
Abstract
Cancer is the first cause of death in developed countries and the second in developing countries. Concerning the most frequent worldwide-diagnosed cancer, primary liver cancer represents approximately 4% of all new cancer cases diagnosed globally. However, among primary liver cancer, hepatocellular carcinoma is by far the most common histological subtype. Notwithstanding the health promotion and disease prevention campaigns, more than half a million new hepatocellular carcinoma cases are reported yearly, being estimated to growth continuously until 2020. Taking this scenario under consideration and the fact that some aspects concerning hepatocellular carcinoma evolution and metastasize process are still unknown, animal models assume a crucial role to understand this disease. The animal models have also provided the opportunity to screen new therapeutic strategies. The present review was supported on research and review papers aiming the complexity and often neglected chemically induced animal models in hepatocarcinogenesis research. Despite the ongoing debate, chemically induced animal models, namely, mice and rat, can provide unique valuable information on the biotransformation mechanisms against xenobiotics and apprehend the deleterious effects on DNA and cell proteins leading to carcinogenic development. In addition, taking under consideration that no model achieves all hepatocellular carcinoma research purposes, criteria to define the “ideal” animal model, depending on the researchers’ approach, are also discussed in this review.
Introduction
Cancer is seen as a threatening disease by public opinion and responsible for one in eight deaths worldwide, being the second cause of death in developed countries and the third leading cause in developing countries. Globally, proportions of liver cancer are more than twice as high in males than in females. In men, it is the fifth most frequently diagnosed cancer worldwide versus the seventh most commonly diagnosed cancer in women. Considering cause of cancer death, liver cancer is the second most frequent in men versus the sixth leading cause of cancer death in women.1,2 Among primary liver cancers, hepatocellular carcinoma (HCC) represents 85% of liver cancers worldwide. 3 Further kinds of liver cancer comprise cholangiocarcinoma, angiosarcoma, or haemangiosarcoma and hepatoblastoma.3–6
HCC has recently received particular attention due to the increasing mortality and morbidity, involving more than half a million new cases yearly, expecting to reach its plateau nearby 2015–2020. 7 Remarkable differences can be found in HCC incidence and mortality in gender, race/ethnicity, age, geographical regions (due to socioeconomic development), and our understanding on behavior change and health promotion (e.g. campaigns promoting the reduction of drinking and smoking habits, health programs on unsafe sexual practices, and needle sharing among injecting drug users1,3,8–10).
All over the years, a myriad of chemicals capable of inducing liver cancer have been tested in animal models, namely, dogs, 11 Mongolian gerbils, 12 Guinea pigs, 13 Syrian golden hamsters, 14 rabbits, 15 pigs, 16 monkeys, 17 rats, 18 and mice.19,20 The methods applied using those models are useful to study premalignant and malignant liver lesions allowing to extrapolate the biopathology mechanisms underlying hepatic human tumors and also to evaluate new therapeutic strategies.21–25
Due to the daily life exposure to countless chemicals, it is important to ensure its safety concerning toxicity and carcinogenicity. Some of these chemicals are suspected to induce cancer in several mammals’ species including humans. The present review will focus on the often neglected importance of chemically induced HCC models and the main mechanisms of action.
Risk factors
HCC may be seen as the ending process of chronic liver diseases, starting from fibrosis, to cirrhosis, and cancer as a final stage; this clinical history has been reported in 80% of people with HCC. 26 In human species, cirrhosis is the major clinical risk factor for hepatic cancer. Chronic hepatitis B virus (HBV) and hepatitis C virus (HCV) infection are well-established HCC etiological factors worldwide. Interestingly, HCV, HBV, and alcohol also cause DNA damage via activation of inflammation and reactive oxygen species (ROS). Despite the worldwide campaigns to prevent HBV and HCV infections, young adults often have difficulty to understand the cumulative implications of recurrent exposures risks overtime, misperceiving the true risk impact on their health. 27
Toxic exposure (alcohol abuse, aflatoxin-B1 (AFB1) intake from contaminated food), obesity, metabolic diseases (starting from non-alcoholic fatty liver disease (NAFLD), non-alcoholic steatohepatitis (NASH)), and autoimmune reactions also contribute to HCC development.24,25,28–30 Recently, it has been suggested that many cryptogenic cirrhosis and HCC cases found in Western countries (30%–40% proportion of patients) characterize in fact severe forms of NAFLD, specifically NASH, which might have been promoted partially by diabetes and obesity. 31
Animal models and HCC research
Animal testing has played a central role in biomedical research throughout history, despite the controversy involving animal research and their use to predict human outcomes is far from settled.32,33 The need of experimental animal models of liver tumors that are analogous to humans has been a challenge to researchers attempting several approaches to elucidate the mechanism of human hepatocarcinogenesis. The “ideal” animal model (Figure 1) should reproduce human HCC natural history, biopathology, and biochemistry; 34 allow the evaluation of potential novel therapeutic drugs in pre-clinical trials against this disease; 25 and contribute to the development of molecular target therapy. One should remind that no model, however, is “ideal” for all HCC research purposes35–37 as well for the study of other diseases. Spontaneous models are scarcely rare. However, they are the ones who give an accurate perspective concerning cell growth pattern, antigenicity, cell metabolism, and cell differentiation. 38 Transplantation models have provided new insights concerning hepatocarcinogenesis stages of invasion-metastasis cascade and validation of new therapeutic approaches.39,40

Advance HCC has no effective treatment or cure, despite the discovery of promising novel therapeutic drugs. The failure of treatments to improve the patients’ survival should be related to tumor heterogeneity and to hepatic dysfunction. Part of the solution to this problem may rely on molecular target therapies and the assessment of their efficacy in HCC. 42 A wide range of HCC animal models are currently available to study HCC development, metastasis, and treatment. For several reasons (size, short life span, breeding capabilities, genetic engineering options, and similarity to human hepatic lesions at histological and molecular levels), the mouse (Mus musculus) has long been regarded as one of the animal models most suitable for this purpose.24,43–45
Experimentally induced liver tumors
A relatively large number of rodent models to study hepatocarcinogenesis are presently available and can be organized as follows: (a) chemically induced models, (b) implantation models (syngeneic and xenograft models), (c) viral models, and (d) genetically engineered mouse (GEM) models (Figure 2).34,38,39,46,47
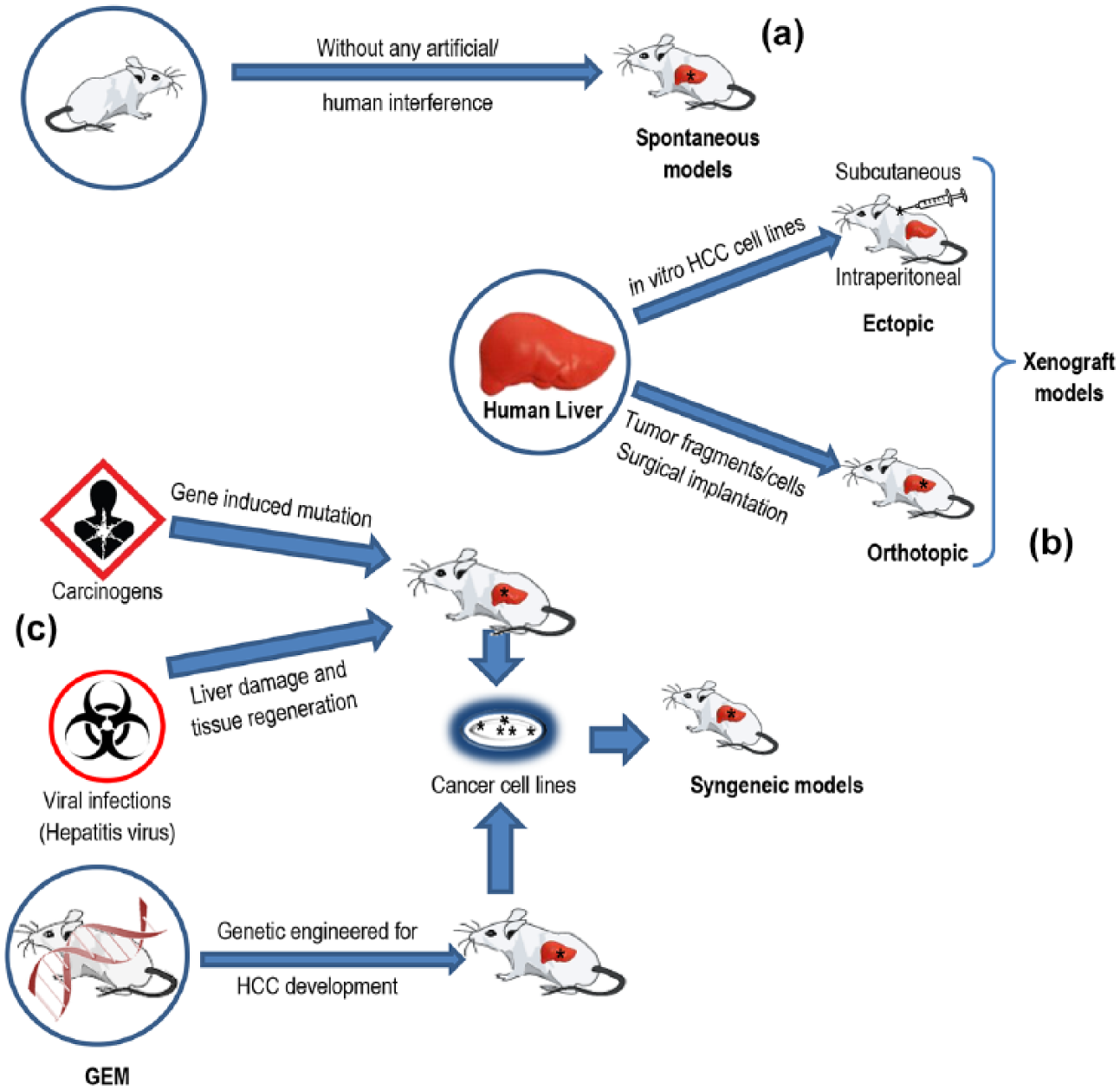
Models of hepatocellular carcinoma: (a) spontaneous, (b) xenograft, and (c) syngeneic models.
Syngeneic models comprise tumors that originated in the same strain as a result of spontaneous causes, hepatocarcinogen, viral infections, and GEMs (Figure 2).34,38,39
Chemically induced models
The human society trusts on the use of chemical compounds to fulfill a large number of needs from food production, process of conservation/storage, agriculture purposes, and novel applications on industrial process. 48 Many of these compounds are also able to induce carcinogenesis after acute, short-, or long-term exposure, depending on the chemical structure, chemical concentration, animal species, and the animals’ susceptibility. Since all living organisms are inevitably exposed to chemical compounds, the liver is the primary target due to its essential role in xenobiotic detoxification. 49 Biotransformation mechanisms against xenobiotics are crucial to prevent its accretion and deleterious consequences. 50 Nonetheless this preventive role, some of the metabolites are capable of establishing covalent bounds called adducts with DNA and cell proteins 51 leading to carcinogenic development.
The International Agency for Research on Cancer (IARC) 52 and the U.S. National Toxicology Program (NTP) 53 publish science-based reports on public health dedicated to identify substances or exposure circumstances to be known or to be reasonably anticipated as human carcinogens (Table 1). The chemical substances mentioned in this review are identified with CAS number and standard worldwide categories according to IARC group and NTP category.
Synopsis of the IARC and NTP categories taking under consideration the overall evaluation of the analyzed substances and its risk to human health. This categorization reflects the global evaluation of relevant data from human and animal models studies.

IARC: International Agency for Research on Cancer; NTP: U.S. National Toxicology Program.
Diethylnitrosamine (CAS No. 55-18-5; Group 2A; 54 RAHC53,55)
N-nitroso compounds (NOCs) are well known for their mutagenic, teratogenic, and carcinogenic activities.56,57 The first hint came in the early 1960s from Norway, when sheep died of liver toxicity after feeding nitrite-preserved herring meal.57,58 The effect of nitrate, nitrite, and NOCs on human health has been a recurrent issue with respect to cancer59–62 due to unawareness of the general population exposure to NOCs, namely, diethylnitrosamine (DEN), present in foods, beverages, tobacco smoke,57,63–65 drinking water, and industrial pollution.66,67 Endogenous synthesis of NOCs are also mentioned for being responsible for 45%–75% of total exposure.68,69 In 1956, Magee and Barnes reported evidence of liver carcinogenesis by N-nitrosamines (dimethylnitrosamine (DMN)) in male and female albino rats fed with powder diet containing DMN solution in arachis oil (10 mL/kg diet powder). Nineteen out of 20 rats developed primary liver tumors, and metastatic spread was noticed in seven rats (pulmonary and intra-abdominal metastasis); these hepatic tumors took between week 26 and 40 to develop. 18 Since then, this class of chemical compounds has received special attention due to their carcinogenesis induction in almost all experimental animal models tested, showing in some cases, target organ specificity, for example, DEN causes tumors mainly in the liver, respiratory tract, kidney, upper digestive tract, and the hematopoietic system. 70 In mice exposed to DEN, dosage, age, sex, and strain influenced tumorigenesis 46 as discussed further. Based on experimental animals studies, DEN is reasonably anticipated to be classified as a human carcinogen,55,56 despite the lack of epidemiological data. 71 Some hepatocarcinogens have the ability to presumably origin cancer throughout DNA adducts formation, oxidative alteration and strand breakage. 72 If not repaired, or the damaged cells destroyed, the genetic changes may enhance cell replication, increasing the probability of mutation; these compounds are known as genotoxic (or direct-acting) carcinogens and act as initiators.19,24,73 Examples of liver genotoxic agents are not only the above-mentioned DEN and DMN but also others such as 2-acetylaminofluorene (2-AAF), N-nitrosomorpholine (NMOR), and AFB1,25,34,74 If a carcinogen displays proprieties of both initiators and promoters, it is considered to be a complete carcinogen; if it only instigates irreversible DNA impairment, it is considered to be an incomplete carcinogen. 48
Conversely, the non-genotoxic carcinogens have no direct interaction with DNA and display intricate and varying mechanisms that are not completely understood. These mechanisms are believed to act through pathways inducing cellular structures disruption or damage, increasing the risk of genetic error, and stimulating cell evolution into malignant neoplasm by affecting the proliferation, differentiation, and apoptosis mechanisms.24,73,75,76 These compounds are called tumor promoters and include phenobarbital (PB), thioacetamide (TAA), di(2-ethylhexyl)phthalate (DEHP), methapyrilene (MP), carbon tetrachloride (CCl4), and tamoxifen.72,77
It is widely described that the chemically induced models for carcinogenesis is an intricate and stepwise process, involving at least three distinct stages (from an operational point of view)—initiation, promoting, and progression.3,38,48 Although being the most common process described, other authors prefer to conceptually divide the multistage chemical carcinogenesis into four steps: tumor initiation, promotion, malignant conversion, and progression, distinguishing the malignant conversion from the tumor progression stage. 78 Notwithstanding the suggested conceptual model, several chemically carcinogenic protocols have been proposed to study liver carcinogenesis, taking under consideration the species and the aim of the research, that is, carcinogenesis target organ, novel therapeutic approach, and characterization of preneoplastic and neoplastic evolution. Concerning the mouse model, besides the administration route and dosage, other aspects are also taken under consideration, that is, age, gender, and strain; hepatocyte proliferation rates are higher in juvenile mice and gender-related differences—higher HCC prevalence in males—appear to be a result of the promoting effects of androgens and the inhibitory effects of estrogens in females.19,47 Table 1 outlines several animal models commonly used in hepatocarcinogenesis investigations as reviewed, including study design, strain, gender, and main features.
According to summarized data in Table 2, DEN is one of the most widely hepatotoxic chemicals used to induce HCC animal model and was first listed in the Second Annual Report on Carcinogenesis in 1981.55,71 Its carcinogenesis process goes through liver injury, liver fibrosis, liver cirrhosis, and ultimately HCC development as proposed in our schematic sequence in Figure 3.94,95 Despite the fact that some of the chemical hepatocarcinogenesis events remain uncertain, the multistage process—Initiation ⇒ Promotion ⇒ Progression—is an accepted theory.72,96 Its complexity can only be studied in vivo involving all aspects of exposure and target organ biology. 96 Regarding DEN, if it is injected into younger mice (less than 2 weeks), it acts as a complete carcinogen, due to higher hepatocyte proliferation rates in the juvenile.47,97 If the mice are older than 2 weeks, subsequent to DEN administration, a tumor promoter is required, which enhances hepatocarcinogenesis. This can be achieved by adding the following promoters: PB, NMOR, and CCl4, as described in Table 2. Most recently, Park et al. 98 demonstrated that either high fat diet (HFD) or genetic obesity can act as a liver tumor promoter in mice. However, despite the mice age difference (i.e. age: 6 weeks), Da Costa et al, 99 using DEN in male ICR mice, reported several preneoplastic features and proliferative lesions, including HCC, 40 weeks after the last exposure to multiple intraperitoneal injections (i.e. for eight consecutive weeks).
Synopsis of chemically induced HCC—models.

HCC: hepatocellular carcinoma; s.d.: single dose; m.d.: multiple doses; i.p.: intraperitoneal injection; s.c.: subcutaneous injection; d.w.: drinking water; d.: diet; i.g.: gavage; DEN: N-diethylnitrosamine; 2-AAF: 2-acetylaminofluorene; DMN: N-nitrosodimethylamine; NMOR: N-nitrosomorpholine; AFB1: aflatoxin B1; DMSO: dimethylsulfoxide; TAA: thioacetamide; CCl4: carbon tetrachloride; PB: phenobarbital;
In two stage model - a: initiator; b: promoter.

The sequences of the main events in DEN-induced liver tumors in mice.
In addition to the above mentioned necrogenic dose of a hepatocarcinogen or a non-necrogenic conjugated with partial hepatectomy (PH) as the promoting stimuli to induce clonal expansion of initiated cells as described in the Solt–Farber protocol 100 have been also used in hepatocarcinogenesis studies.82,101 Cancer development, both in vivo and in vitro models, is associated with genotoxic and cell proliferative stages; PH is associated with genotoxic action in order to induce cell proliferation in the liver, both in adults102,103 and in neonatal animals. 104
According to the chemical model option (one-stage model—using only initiator proprieties, or two-stage model—using the association of initiator and promoter), the genomic expression should result in activation of Ha-ras mutations, induced by single DEN injection, while DEN/PB (two-stage model protocol) has a propensity to activate β-catenin proto-oncogene instead of causing genomic instability.19,30,47,105–107
DEN biotransformation requires metabolic activation catalyzed by the liver cytochrome P450 (CYP) system (subfamily CYP2A6 and mainly by CYP2E1), having its highest activity in the centrilobular hepatocytes—zone III (pericentral) of the liver lobule.19,20,46,64,108,109 The CYPs are membrane-bound heme-containing enzymes that catalyze the oxidation of several endogenous and exogenous compounds. 20 Severe centrilobular hydropic degeneration was noticed in mice exposed to DEN, after 8 weeks (post-exposure).44,84,110 Experimental models have clearly demonstrated that the modulation of P450 expression can modify the susceptibility of animals to cancer development induced by various chemical carcinogens. 111
The hydroxylation of DEN to α-hydroxylnitrosamine is molecular oxygen and nicotinamide adenine dinucleotide (NADPH)-reductase dependent. 71 DNA-adducts formation continues through the cleavage of acetaldehyde resulting the production of an electrophilic ethyldiazonium ion, triggering DNA damage by reacting with nucleophiles such as DNA bases, causing its methylation.19,46,71,112 The O 6 -ethyl-guanine is one of the main adducts associated with DEN carcinogenicity. 46 If the enzymatic repair mechanisms fail to repair DNA adducts, the initiation stage of carcinogenesis may occur. 48 Kaina et al. 113 emphasize on factors responsible for DEN carcinogenicity and the effective capability to remove adducts by using the enzyme O 6 -alkylguanine-DNA alkyltransferase by means of base excision and the repair of interstrand crosslinking and strand breaks. In addition to alkylation of DNA, ROS and free radicals resulting from P450-dependent enzymatic system, namely, CYP2E1, might induce oxidative stress by the formation of hydrogen peroxide and superoxide anions.19,112 ROS are known for interacting and inducing damages at different levels, for example, cellular protein, lipids, and DNA, causing cellular dysfunction. Therefore, oxidative stress is broadly recognized as a key player in hepatocarcinogenesis, both at initiation and progression stages.114–116
The activation of the β-catenin pathway in mice model is useful to support the importance of this model to decipher human HCC, since almost 30% of this cancers display mutations responsible for activating the β-catenin pathway.47,117–119 Nevertheless, one should be aware that despite the resemblance in metabolic pathways and their importance for both human and other animal species used in cancer research, qualitative and quantitative differences should be taken under consideration to avoid misinterpretation of the carcinogenic effect of tested chemical compounds when deducing to human risk assessment. 48
NMOR (CAS No. 59-89-2; Group 2B; 54 RAHC53,55)
NMOR belongs to the N-nitrosamine group likewise DEN and requires liver metabolic activation, and it was first listed in the Second Annual Report on Carcinogenesis in 1981. 55 According to Wesierska-Gaadek et al., 120 NMOR causes severe hepatotoxicity, and its carcinogenic effect has been corroborated in numerous animal models in diverse tissues sites using distinctive routes of exposure.55,121 Oral administration of NMOR (drinking water) in male mice caused benign hepatocellular tumors, several lung adenomas, and a restricted number of squamous cell carcinomas of the lung. 122 Identical results were achieved in rats (BD, Sprague-Dawley and Wistar), in addition to malignant liver and bile duct tumors (HCC, cholangiofibroma, or cholangiocarcinoma). 122 Intravenous injection of NMOR in rats was responsible for HCC. Immersion assays were also performed using animal models such as zebra fish (Danio rerio) and guppies (Lebistes reticulatus); both species after 31 weeks of exposure to NMOR developed HCC and hepatoadenomas. 122 The routes of potential human exposure to NMOR are dermal contact, ingestion, and inhalation. 55
Initiated liver cells arise and divide resulting in small preneoplastic cell clones. Due to its alkylating features, NMOR might modify the biological macromolecules and a defense strategy against potentially harmful cells may increase resulting in apoptosis, necrosis, and, subsequently, regenerative DNA synthesis, during tumor initiation.120,121
2-AAF (CAS No. 53-96-3; IARC not available; RAHC53,55)
2-AAF is an aromatic amine used as a research tool, mainly as a positive control in studies of the carcinogenicity and mutagenicity of other chemicals. 55 Since 2-AAF was first listed in the Second Annual Report on Carcinogenesis in 1981, further studies in experimental animal models were conducted. This compound is converted by liver microsomal monooxygenases to N-hydroxy-AAF123–125 followed by further activation steps leading to the formation of reactive forms causing toxicity, cell death, and carcinogenesis. 125 Oral exposure and subcutaneous and intraperitoneal administration caused liver cancer in mice, rats, dogs, and fishes. 55 2-AAF has also been used as a promoter agent associated with DEN in male Sprague-Dawley rats 126 and with PH approach in male Wistar rats and C57BL/6 mice. 127 Dietary administration of 2-AAF induced liver cancer (HCC) in both genders of mice and rats. 55 A single subcutaneous injection in newborn male mice induced hepatocellular tumors. Concerning the hamster model, intraperitoneal injection in newborn until weaning, associated with dietary administration, caused not only liver cancer but also urinary bladder and benign stomach tumors in both genders. 55 The routes of potential human exposure to 2-AAF are inhalation, ingestion, and dermal contact. 55
Aflatoxins (CAS No. 1402-68-2; Group 1; 54 KHC53,55)
A major risk factor concerning HCC is the aflatoxins intake (mostly AFB1). Human diet consisting of Aspergillus fumigatus, Aspergillus flavus, Aspergillus parasiticus, Aspergillus australis, and Aspergillus nomius contaminated food has been associated with synergistic effect between aflatoxin and HBV infections in Africa and Asia.1,9,128 However, despite the strong and significant positive correlation between estimated aflatoxin intake or its levels in food samples and the incidence of liver cancer, AFB1 contribution to HCC cases in those regions is still unknown.1,9,55 AFB1 was first listed in the First Annual Report on Carcinogenesis in 1980.55,129 The aflatoxins are a group of difuranocoumarin compounds, closely related mycotoxins that are widely distributed in nature in different agricultural communities, causing higher contamination on crops growing in hot and humid tropical climates.55,130,131 They have a very wide range of biological activities, causing great economic losses and health risks to both humans and farm animals. These mycotoxins are responsible for genetic damage, including formation of DNA and albumin adducts, gene mutation (e.g. mutation in p53 tumor-supressor gene has been found in 30%–60% of HCC in aflotoxin-endemic areas), micronucleus formation, sister chromatid exchange, and mitotic recombination.8,55,131,132
In susceptible animal species including humans, once ingested, AFB1 is metabolized by cytochrome P450 enzymes as an active intermediate, AFB1-exo-8,9-epoxide, a very reactive form that binds to DNA (forming adducts with the N7 position of guanine residues) and also to albumin in the blood serum.8,130 In several assessed biological systems, the most frequently observed mutation induced by chemically reactive forms of AFB1 is the GC→TA transversion (interchanging of purine ↔ pyrimidine).129,133 This mutation was confirmed in Escherichia coli, rainbow trout, F344 rats, transgenic C57BL/ 6N (BigBlue) mice, human hepatocytes in culture, and in human liver tumors. 129 Susceptibility differences among mice strains are a consequence of several AFB detoxification loci. According to McGlynn et al., 132 7-day-old mice (from strains inbred C57BL/6J, DBA/2J) exposed to a single administration of 6 mg/kg body weight of AFB1 developed HCC after 52 weeks. DBA/2J animals were threefold more sensitive to AFB1-induced HCC and significantly more sensitive to AFB1 acute toxicity than were C57BL/6J animals.19,132
The AFB1-exo-8,9-epoxide metabolite can be detoxified through conjugation with glutathione, mediated by the enzyme glutathione S-transferase (GST). This enzyme is particularly higher in animal species resistant to aflatoxin carcinogenicity, such as mice, than in susceptible animal species such as rats. Compared to rats and mice, humans have lower GST activity, which suggests a minor capacity of detoxifying AFB1-exo-8,9-epoxide. 131
TAA (CAS No. 62-55-5; Group 2B; 54 RAHC53,55)
TAA is an organosulfur compound first listed in the Third Annual Report on Carcinogenesis in 1983.55,129 It has been used as an organic solvent in the leather, textile, and paper industries, 134 as a fungicide, accelerator in the vulcanization of rubber, and as a stabilizer of motor oil. 135 TAA is used to induce acute and chronic liver injury in mice and rats either in diet containing 0.03%–0.1%, 136 drinking water (0.02%–0.05%), or by intraperitoneal administration (0.1%).19,137 As a potent centrilobular hepatotoxicant, TAA undergoes a two-step bioactivation by a mixed-function oxidase system, namely, by CYP and/or flavin-containing monooxygenase (FMO) systems to sulfine (sulfoxide) and sulfene (sulfone) metabolites.134,135,137,138 In vivo administration in rodents induced highly selective liver damages including cirrhosis, fibrosis, as well as hepatic necrosis/apoptosis.135,139 Liver damage results from the mediation of microsomal CYP2E1 to TA sulfoxide (TASO), and further to TA-S,S-dioxide (TASO2), which ultimately modifies amine-lipids and proteins to initiate cellular necrosis.138–140
The primary routes of potential human exposure to TAA are inhalation and dermal contact. 55
CCl4 (CAS No. 56-23-5; Group 2B; 54 RAHC53,55)
Since the 1950s, that CCl4 has been used for free radical-induced liver injury model.141,142 The experimental evidences of carcinogenicity in several animal models supported its classification as reasonably anticipated to be a human carcinogen; it was first listed in the Second Annual Report on Carcinogenesis in 1981. 55 Benign or malignant liver tumors were found in both genders in mice, rats, and hamsters orally exposed to CCl4; identical results were also described in rainbow trout. In male rats, subcutaneous administration was responsible for hepatocarcinogenesis; early to well establish liver carcinomas were additionally reported in rats (unspecified gender) that inhaled CCl4.55,143,144
Concerning the potential human exposure to CCl4 the primary routes mentioned are inhalation, ingestion, and dermal contact through air and water. 55
It is widely accepted that the cytochrome P450 isozymes are involved in this halomethane toxic effect. Bioactivation of CCl4 results a trichloromethyl free radical that reacts with oxygen resulting in a toxic reactive trichloromethyl peroxy radical. 141 Cell membrane integrity is affected due to the resulting chain reaction triggered by the attack of these highly energetic radicals on the polyunsaturated fatty acids of the cell membrane lipids. Cell energy processes and protein synthesis are also disturbed by the free radicals.19,141 According to Avasarala et al., 141 an inflammatory response is induced by Kupffer and stellate cells, as a result of the release of cytokines, chemokines, and other proinflammatory factors. The direct cytotoxic effect caused by these factors are also strengthen with the activation and recruitment of monocytes, neutrophils and lymphocytes. Heindryckx et al. 19 stressed fibrosis as a consequence of the repeated cycle of injury, inflammation, and repair, leading eventually to HCC.
PB (CAS No. 50-06-6; Group 2B; 54 NTP not available)
PB is a drug with sedative, hypnotic, and anti-epileptic proprieties. 145 Studies on cancer in experimental animals emphasize the carcinogenic risk involving long-term PB exposure in mice and rats after oral administration (drinking water and diet). 145 IARC selected several well-conducted studies involving long-term oral administration of PB; strains, adequate numbers of animals, dosage, and protocol duration were taken under consideration to analyze the carcinogenicity risk of this compound. Mice strains (e.g. C3H, CF-1, BALB/c, B6C3F1, and c-myc) and rat strains (e.g. Wistar, F344, and Sprague-Dawley) showed liver adenomas and carcinomas either after oral exposure or after injection administration. Age and gender were also taken under consideration. In mice, for instance, hepatocarcinogenesis enhancement by PB depends on the strain susceptibility, gender, age at the start of exposure, and type of initiator used.145–148 The incidence of HCC in weaning and/or male gender was higher (Table 2). 145 However, several studies have displayed that oral administration of PB resulted not only in tumor promoting but also in inhibition in mouse and rat liver, when associated with known carcinogens and modifying agents, depending on the carcinogen and time of administration. Nevertheless, DEN/PB-induced HCC is one of the most widely used initiator-promoter in experimental models to study hepatocarcinogenesis. 148 Likewise DEN, 2-AAF, and NMOR have also been used as initiator agents, followed by the association of PB in drinking water and diet to promote preneoplastic and neoplastic liver lesions in mice, rats, and hamsters. 145
Yang and Wang 149 highlighted the role of the nuclear receptor superfamily, named constitutive androstane receptor (CAR, NR1I3), in the regulation of drug metabolism and cancer development. CAR is activated by a restricted set of chemical carcinogens, including PB inducing the hepatic cytochrome P450 (CYP) 2B gene family in the liver.149,150 Prolonged PB treatment in mice induces CAR for gene expression changes, hepatomegaly, and liver tumor formation. 151
Syngeneic and xenograft models
Presently, the access to animal and human HCC cell lines has broadened the technical skills of HCC researchers. Genetic and malignant cell transformation processes, metastatic cell potential, cell response to external stimuli, and its influence in signaling mediation of the pathways involved represent an opportunity to study cancer in vitro. 34 Nevertheless, when using cell cultures, the interaction between neoplastic cells and the surrounding microenvironment may be lost. Recent advances in tissue culture techniques (e.g. 3D culture systems) are progressively circumventing this limitation since they provide promising in vitro models, allowing the study of cellular responses in a setting that resembles in vivo environments. 152 Understanding the tumor evolution is often challenging, due to the heterogeneity nature of the cell lines. 153 As a consequence of the tumor cell heterogeneity, when screening new anticancer drugs, it is essential to use multiple cell lines in xenograft models, in order to prevent misleading results.19,24,154 Cell lines used in transplantable HCC mice models are described in Table 3.24,34,38
Synopsis of the most used cell lines in xenografts and syngeneic HCC models; the present table does not intend to be an exhaustive cell line list.

AFP+: presence of α-fetoprotein; AFP−: absence of α-fetoprotein; HBV: donor infected with hepatitis B virus.
To perform a transplantable model, immunocompetent and immunodeficient animals can be used. The syngeneic models comprise immunocompetent mice carrying tumor cells or fragments originated in the same strain as a result of spontaneous causes, hepatocarcinogen, and viral infections.34,38,39,177
The discovery of nude (nu/nu) athymic mice and severe combined immunodeficient mice—SCID (scid/scid), that is, T-cell deficient and B + T-cell deficient, respectively, endorsed proficiency into human tissues transplantation in this species—the xenograft models (Figure 2), in order to avoid cell and tissue rejections.177–179 Both models are established by the implantation of fragments of human tumors or cultured human or other species cancer cells, either under the skin (ectopic) or into the organ of tumor origin (orthotopic).34,180–182
To reproduce the tumor microenvironment and organ tropism, an orthotopic model is better than the ectopic model; the advantages and disadvantages of both models are summarized in Table 3. If the influence of the immune system is required to study the tumor–host interactions or test anticancer drugs that influence the immune system activation, the syngeneic orthotopic mice model should be the right option. Nevertheless, creating high-quality orthotopic models is technically more puzzling than the construction of ectopic models.24,34,41 As mentioned above, depending on the research purposes, the choice of the implant anatomical site is crucial to establish if the tumor grows as a local nodule (subcutaneous and orthotopic) or if it will be disseminated since the beginning of the experiment as a result of the injections of cancer cells suspension (intraportal or intrasplenic).38,183
Due to the influence of the 3R’s philosophy—replacement, refinement, and reduction184–186—and taking under consideration the need for anticancer drugs screening, an efficient method was established by Hollingshead et al. 187 —the Hollow Fiber Assay (HFA). This method developed at the National Cancer Institute has the advantage, compared to traditional tumor xenograft, of having a minimal influence on the animal welfare; it also allows the uncontaminated retrieval of tumor cells for subsequent analysis.19,187 The HFA assay (Figure 4) allows the implantation of multiple polyvinylidene fluoride fibers, either subcutaneously or intraperitoneally, at once in a single nude mouse, consequently reducing the number of laboratory animals without compromising the reliability of the results.19,188 Tumor cell lines are positioned into semi-permeable hollow polyvinylidene fluoride fibers—1 mm inner diameter; after heat-sealed and cut, the fibers are placed into an in vitro culture medium for 24–48 h.19,188 After mouse recovery from surgery (3–4 days), drug-screening assay may be performed. After a period of 3–4 days of drug treatment, the HFA is surgically removed and surviving cells are retrieved for analysis to determine anticancer drug efficacy.19,189 According to Suggitt et al., 188 the HFA may offer an amenable, rapid, cost-effective, and ethically acceptable model to investigate unlimited drug–target interaction/target inhibition pre-clinically (Table 4).

Schematic representation of the HFA model. (●, *, and + represent different cell lines). Human or mice tumor cell lines are placed into semi-permeable polyvinylidene fluoride fibers (1 mm internal diameter). Fibers are then heat-sealed in 2 cm intervals and subsequently cultured in vitro (standard tissue culture conditions) for 24–48 h, preceding subcutaneous or intraperitoneal implantation. Since the cell lines are isolated in the hollow fibers, it is possible to implant several different tumor cell lines at the same time into a single mouse. After drug-screening assay, the cell lines can be retrieved for analysis.

Viral models
Despite the preventive strategies against HBV and HCV, as recommended by World Health Organization (WHO), the high prevalence of these hepatotropic viruses is implicated in HCC etiology and its asymmetric distribution worldwide.1,38 In some Asian regions and sub-Saharan Africa, 8% of the population is chronically infected with HBV; this infection is responsible for nearly 60% of total liver cancers in developing countries while HCV accounts for 33% versus 23% (HBV) and 20% (HCV) in developed countries.1,8 As largely discussed in the literature, HBV and HCV are causative agents of HCC due to the chronic inflammation and regeneration of hepatocytes that increase the odds of DNA damage and consequent alterations associated with cancer development.190,191 Although the progress achieved in recent years, the absence of feasible small animal model of human HCV infection has been regarded as an obstacle for further knowledge concerning the virus life cycle and development of effective therapeutics.192,193
Concerning the animal models reported in current literature related to HBV-induced HCC, woodchucks (Marmota monax) and ground squirrels (Spermophilus beecheyi), chronically infected, respectively, with woodchuck hepatitis virus (WHV) and ground squirrel hepatitis virus (GSHV), are mentioned as reliable models.34,194 Some human hepatitis virus-infected chimpanzee (Pan troglodytes), tree shrews (Tupaia belangeri chinensis), and rhesus macaques (Macacca mulatta) also developed HCC mainly when combined with chemical carcinogens.34,195,196 Concerning rhesus macaques, neoplasia in juvenile is extremely uncommon. A recent case report of naturally occurring neoplasia and metastatic liver was identified in a 3.75-year-old primiparous female rhesus macaque. 197 Due to its scarcity and clinical history, this case reflects similitude to that of human adolescents with HCC.
Regarding the animal models related to HCV-induced HCC, chimpanzees (P. troglodytes) and tree shrews (T. belangeri chinensis) are also presented in current literature.192,198 HCV infects only chimpanzees and humans, which makes chimpanzees a natural choice as animal model to study the virus pathogenicity. However, due to availability and economic and ethical constraints, this animal model is considered unacceptable for scientific research.192,199 Concerning Tupaias, they are susceptible to be infected in vivo with HCV and immunosuppression, caused after whole-body radiation, increased HCV replication.193,198 Tree shrews are also infected with human HBV in vivo, resulting in viral DNA replication and gene expression in the livers. 193
The WHV belongs to the hepadnaviruses family, which also includes the GSHV, duck hepatitis B virus (DHBV), and human hepatitis virus (HBV).193,200 The WHV induces the formation of foci of altered hepatocytes, which ultimately will lead to well-differentiated HCC. 34 Despite the resemblance in liver inflammation, injury, and repair processes as observed in HBV-positive patients, important differences should be noted. WHV DNA is integrated into hepatocyte DNA within or nearly contiguous to a specific locus associated with overexpression of c-myc and N-myc genes.34,38,97 However, it should be noticed that the insertion of activated dominant oncogene is not a typical mechanism for HBV carcinogenesis.
GEM model
The first report concerning the technology for generating transgenic mice carrying cloned oncogenes was published in 1984; following reports (1985–1987) supported the hypothesis that oncogene expression in normal cells within normal tissues of a mammalian organism might lead to tumor development. 205 This development expanded the possibilities to study the molecular features of human malignancies in vivo, namely the role of a specific gene and the interaction of different genes such as oncogenes and tumor-suppressor genes (TSGs) related to HCC development. 24 Tracking the possibility of creating adequate mice model for the study of HBV infection in liver cancer development, Kim et al. 201 placed the HBx (hepatitis B virus X) regulatory gene directly into the germ line of mice. The HBx protein affected the host gene expression by means of transcriptional transactivator of viral genes, leading ultimately to HCC development. However, despite the development of chronic hepatitis and liver cancer, some limitations may be noticed, due to differences in the establishment of this transgenic mouse model and the situation of infection with the intact HBV (Dane) particles. 193
The pursuit of an animal model for HBV and HCV infection in genetically altered mice led to advances of human hepatocyte chimeric mice. As a result of the work of Degen and Palmiter, the AL-uPA (Albumin enhancer/promoter urokinase-type plasminogen activator) transgenic mice line was established in Ralph Brinster’s laboratory.202,203 These transgenic mice show accelerated hepatocyte death with consequent chronic hepatocyte growth stimulation. 202 Recently, Chayama et al. 190 drew the attention to the development of human hepatocyte chimeric mice and hepatitis virus infection model. The uPA/scid mice were created by crossing uPA transgenic mice and scid mice. 190 A similar chimeric mice model was generated by Mercer and co-workers for studying the human HCV in vivo. 204 Despite being useful for the assessment of efficacy of anti-HBV agents, the chimeric mice models are relatively complex, expensive, and have no immune response derived from uPA/scid mouse. 199
GEMs are designed to mimic the pathophysiological and molecular features of human malignancies, namely, HCC.206,207 Apart from their complexity, GEM can be summarized, according to Frese and Tuveson, 207 either as transgenic or as endogenous (Table 5).
Summary of the transgenic and endogenous mouse models of cancer according to Frese and Tuveson. 207

GEM: genetically engineered mouse model; TSG: tumor-suppressor genes.
In HCC, likewise other cancer types, the analysis of the genetic variations in the pathways involved in tumor growth, angiogenesis, proliferation and apoptosis are crucial to accompany the development in its microenvironment.24,208 Gene targeting technology has provided knockout models, which are used in loss-of-function studies 209 or conditional GEM, that rely on the use of site-specific recombinase systems in order to control the spatiotemporal gene expression in a restricted manner. 207 In studies concerning allele gain-of-function, conditional transgenic and knockin is the most preferable approach. 209
Despite the fact that the hepatocarcinogenesis genetic events are not entirely understood, evidence point to several pathways involved in the process, namely, p53, Rb, and Wnt/β-catenin. 24 The use of transgenic SV40 T-Ag (Simian Virus 40 T-antigen) has been extensively described.210–213 SV40 was one of the first DNA viruses discovered with tumorigenic properties and cell transforming capabilities. 213 The genome of SV40 encodes two oncogenic proteins, large and small T-antigen (T-Ag and tAg, respectively),210,212 which play a key role in inactivating the tumor suppressor genes p53 and Rb and cell regulatory proteins.24,212
Murakami et al. 214 describe another transgenic mouse model, taking under consideration the deregulation of c-myc expression detected in primary liver tumors and induced HCC in rodents and the frequent coexpression of the transforming growth factor alpha (TGF-α) observed in both human tumors and in transformed liver cultured cells.24,34,214 Liver tumors occur in c-myc transgenic mice as a result of the selective expression of c-myc in the liver directed by the albumin enhancer/promoter or the α-1-antitrypsin promoter.24,214 This c-myc/TGF-α synergistic effect was demonstrated by Santoni-Rugiu et al. 215 resulting in a major boost of neoplastic development compared with the expression of these isolated transgenes. Furthermore, in c-myc/TGF-α transgenic mice, the TGF-βRI was reduced in a subgroup of undifferentiated HCC, which may be implied in dedifferentiation and acquirement of high-grade malignancy. 34
Nevertheless the fact of the importance of these models to study the role of specific genes in hepatocarcinogenesis, some limitations have been pointed out related to the presence of the mutations during embryogenesis, compensatory molecular pathways which might be activated, and the expression of the transgene in all hepatocytes. 24 To circumvent the above-mentioned limitations, the advent of the conditional mice models seems to be an effective alternative in the generation of cancer-prone GEM. 207
Somatic mutations in specific tissue can be induced and spatiotemporally controlled using conditional and the inducible systems. According to Cheon and Orsulic 209 the most common systems are as follows: the Cre-loxP system; the flippase (FLP)-flippase recognition target (FRT) system; the inducible Cre system; and the tetracycline (tet)-inducible system. Most recently, a combination of these technologies has been used to more accurately mimic tumor onset and its progression. 209
Conclusion
It is clear that the availability of a wide number of mice models to study HCC gave researchers the opportunity to further expand the knowledge of the biopathology signaling pathways by assessing novel therapeutic approaches and genetic manipulation methods. The variety of animal models of HCC, namely, rodents, such as mouse and rat, will continue to contribute to understand the mechanisms underlying liver carcinogenesis. Tumor progression, tumor–host microenvironment, mechanisms that trigger metastatic dissemination, immune system response, and the development of new therapeutic strategies will continue to focus on the researchers’ attention and their investigation purposes. At present time, there is no single animal model which is capable of recapitulating all the genetic and biological aspects of the human cancer. Based on this assumption, knowing the limitations and strengths of the existing animal models will allow to emphasize their potential in precocious detections of human tumors and treatment, thus reducing health care expenses and providing human comfort and welfare. Future trends in HCC research will be certainly related to personalized gene-based treatments capable of offering an accurate intervention in the key pathways and mediators concerning HCC.
Although the advent of GEM, molecular medicine and cell culture recent achievements, (due to the fact of being less expensive, not requiring technically demanding laboratory, mimicking human genomic instability, namely, gene mutation, activation of certain pathways and consequent protein expression/gene regulation), the chemically induced models will continue to provide usefulness evidence concerning HCC research, unveiling its complexity.
Footnotes
Declaration of conflicting interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was partially supported by a PhD Grant of the Foundation for Science and Technology (FCT) Portuguese Ministry of Education and Science (SFRH/BD/60280/2009).