
Abstract
Laryngeal cancer is one of the most common fatal cancers among head and neck carcinomas, whose mechanism, however, remains unclear. The proneural basic-helix-loop-helix protein achaete-scute complex homologue-1, a member of the basic helix-loop-helix family, plays a very important role in many cancers. This study aims to explore the clinical value and mechanism of achaete-scute complex homologue-1 in laryngeal cancer. Methods including Cell Counting Kit-8, flow cytometry, Transwell invasion assays, and scratch assay were adopted to further explore the bio-function of achaete-scute complex homologue-1, whose expression was examined in fresh and paraffin chip of laryngeal carcinoma tissues by means of western blot and immunohistochemistry, after the interference of achaete-scute complex homologue-1; achaete-scute complex homologue-1, an overexpression in laryngeal carcinoma whose carcinogenicity potential was confirmed via western blot, was correlative with T classification (p = 0.002), histological differentiation (p = 0.000), lymph node metastasis (p = 0.000), and poor survival (p = 0.000). Multivariate analysis shows that achaete-scute complex homologue-1 overexpression is an independent prognostic factor unfavorable to laryngeal carcinoma patients (p = 0.000). Moreover, knocking down achaete-scute complex homologue-1 expression could significantly suppress the proliferation, migration, and invasion of laryngeal carcinoma cell in vitro and disorder epithelial–mesenchymal transformation–associated protein expression. Achaete-scute complex homologue-1 plays an important role in the genesis and progression of laryngeal carcinoma and may act as a potential biomarker for therapeutic target and prognostic prediction.
Keywords
Introduction
Among head and neck carcinomas, laryngeal carcinoma is a kind of solid tumor with high incidence and mortality. 1 In recent years, all newly diagnosed malignancies of laryngeal carcinoma have accounted for approximately 2.4% all over the world every year. 2 Although diagnosis and treatment of laryngeal carcinoma have made huge progress, the survival rate of patients with laryngeal carcinoma has failed to see dramatic improvement over the past two decades. 3 It is therefore important to find out the molecular mechanisms of the expansion and metastasis of laryngeal carcinoma, which will provide some new diagnostic and therapeutic targets for this disease and improve the prognosis of laryngeal carcinoma patients.
As a member of the basic helix-loop-helix (bHLH) transcription factor family, achaete-scute complex homologue-1 (ASCL1) is one of the most ubiquitous expressions ranging from plants to animals and activates transcription by being bonded to the E-box (5′-CANNTG-3′) in the promoter region. 4 It plays an important role in a series of cellular processes such as intracellular transportation and cell division. 5 Some studies indicated that the abnormal expression and function of ASCL1 protein are crucial to the occurrence of human cancers6–12 of many kinds, in which increased ASCL1 expression levels have been detected, and silencing ASCL1 expression could suppress the proliferation of both cancer6,7,10 and neuroendocrine tumor.5,13,14 And some report that suppression of ASCL1 expression by RNA interference could lead to cell-cycle procession as well as cell differentiation and apoptosis and inhibit growth in vitro in an ASCL1 expression–dependent manner.11,15,16 It was found that ASCL1 is also involved in the migration of cancer cells in various carcinoma cell lines such as oral squamous cell carcinoma line 17 and lung cancer cell lines. 11 These findings proved the involvement of ASCL1 in carcinogenesis. The significance of ASCL1 and its mechanisms of ASCL1 in laryngeal cancer, however, are still unknown. In this study, we find that ASCL1 can promote the proliferation and migration of laryngeal cancer by combining with DKK1 to facilitate the development of laryngeal cancer occurs.
Materials and methods
Patients and tissue samples
In all, 98 laryngeal cancer samples and 28 paired adjacent normal laryngeal tissue samples were collected from the Department of Otolaryngology/Head and Neck Surgery of the Affiliated Hospital of Nantong University. Each tumor was classified according to its location and histology by referring to the latest WHO classification 18 and tumor–node–metastasis (TNM) classification (Union Internationale Contre le Cancer (UICC). 19 Table 1 gives a clear description of the primary clinical and pathological features of all laryngeal patients. Among the 98 patients whose age ranges from 2 to 82 years (52.16 years on average), 96 males and 2 females, 28 were supraglottic and 70 were glottic. Histological differentiation was classified into high differentiation (grade I; n = 48), middle differentiation (grade II; n = 27), and low differentiation (grade III; n = 23). The overall survival time for the 98 patients after operation ranges from 1 to 120.7 months (median, 72 months). Another 8 fresh laryngeal cancer tissues and their surrounding normal mucosa tissues were collected as controls, and none of the samples were taken during postoperative adjuvant therapy. Follow-up information concerning all these samples was completely documented until May 2010.
ASCL1 expression and clinicopathological parameters in 98 laryngeal SCC specimens.

SCC: squamous cell carcinoma.
More relatively significant.
Most relatively significant.
Immunohistochemistry study of clinical samples
The tissues were cut into sections with a diameter of 1 µm, placed on 3-aminopropyltriethoxysilane (APES)-preliminary treatment slides for dewaxing, rehydrated through graded alcohol, and quenched in 3% hydrogen peroxide. Antigen retrieval was carried out through microwave heating at high power (750 W) in 10 mM sodium citrate buffer (pH 6.0) for three cycles, each cycle taking 5 min. After blocking them with normal serum for 1 h at room temperature, the sections were incubated with anti-human ASCL1 mouse polyclonal antibody (diluted 1:100; R&D; Zymed Laboratories, Inc., San Francisco, CA, USA) overnight at a temperature of 4°C, which is also the secondary antibody. Negative control slides were also handled in the same way using a heterogenetic antigen IgG (Sigma Chemical Co., St. Louis, MO, USA) of the same concentration as the primary antibody. All slides were processed using the anti-peroxidase method (Dako, Hamburg, Germany), with diaminobenzidine used for the final chromogen and Gill’s hematoxylin for counter staining.
The assessment of ASCL1 was carried out individually by two pathologists in a double-blind manner. 20 The ASCL1 immunostaining score was calculated including both the percentage of positively stained tumor cells and their staining intensity. The positive percentage of ASCL1 was scored as follows: 0 (tumor cells’ positive percentage ≤10%), 1 (tumor cells’ positive percentage 10%–30%), 2 (tumor cells’ positive percentage 30%–50%), 3 (tumor cells’ positive percentage 50%–70%), and 4 (tumor cells’ positive percentage >70%). And the staining intensity of ASCL1 was scored as “0” (unstained), “1” (lowly stained), “2” (moderately stained), and “3” (highly stained). The immunostaining score ranging from 0 to 12 was calculated as the positive score × the staining intensity score; 0–4 was considered as weak, while 5–12 was deemed as overexpression. 21
Western blot analysis
Tissues and cells were collected and lysed in radioimmunoprecipitation assay buffer, which contains 50 mM Tris-HCl (pH 8.0), 150 mM sodium chloride, 1.0% NP-40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, and 1 mM phenylmethylsulfonyl fluoride, and then centrifuged at 10,000g for 30 min to collect the supernatant, which was then diluted in 2× sodium dodecyl sulfate (SDS) loading buffer and boiled. Protein content in the cell lysates was measured using the Bio-Rad protein assay (Bio-Rad Laboratories, Hercules, CA, USA). Proteins were separated with SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride (PVDF) filter membranes (Millipore, Bedford, MA, USA), which were sealed with 8% dried skim milk in phosphate-buffered saline with Tween 20 (PBST; 20 mM Tris, 150 mM NaCl, and 0.05% Tween 20). After exposing at room temperature for 2 h, the membranes were incubated overnight with polyclonal antibody using the primary antibodies. Antibodies used were as follows: anti-ASCL1 (1:3000; Santa Cruz Biotechnology), anti-p27Kip1 (1:1000; Santa Cruz Biotechnology), anti-CDK2 (1:1000; Santa Cruz Biotechnology), anti-cyclin A (1:1000; Santa Cruz Biotechnology), anti-DKK1 (1:1000; Santa Cruz Biotechnology), anti-E-cadherin (1:1000; Santa Cruz Biotechnology), anti-N-cadherin (1:1000; Santa Cruz Biotechnology), anti-matrix metalloproteinase-3 (MMP-3; 1:1000; Santa Cruz Biotechnology), anti-matrix metalloproteinase-9 (MMP-9; 1:1000; Santa Cruz Biotechnology), anti-ZO-1 (1:1000; Santa Cruz Biotechnology, CA, USA), and anti–glyceraldehyde 3-phosphate dehydrogenase (GAPDH; 1:1000; Sigma Chemical Co., St. Louis, MO, USA). Then, horseradish peroxidase–linked IgG was used as the secondary antibody. Immunoreactive bands were visualized by chemiluminescence (NEN Life Science Products, Boston, MA, USA). After exposing the chemiluminescence to X-ray films, the films were scanned using a Molecular Dynamics Densitometer (Imaging Technology, London, ON, Canada) to collect values that are responsible for at least three independent reactions.
Plasmid constructs small interfering RNA and transfection
The small interfering RNA (siRNA) species used for knocking down ASCL1 were synthesized by Biomics (Nantong, China), and the siRNA sequences were as follows: ASCL1-siRNA1: 5′-TCGACTTCACCAACTGGTT-3′; ASCL1-siRNA2: 5′-ACTACTCCAACGACTTGAA-3′; ASCL1-siRNA3: 5′-GCCAACAAGAAGATGAGTA-3′; ASCL1-siRNA4: 5′-CGAACTGATGCGCTGCAAA-3′. The ASCL1 inhibitors and negative control molecules, synthesized by GeneChem Co., Ltd. (Shanghai, China), were added to culture media at a final concentration of 100 nM and were transfected into cells using Lipofectamine 2000 (Invitrogen Life Technologies, USA) according to the manufacturer’s protocols.
Cell lines and cell cultures
Laryngeal cancer cell lines Tu212 and Hep2 were obtained from BioHermes Co. (Wuxi, China) and were cultured in RIPA 1640 (Invitrogen, Carlsbad, CA, USA) which was supplemented with 10% fetal bovine serum (FBS; HyClone Laboratories, Logan, UT, USA), penicillin 100 U/mL, and streptomycin 100 µg/mL in 5% CO2 at 37°C.
Cell viability analysis
The viability of the TU212 cell line was assessed by performing the assay with Cell Counting Kit-8 (CCK-8; Dojindo, Kumamoto, Japan) according to the manufacturer’s instructions. The absorbance at 490 nm was measured by spectrophotometry with a microplate reader (model 680; Bio-Rad Laboratories, Hercules, CA, USA). Assays were repeated at least three times during separate experiments.
Cell-cycle analysis
TU212 cells were transfected with 20 mM ASCL1-siRNA1 or the control ASCL1-siRNA. After 48 h, the cells were harvested and placed in 70% ice-cold ethanol at a temperature of −20°C for 24 h and incubated with propidium iodide (20 µg/mL) and RNase A (100 µg/mL) in an incubator for 1 h at a temperature of 37°C. Assessment of cells containing propidium iodide–stained DNA was done using FACSCalibur flow cytometer (BD Biosciences, Sydney, Australia), and analysis of the percentage of cells in each phase of the cell cycle was made using a flow cytometer (Beckman Coulter Inc., CA, USA).
Apoptosis assay
The apoptosis assay was carried out at 48 h after the cells were transfected with 20 mM ASCL1-siRNA1 or the control ASCL1-siRNA. According to the operation instructions, the TU212 cells transfected with ASCL1-siRNA1 or the control ASCL1-siRNA were washed three times with ice-cold PBS and resuspended in 100 µL of 1× binding buffer and incubated with Annexin V-fluorescein isothiocyanate (FITC; BestBio, Shanghai, China) for 15 min at a temperature of 4°C in the dark. After being colored, the cells were incubated with propidium iodide for 5 min at a temperature of 4°C in the dark and were then analyzed using a flow cytometer (Beckman Coulter Inc., USA).
Cell migratory assays
The capability of cell migration was examined using the scratch assay. 22 Migration of Hep2 cells treated with ASCL1-siRNA1 or non-targeting siRNAs was examined on the six-well plates. A wound was created by manually scraping the monolayer of cells with a 10 µL pipette tip. The cells were washed with PBS and a filtered medium was added. An image of cells migrating into the wound was taken at the 12-h interval during a period of 48 h, and the ImageJ software was used to measure the wound area at the 12-h interval.
Cell invasion assays
The Transwell assay was used to measure the invasion capability of Hep2 cells which were transfected with control siRNA or ASCL1-siRNA1. After 16 h, transfected cells were trypsinized and resuspended, and 1.0 × 104 cells in 200 µL RPMI 1640 medium were placed into the upper chambers (8-mm pore size; Millipore). The lower chambers were filled with 600 µL complete medium with 10% FBS. After incubating for 48 h at a temperature of 37°C, non-invading cells were removed from the top of the chamber with a cotton swab, except that the Transwell membranes were pre-coated with 24 mg/mL Matrigel (Corning, NY, USA). The invaded cells on the lower surface of the inserts were fixed and stained with 0.1% crystal violet, and five random fields for each insert were counted at 200× magnifications. The Transwell assays were conducted two times during two separate experiments. Experiments were performed three times.
Co-immunoprecipitation
Lysate samples (800 µg of protein) were subjected to immunoprecipitation with 8 µg of anti-ASCL1 (Santa Cruz Biotechnology) antibody. Immune complex was formed overnight at a temperature of 4°C with Carousel. Following an overnight incubation, hatch samples were loaded with 80 µL of PRO-A bead slurry and hatched for 3 h at a temperature of 4°C. Samples were then centrifuged and washed, and 30 µL of Laemmli sample buffer was added to the PRO-A bead slurry and heated at a temperature of 100°C for 5 min. A volume of 10 µL of eluted sample was loaded in duplicate onto a 12.5% for western blot. Following electrophoresis, resolved proteins were transferred onto PVDF membranes, blocked, and then probed with anti-DKK1 (Santa Cruz Biotechnology) primary antibody overnight.
Statistical analysis
Statistical analysis was carried out using the SPSS statistics 19 software package through a paired test of comparison with the expression of ASCL1 protein. A rank-sum test was done to analyze the clinicopathological parameters of ASCL1 expression in laryngeal cancer. A survival curve was made using the Kaplan–Meier method, and the log-rank test was used for analysis. Multivariate analysis was conducted using Cox proportional hazards’ model; the risk ratio and its 95% confidence interval were recorded for each marker. Probability values <0.05 were considered statistically significant in all of the analyses.
Results
Expression of ASCL1 in sections from fresh and paraffin-embedded sections
The expression of ASCL1 was detected in eight paired fresh tissues (Figure 1(a)) using the western blot assay. The expression of ASCL1 in the tumors was dramatically higher than that in the adjacent non-tumorous tissues, while the amount of GAPDH, a housekeeping protein, was demonstrated to be rather constant among the samples. Figure 1(b) shows that the expression of ASCL1 is relative to GAPDH constant value (p < 0.05).

ASCL1 was found to be overexpressed in laryngeal carcinoma but is low or does not exist in paired adjacent non-tumorous tissues. (a) Western blot of eight representative paired samples of laryngeal carcinoma tissue (T) and adjacent non-tumorous tissues (N). In all samples tested, ASCL1 expression levels in laryngeal carcinoma were considerably higher than those in paired adjacent non-tumorous tissues. (b) The bar chart demonstrates the ratio of ASCL1 protein to GAPDH by densitometry. The data are mean ± SD of three independent experiments (p < 0.01, tumor tissues compared with adjacent non-tumorous tissues). Immunohistochemical analysis of ASCL1 expression. (c) and (d) Adjacent tissues and well differentiated laryngeal carcinoma tissues, respectively. (e) The high expression of ASCL1 in poorly differentiated histological grade of laryngeal cancer tissues (SP 400×). Details of the experiments are given in the “Materials and methods” section.
To further evaluate the ASCL1 expression and its clinical significance in 98 laryngeal cancer biopsy samples, immunohistochemical (IHC) stain was performed to determine ASCL1 expression in different histological grades as shown in Figure 1. It was clear that the low histological grade examples had a higher dyeing depth of ASCL1 than the well and moderate histological grade examples (Figure 1(c)–(e)). ASCL1 expression in laryngeal cancer was scored as positive when expressing in the cytoplasm, whose positive percentage ranges from 7% to 95%, with an average positive percentage of 53%. The common ground between western blot and paraffin-embedded sections is that both of them had a higher expression level of ASCL1 in laryngeal cancer.
Correlation of ASCL1 expression with clinicopathological variables in laryngeal cancer
The association of ASCL1 expression with clinicopathological variables was evaluated by rank-sum test. The clinicopathological data of the patients were summarized in Table 1. To statistically analyze the expression of ASCL1, carcinoma specimens were divided into high and low expressers according to the score of ASCL1-positive cells. Thus, ASCL1 expression in laryngeal cancer was significantly correlated with the level of histological differentiation (p = 0.000), T classification (p = 0.002), and lymph node metastasis (p = 0.000). There were no statistically significant differences between ASCL1 expression and other prognostic factors (Table 1).
Univariate and multivariate analyses of prognostic variables of patients with laryngeal cancer
Further univariate and multivariate analyses were conducted using a Cox proportional hazards model to examine the impact of ASCL1 expression and other clinical pathological parameters on patients suffering from laryngeal cancer. ASCL1 expression (p = 0.000), T classification (p = 0.000), lymph node metastasis (p = 0.001), histological differentiation (p = 0.002), and tumor site (p = 0.007) were essential prognostic factors for the univariate analysis (Table 2), while multivariate Cox regression analyses showed that ASCL1 was an independent predictor. Thus, ASCL1 expression may be a useful predicting factor for the overall survival of laryngeal cancer patients (p < 0.05, Table 2).
Univariate and multivariate analyses of overall survival in 98 laryngeal SCC specimens.

SCC: squamous cell carcinoma; CI: confidence interval.
Statistical analyses were performed by log-rank test.
Statistical analyses were performed by Cox regression model.
Relatively significant.
More relatively significant.
Most relatively significant.
High expression of ASCL1 predicted poor prognosis of laryngeal cancer patients
The Kaplan–Meier survival analysis was carried out in order to evaluate the prognostic value of ASCL1 expression for the overall survival rate of laryngeal cancer patients. It was suggested that the overexpression of ASCL1 is significantly connected with poor prognosis. In other words, the laryngeal cancer patients with low ASCL1 expression have obviously longer overall survival rates than those with high ASCL1 expression (Figure 2; p = 0.000).
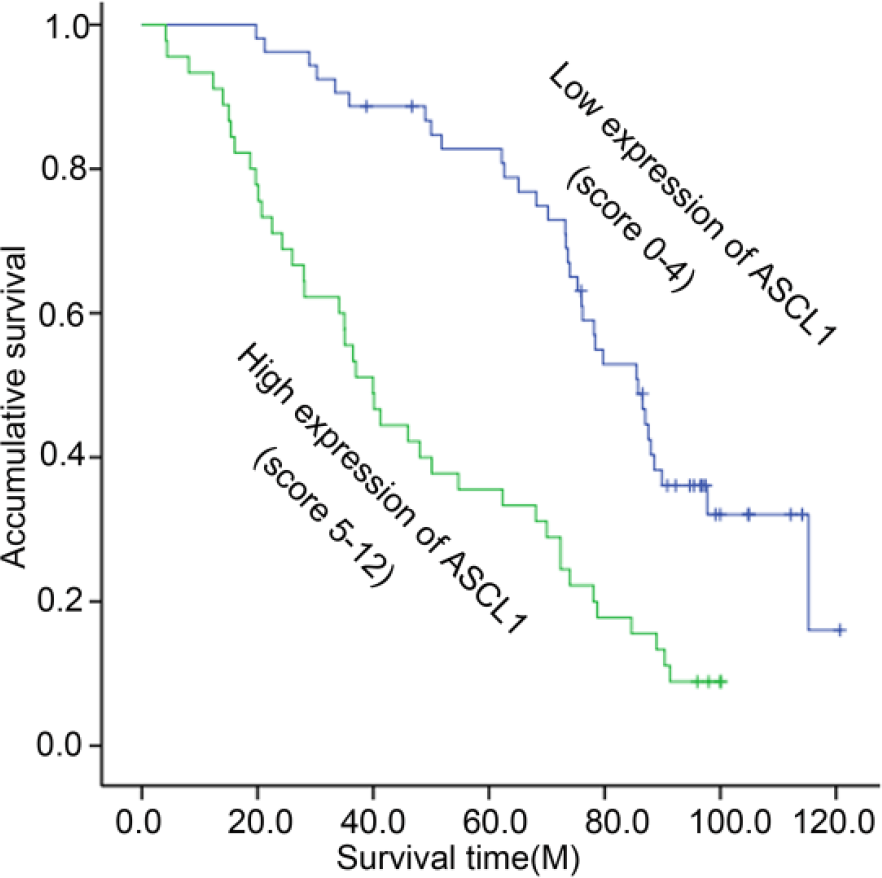
Kaplan–Meier survival curves for low ASCL1 expression versus high ASCL1 expression of 98 patients with laryngeal cancer. On the basis of average ASCL1 percentages, patients were divided into low ASCL1 expressers (score ≤ 4) and high ASCL1 expressers (score ≥ 5). Patients with high ASCL1 expression had significantly shorter overall survival (p < 0.01) than those with low ASCL1 expression.
Knockdown of ASCL1 expression suppressed cell proliferation in laryngeal cancer cell lines TU212
In order to study the imperceptible role of ASCL1 in laryngeal cancer cell proliferation, TU212 cells were transiently transfected with ASCL1-siRNA or control siRNA via lipofection 72 h later, and ASCL1 protein levels were substantially decreased in cells transfected with ASCL1-siRNA compared to the control siRNA-transfected cells. Among the four siRNAs tested (Figure 3(a)), ASCL1-siRNA1 had the highest knockdown efficiency and therefore used in all the experiments. To assess the imperceptible role of ASCL1 knockdown on cell viability, ASCL1-siRNA1 and control siRNA were transfected into TU212 cells by means of CCK-8 assay, showing that TU212 cells which were treated with ASCL1-siRNA1 had a significant decrease in cell viability compared with the control siRNA (Figure 3(b)). And western blot was adopted to detect the expressions of cyclin A and CDK2, and cell proliferation markers were downregulated, while the expression of p27Kip1 was upregulated (Figure 3(c)). The flow cytometry was used to examine the imperceptible role of ASCL1 in the cell-cycle distribution changed after transient transfection of the ASCL1-siRNA1 expression vector in laryngeal cancer cells. FACS analysis of cell-cycle distribution discovered that ASCL1 knockdown led to the arrest of G0/G1–S phase a concomitant reduction in the number of cells in S phase compared with control siRNA (Figure 3(d)), showing that ASCL1 might have participated in G0/G1–S transition of cell cycle. These results further prove that ASCL1 plays an important role in the proliferation of laryngeal cancer cells.

The knockdown of ASCL1 inhibited the laryngeal cancer cells from proliferation and S-phase entry. (a) ASCL protein expression was detected using the method of western blot after siRNA transfection of TU212 cells for 72 h. Western blot showed that ASCL1-siRNA1 had the highest knockout efficiency among the four siRNAs tested. The bar chart below demonstrates the ratio of ASCL1 protein to GAPDH by densitometry. The data are mean ± SD (p < 0.01 compared with the control). (b) Cell viability was measured using the CCK-8 assay. Cell Counting Kit-8 reagents were added to the medium and incubated for an additional 2 h. Absorbance was measured to examine the siRNA-treated TU212 cells. Data show mean ± SD of triplicates from one experiment representative of three experiments done versus the control (p < 0.05). (c) Western blot analysis of cyclins A, CDK2, and p27Kip1 in ASCL1 knocked down TU212 cells. (d) As described above, the cells were stained with PI for DNA content analysis by FACS. Details of the experiments are given in the “Materials and methods” section.
Knockdown of ASCL1 expression induces cell apoptosis in laryngeal cancer cell lines TU212
Finally, we tested the effects of ASCL1 on the apoptosis of laryngeal cancer cell. Since knockdown of ASCL1 expression could prevent the cancer cell from growing, we detected the survival of laryngeal cells with ASCL1-siRNA1-transfected or control RNAi-transfected cells after their detaching and replanting in dishes coated with polyhydroxyethylmethacrylate. And we found that silencing the ASCL1 expression could significantly increase the cell apoptosis compared with control siRNA 48 h after being transfected in the cell apoptosis assay (Figure 4(a)). And we also learnt that ASCL1 could activate the antiapoptotic regulator BCL2 (Figure 4(b)). All in all, these findings suggest that the inhibition of ASCL1 in TU212 cells could not only limit cell growth but also bring about its apoptosis.

The knockdown of ASCL1 inhibited the TU212 cells. (a) Downregulation of ASCL1 in TU212 cells could cause cell apoptosis. (b) Cells were lysed and analyzed using western blot of BCL2, and GAPDH was used as loading control (*p < 0.05). Details of the experiments are given in the “Materials and methods” section.
ASCL1 promotes the migration and invasion of Hep2 cells
To find out the effects of ASCL1 on the migration of laryngeal cancer cells, cell scratch assays were performed using two independent assays. The results of the cell scratch assays show that the area changed as a result of cell scratch in SiRNA1-ASCL1 was reduced compared with the control (Figure 5(a) and (b)). And we used the method of Transwell assay to examine the potential effects of ASCL1 inhibition on laryngeal cancer cell invasion. As shown in Figure 5(c), the invasion of Hep2 cells facilitated by control ASCL1-siRNA was stronger than by ASCL1-siRNA1.

The knockdown of ASCL1 inhibited the Hep2 cells. (a) Cell migration was measured using the cell scratch assay and reagents were added to the medium and incubated for an additional 0, 24, and 48 h. (b) A graph showing the healing process at different time points. (c) Cell invasion was measured using the Transwell (SP 400×). Details of the experiments are given in the “Materials and methods” section.
ASCL1 regulates the expression of epithelial–mesenchymal transformation–associated proteins in laryngeal cancer cell lines Hep2
To better understand the mechanism of ASCL1 which regulates cell migration and invasion, the protein levels of metastasis-associated and epithelial–mesenchymal transformation (EMT)-associated proteins in laryngeal cancer cell lines were tested by knocking down the ASCL1 expression. It was found that suppressing ASCL1 expression could increase the expression of epithelial marker proteins including Zo-1 and E-cadherin and decrease the expression of mesenchymal marker proteins including vimentin and N-cadherin, an indication that the knockdown of ASCL1 could suppress the EMT process (Figure 6(a)). In addition, expressions of MMP-3 and MMP-9 underwent no change after suppressing ASCL1 in laryngeal cancer cell lines (Figure 6(a)). In order to further explore the mechanisms by which ASCL1 promoted the invasion of Hep2 cells, as shown in Figure 6(b), ASCL1 interacted with DKK1 in Hep2 cell lines via co-immunoprecipitation. It was also found that inhibition of ASCL1 expression could lead to downregulation of DKK1 (Figure 6(b)).

ASCL1 controls the expression of EMT-associated genes in laryngeal cancer cell lines. (a) ASCL1 expression could decrease the expression of epithelial marker genes including ZO-1 and E-cadherin and increase the expression of mesenchymal marker genes including vimentin and N-cadherin. (b) The interaction between ASCL1 and DKK1 in laryngeal cancer cell lines Hep2. Details of the experiments are given in the “Materials and methods” section.
Discussion
It was found that ASCL1 could facilitate the development of various human carcinomas, such as oral squamous cell carcinoma, 17 cervical carcinoma, 10 gastrointestinal carcinoid, 7 small-cell lung carcinomas, 23 high-grade neuroendocrine lung cancers, 11 pancreatic endocrine tumors, 9 and medullary thyroid cancer. 14 Osada et al. 11 suggest that the interaction between ASCL1 and DKK1 could promote the migration and proliferation of pancreatic and lung cancers. 9 The expression of ASCL1 in laryngeal cancer is, however, still unknown. And we were the first to find that the expression of ASCL1 in laryngeal cancer tissues was higher than its paired adjacent normal laryngeal mucosa, a manifestation that ASCL1 does have some effects on the proliferation and migration of laryngeal cancer cell lines.
In this study, we first characterized the expression of ASCL1 by comparing the laryngeal carcinoma with their corresponding noncancerous tissues and found that the level of expression of ASCL1 in laryngeal carcinoma was increased, which was considerably connected with histological differentiation, T classification, lymph node metastasis, and poor survival. The relationship between the overexpression of ASCL1 and T classification suggested that the overexpression of ASCL1 may bring about rapid increase in laryngeal tumor volume. This result is consistent with that of the research of lung adenocarcinoma. 5 Additionally, ASCL1 was overexpressed in laryngeal cancer with high migration level, but profoundly expressed weakly in laryngeal cancer with low migration level, suggesting that ASCL1 may be of a tumor-accelerating property. 24 Besides, multivariate analysis showed that ASCL1 protein was an important independent prognostic factor for overall survival, and ASCL1 could be used as a new predictor of the prognosis of cases with laryngeal cancer.
The previous study suggested that ASCL1 could delay cell-cycle exit and differentiation16,24–26 and induce the proliferation of medullary thyroid cancer cells 27 and gastrointestinal carcinoid cells. 7 However, the biological function of ASCL1 in laryngeal cancer is not known yet. In order to explore the function of ASCL1 in laryngeal cancer, loss-of-function studies were carried out and found that decreased ASCL1 expression could reduce the growth, migration, and invasion of laryngeal cancer cell lines and induce their apoptosis. In addition, immunoblotting also suggested that cyclins A and CDK2 expressions were downregulated, while the expression of p27Kip1 in ASCL1 interference cells, a tumor suppressor gene28,29 which was found to be vital in cell-cycle progression, was on the contrary. A further research has found that ASCL1 could activate the antiapoptotic regulator BCL2, which is similar to the role of ASCL1 during lung carcinogenesis. 15 In a word, the suppression of ASCL1 may inhibit cell proliferation and lead to apoptosis.
Besides the finding that ASCL1 could not only promote proliferation and decrease apoptosis but also facilitate the migration of oral squamous cell 17 and pancreatic endocrine tumor cell line, 9 a further study has confirmed the potential role of ASCL1 in the migration and invasion of tumor cells in laryngeal cancer cell lines. Cell movement in Hep2 cells was detected through the use of small interfering technology, with RNAi-ASCL1 as control. In the wound-healing and Transwell assays, we found that suppressing ASCL1 expression by siRNA could inhibit cell migration and invasion.
While exploring the molecular mechanism of ASCL1, we found that suppressing ASCL1 expression could increase the expression of epithelial marker genes including ZO-1 and E-cadherin and decrease the expression of mesenchymal marker genes including vimentin and N-cadherin, which exerted no effect on MMP-3 and MMP-9; however, this is an indication that ASCL1 might regulate the migration of laryngeal cancer cells through EMT process. We also found that ASCL1 could form complex with DKK1 and reduce the expressions of DKK1 and E-cadherin.9,11 DKK1, as an inhibitory factor of Wnt/beta-catenin signal pathways, could destroy the signal pathway of lipoprotein receptor–related factor of low density (LRP5/6) which was combined with the Wnt/FRZ complexes and blocks the signal transduction pathways and were involved in human cancer development, 30 which was similar to ASCL2 promoting colon EMT. 31 So, we speculated that ASCL1 could promote the migration and invasion of laryngeal cancer of EMT by activating Wnt/β-catenin signal pathways.
Conclusion
This study is the first to explore ASCL1 effects on laryngeal carcinogenesis and especially clarifies for the first time the relationship between ASCL1 and DKK1 in laryngeal carcinoma. It also found that ASCL1 can facilitate EMT, cell proliferation, migration, and invasion as well as cell-cycle arrest and induce apoptosis of laryngeal carcinoma by binding to Wnt inhibitor DKK1. The findings prove that ASCL1 may serve as a novel prognostic biomarker and a new molecular therapy for laryngeal cancer. More medicine-related studies will be necessary to better prove the function of ASCL1 in treating laryngeal cancer.
Footnotes
Acknowledgements
H.M. and X.D. are co-first authors.
Compliance with ethical standards
All procedures performed in this study involving human participants were in accordance with the ethical standards of the institutional and national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards. This article does not contain any studies with animals performed by any of the authors.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by Six Talent Peaks Project of Jiangsu Province (WSW-054).