditions of multiple linear regression analysis. ln was established (telomere length) as the dependent variable. The independent variables included group (0 represents control,1 represents exposure), gender (female = 0, male = 1), smoking (non-smoking = 0, smoking = 1), drinking (non-drinking = 0, drinking = 1), working duration (control = 0, exposure <15 = 1, 15–30 = 2, and >30 = 3), and genetic polymorphisms loci (considering the combined genotype as a standard and wild homozygous genotype as a reference; p53 rs1042522: CC = 0, CG + GG = 1; p53 rs17878362: SS = 0, SL + LL = 1; p53 rs1625895: AA = 0, AG + GG = 1; p21 rs1801270: CC = 0, AC + AA = 1; p21 rs1059234: CC = 0, CT + TT = 1; and MDM2 rs3730485: SS = 0, SL + LL = 1) and the independent variables were screened using a stepwise method. The variables kept in the models included p53 mRNA (b = −2.057, p < 0.001) and p21 mRNA (b = 0.645, p < 0.001). No found that group, gender, alcohol, smoking, age, working duration, and genetic polymorphisms were included in the model. The results are shown in Table 5.
Independent variables of entering the regression model.

mRNA: messenger RNA.
Discussion
Telomeres are specialized DNA–protein structures located at the ends of the linear chromosomes. In mammalian cells, they are composed of multifold hexameric TTAGGG repeats and a number of associated proteins. Their role is to protect the integrity and stability of the chromosome.12,30 At present, a large number of studies have suggested that telomeres and telomerase abnormalities are related to tumor formation; however, Professor Carol Greider has proposed that telomerase was not able to help explain why telomeres have a certain length and how to extend the length of telomeres, which will bring a huge impact on human health. Although telomere shortening could trigger the body’s aging and a variety of diseases, longer telomeres are often directly related to cancer. 31
At present, the research results on the changes in telomere length caused by poisons were different; most of the studies suggested that different toxicants could cause telomere shortening, Wu etal. 32 explored the influencing factors of peripheral white blood cell telomere length in Chinese battery manufacturing plant workers and found that telomere length in 144 battery plant workers was significantly shorter than that of normal ones. In addition, telomere length in lead-exposed workers was in negative correlations with the lead levels in blood and urine, working length, and body mass index. Li et al. 33 detected the telomere length of 157 workers working in the rubber industry in Sweden. N-nitrosamines exposure may lead to telomere shortening, but there were no strong associations between other exposures and telomere length. El-Zein et al. 34 suggested that exposure to ethylene glycol monomethyl ether (EGME) in utero could result in terminal chromosome rearrangements and shortening of telomere length, leading to observed dysmorphic features and idiopathic mental retardation. However, studies have found that toxicants could extend telomere length. Shin et al. 35 explored the relationship between low-dose exposure to persistent organic pollutants (POPs) and telomere length and found that telomere length was increasing across low doses of exposure to POPs in which the majority of study subjects were found, suggesting that low-dose POPs may act as a tumor promoter in carcinogenesis in humans. Dioni et al. 36 studied the effects of short-term exposure to inhalable particulate matter (PM) on telomere length, telomerase expression, and telomerase methylation in steel workers and found that short-term exposure to PM caused a rapid increase in blood leukocyte telomere length.
Although shortened telomeres have been found in many studies, elongated telomere length has been observed as an early response after low-dose treatment with various chemical carcinogens in vitro and animal experiments, suggesting that low-dose exposure to carcinogenic chemicals may function as a tumor promoter at the very early stage of carcinogenesis in humans. 35 The results of this study also found that relative telomere length in an omethoate-exposed population was longer than that of the control, and the differences had statistical significance (Z = −7.910, p < 0.001); in addition, the results showed that the relative telomere length decreased with the extension of working duration in the omethoate-exposed group. The assessment of occupational exposure level in the research workers was from the results of the detecting reports of a pesticide plant from 2011 to 2013. The 8 h time-weighted average concentration and short-term exposure concentrations were not exceeded in the omethoate packaging workshops of the pesticide plant, which is lower than occupational exposure limits in the national occupational health standards. Our study found that omethoate in low-dose exposure caused the extension of telomere length.
As almost epidemiological studies have been performed to investigate whether short telomere length is associated with unhealthy health behaviors and known risk factors for various diseases or short telomere length at baseline predicts future risk of diseases or death.37,38 Also, shortened telomeres have been found in many cancers although telomerase is activated in most cancers to prevent the telomeres from getting even shorter. 39 From this viewpoint, the positive associations between low-dose omethoate and telomere length could be awkward to explain because omethoate was associated with increased risk of various diseases in general population.3–5 However, telomere length may have different implications depending on the stage of carcinogenesis. Telomeres generally shorten with age, so the occurrence of telomere lengthening should also be viewed as possibly adverse. Considering previous experimental studies on carcinogenic chemicals, 40 telomere lengthening due to the exposure to carcinogenic chemicals such as omethoate may be a very early phenomenon in carcinogenesis. In this sense, when our current findings are evaluated, it is critical to consider that our study was cross-sectional and performed among healthy subjects and that our comments on relations to carcinogenesis are therefore speculations that should be pursued in other research. Therefore, the positive associations between low-dose omethoate and telomere length observed in this study are worthy of note.
This study also analyzed the effects of gender, age, smoking, and drinking on telomere length. Considering the influence of age on telomere length, we analyzed the correlation between age and telomere length and found that there were no correlations between the exposure group and control group (rs = 0.049, p = 0.517; rs = 0.116, p = 0.218). Single factor analysis found that gender, age, smoking, drinking, and working duration had no effect on the relative telomere length in the exposure or control group (p > 0.05). Multi-factor analysis has not found that factors such as gender, age, smoking, drinking, and working duration had an effect on telomere length (p > 0.05), which was consistent with that of single factor analysis.
Genetic injury is a result of the chronic toxic effect of organophosphorus pesticide on the exposure-crowd. The research in this area is mainly concentrated on the micronucleus test and comet assay. Benitez-Trinidad et al. 41 found that temephos could increase the micronucleus frequency and cause DNA damage in human lymphocytes and hepatoma cells (HepG2). Timoroglu et al. 42 indicated that Phorate (PHR) and Trichlorfon (TCF) in organophosphorus insecticides possessed clastogenic, mutagenic, and DNA damaging effects in human lymphocytes in vitro. Moreover, it was also reported that chromosome aberration frequency and micronucleus rate increased in the farmers who were occupationally exposed to pesticides during spraying activities. 43 Hreljac and Filipic 44 found that the combined function of organophosphorus pesticides and benzopyrene can increase the incidence of tail moment in the comet assay. However, these genetic damages are often accompanied by changes in gene expression levels, and individual gene polymorphisms may lead to different abilities of repairing damage DNA, thus causing different susceptibility to diseases among individuals.
Damaged DNA is to be repaired by various types of DNA damage repair proteins to maintain the stability of the genome. Cell can give sufficient time to damaged DNA for repairing through cell cycle arrest. DNA damage can induce p53 expression, p53 can prompt the cell cycle arrest by inducing the p21 protein expression, which can directly induce the damaged cells to enter into G1 phase to repair; In addition, p53 can also be inactivated by the interaction of cell gene products, such as MDM2 can inactivate p53 by combining with it; in some common human sarcoma, MDM2 gene amplification may interfere the normal function of p53 gene. If the p53 or p21 gene changes, they may lead to the functional changes in promoting cell cycle arrest, thus the damaged DNA will not have enough time to repair and the parent cells will pass damaged DNA information to the progeny cells, this causes the genome instability leading to cell malignancy.45,46
Therefore, this study analyzed the impact of p53, p21, and MDM2 cell cycle regulation genetic polymorphisms on telomere length. The results showed that telomere length of the CA genotype for p21 rs1801270 polymorphism locus was significantly longer than that of the CC genotype in the control group, and difference had statistical significance (p = 0.008), and there were no statistically significant differences among the genotypes of other loci (p > 0.05). Vargas-Torres et al. 18 studied p21 gene polymorphisms in 283 cases of cervical cancer patients and 189 cases of disease-free people, the results showed that homozygous and heterozygous mutant genotypes of the above two polymorphism loci were the risk factors for cervical cancer and its results had inconsistent places with this study.
The MDM2 gene is a target gene of p53 gene downstream regulation and is an important regulator of p53 gene. It participates in tumor growth and metastasis which is highly expressed in tumor tissues. Studies have shown that 40 bp missing genotype (SS) for MDM2 rs3730485 locus may be a risk factor for uterine fibroids. 47 Another study also suggested that the SS type of this locus was a risk factor of hepatocellular carcinoma. 48 Our study found that telomere length had no statistically significant differences among the different genotypes of MDM2 rs3730485.
p53 and p21 are the important tumor suppressor genes that play an important role in the development of cancer. To detect the relative mRNA expression levels of cell cycle regulation genes for the research objects, we found that the mRNA expression levels of the p53 and p21 genes in the exposure group were lower than those in the control group (p < 0.05), suggesting that the ability of cell damage repair has decreased in the exposed group, which were not consistent with previous studies.23,49 The core of the cell cycle is mainly regulated by cyclin-dependent kinases (CDKs), and the key to CDKs regulation is the cell cycle monitoring point; CDKs activation is controlled by the positive and negative regulation of cyclin and CDKIs. 50 When the CDK combines with cyclin, CDKs show kinase activity; cyclin D is expressed in G1 phase, in which cyclin D1 plays a key role in cell G1/S phase, which is the initiation factor of cell cycle, its expression has a close relationship with the formation of tumors in the early G1 phase. In G1 phase, cyclin D binds to and activates CDK4/6, which further phosphorylates the downstream proteins. In the G–S phase, cyclin E binds to CDK2 and promotes cell entry into the S phase through the G1/S restriction point. 51 However, The p21 gene is located in the downstream position of p53 gene in the signaling pathway and is a CDKI which makes damaged cells stagnate in the G1 phase to reduce the formation of mutations.
Multivariate analysis found that the mRNA expression level of p53 gene was negatively correlated with telomere length (b = −2.057, p < 0.001), and the mRNA expression level of p21 gene was positively correlated with telomere length (b = 0.645, p < 0.001), which were consistent with the results of bivariate correlation analysis. The decreased expression levels of the two genes led to a decline in the ability of cell damage repair which causes the genome instability leading to cell malignancy, malignant cells might have extended the telomere length for prolonging the life of their own. This study also analyzed the relationships between the genotypes of p53 genetic polymorphisms and their mRNA expression levels; mRNA expression levels had no statistically significant differences among the different genotypes in the exposure group or control group. For analyzing the relationship between the genotypes of p21 genetic polymorphisms and their mRNA expression levels, we also found no connections between them.
Conclusion
In conclusion, the relative telomere length became longer in peripheral blood leukocyte DNA of the omethoate-exposed workers. The length was associated with the mRNA expression levels of p53 and p21 gene. This study found that telomere length of the CA genotype for p21 rs1801270 polymorphism locus was significantly longer than that of the CC genotype in the control group for the first time. The research results provide strong evidence for the mechanism of telomere damage induced by poison and for screening effective susceptible biomarkers.
Footnotes
Acknowledgements
The authors express their gratitude to all the individuals who volunteered to participate in this study.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Program for Science and Technology Development of Zhengzhou (131PPTGG376) and the Outstanding Youth Grant of Zhengzhou University (1521329035).