
Abstract
Dendritic cells are important in initiating immune responses; therefore, a range of dendritic cell–based approaches have been established to induce immune response against cancer cells. However, the presence of immunosuppressive mediators such as adenosine in the tumor microenvironment reduces the efficacy of dendritic cell–based cancer immunotherapy. In this study, we investigated whether blockade of the A2A adenosine receptor with a selective antagonist and a CD73 inhibitor may increase the efficacy of a dendritic cell–based cancer vaccine. According to the findings, this therapeutic combination reduced tumor growth, prolonged survival of tumor-bearing mice, and enhanced specific antitumor immune responses. Thus, we suggest that targeting cancer-derived adenosine improves the outcomes of dendritic cell–based cancer immunotherapy.
Introduction
There is substantial evidence that inflammation is a critical component of tumor progression.1,2 Adenosine is an inflammatory mediator that is produced in the conditions of hypoxia, ischemia, inflammation, and trauma and serves as a danger signal.3,4 Adenosine is present in high concentrations in cancerous tissues and acts as a crucial mediator in the alteration of immune cell functions.5–7 Cellular signaling by adenosine occurs through a family of G-protein-coupled cell-surface receptors including A1, A2A, A2B, and A3. Signaling through the A2A receptor (A2AR) leads to the stimulation of adenyl cyclase and increases intracellular levels of cyclic adenosine monophosphate (cAMP).8,9 The intracellular cAMP is known to be a highly immunosuppressive mediator.3,10 Several studies also support the idea that adenosine stimulates the production of vascular endothelial growth factor (VEGF) and therefore supports tumor angiogenesis.11–13
Adenosine in the tumor milieu is generated from extracellular nucleotides by ectonucleotidases such as CD39 and CD73. CD39 catalyzes the degradation of adenosine triphosphate (ATP) into AMP, and CD73 subsequently hydrolyzes AMP into adenosine.14,15 High levels of CD39 and CD73 have been reported in several solid tumors in humans.16,17 Elevated expression of these molecules is also associated with poor outcomes in breast cancer patients.18,19 It has been shown that targeting the CD73–adenosine pathway provides beneficial outcomes for cancer patients. 20 Besides, CD73-gene-silencing or interfering with CD73 activity in tumor cells enhances antitumor T-cell response and inhibits tumor growth.21–23 Moreover, immunosuppressive immune cells such as regulatory T (Treg) cells and myeloid-derived suppressor cells (MDSCs) express high levels of the enzyme ecto-5-nucleotide CD73.24,25
Dendritic cells (DCs) as professional antigen (Ag)-presenting cells (APCs) are capable of directing Ag-specific immune responses; 26 therefore, the use of DCs has become a promising method for cancer immunotherapy.27–29 The purpose of DC vaccination is to induce tumor-specific effector T cells with immunological memory in order to control tumor recurrence.29,30 To improve the outcomes, DC vaccines need to be combined with other therapies that target immunosuppressive mechanisms by which tumor cells escape T-cell responses.31,32 In this study, we targeted A2AR and CD73 in conjunction with DC therapy in a mouse breast tumor model. We demonstrated for the first time, to our knowledge, that this combination maybe exploited to induce tumor regression and increase survival. Targeting A2AR and CD73 improved the efficacy of DC vaccines via the induction of tumor-specific T-cell activity.
Methods and materials
Mice, cell line, and reagents
Six- to eight-week-old female BALB/c mice were purchased from the Lab Animal Center, Pasteur Institute of Iran. They were housed in similar conditions with a 12-h dark/light cycle. BALB/c-derived breast carcinoma (4T1) cell lines were cultured in RPMI-1640 (Biosera, UK) supplemented with 10% heat-inactivated fetal bovine serum (Gibco, Grand Island, USA), 2 mM
Preparation of tumor lysate
The 4T1 cells were suspended at 4 × 107 cells/ml in phosphate buffered saline (PBS) were subjected to 6–7 cycles of rapid freezing in liquid nitrogen and thawing at 37°C. The disrupted cells were centrifuged at 1500 r/min for 15 min to remove particulate cellular debris. The protein concentration of the lysate was determined by BCA Protein Assay Kit (Thermo Fisher Scientific, USA).
Generation of bone marrow–derived DCs
Bone marrow–derived DCs (BMDCs) were generated according to the Inaba et al. Protocol
33
with some modifications. In brief, bone marrow cells were flushed from BALB/c femurs and tibia. Contaminating erythrocytes were lysed with 0.8% NH4Cl in an aqueous solution. Bone marrow cells (5 × 105 cells/mL) seeded in 24-well plates in RPMI-1640 supplemented with 10% heat-inactivated fetal calf serum 2 mM
Tumor challenge and treatment
Tumor cells (7 × 105 4T1 in 200 µL PBS) were injected subcutaneously in the right flanks of the mice. At days 5, 7, 9, 12, 14, and 16, SCH58261 (1 mg/kg) and/or APCP (20 mg/kg) or PBS alone (200 µl) were administered intraperitoneally. On day 6, BMDCs (106) were injected around the tumor site. When the tumor was palpable, the shortest and longest surface diameters were measured every 2 days by digital calipers; the tumor area was calculated through multiplying the shortest and longest axes. One day after the last injection, 3 out of 8 mice of each group were killed by cervical dislocation for evaluation. The remaining mice were sacrificed when the tumor diameters reached 400 mm 2 .
Flow cytometry
The following conjugated antibodies (Abs) were used in flow cytometry: APC-conjugated anti-Gr-1, phycoerythrin (PE)-conjugated anti-CD11b, APC-conjugated anti-CD4, APC-conjugated anti-CD3, and Alexa Fluor 488-conjugated anti-Foxp3 (all obtained from BioLegend, USA). For Foxp3 intracellular staining, after cell surface staining, CD4+T cells were processed using anti-mouse Foxp3 Ab according to the manufacturer’s instructions. To determine the phenotypes of the cultured DCs, cells from the seventh day cultures were stained with monoclonal Abs (mAbs) against cell surface molecules such as CD86, CD80, CD40, MHC-II, and CD11c. Flow cytometry was conducted using a FACS Calibur (Becton-Dickenson, USA) and analyzed with FlowJo software. In all experiments, isotype controls with the same immunoglobulin (Ig) class were included.
ELISA
To assess the production of cytokines and granzyme B (GrB), splenocytes from BALB/c mice with 4T1 tumors were depleted of red blood cells; then the cells were incubated at 106 cells/well with 80 µg/mL tumor antigen lysate in a complete culture medium at 37°C and 5% CO2 for 72 h (cytokine measurement) or 78 h (GrB measurement) in a total volume of 1 mL. Supernatants were evaluated using ELISA kits for GrB (eBioscience), interferon (IFN)-γ, IL-10, VEGF, and cAMP (R&D Systems).
Analyzing lymphocyte proliferation using measurement of carboxyfluorescein succinimidyl ester dye dilution
Carboxyfluorescein succinimidyl ester (CFSE)-labeled splenocyte cells (2 × 105/well) were cultured in the presence of 80 µg/mL tumor antigen in 96-well plates. After 120 h, the cells were washed and stained with anti-CD3 antibody for 30 min and then analyzed with FACS Calibur.
Analysis of gene expression by real-time polymerase chain reaction
Total RNA was isolated from tumor tissues using Hybrid-R RNA purification kit (Gene All Biotechnology, Korea). cDNA was prepared using QuantiTect reverse transcription kit (Qiagen). The expression of VEGF and IL-10 were quantified by real-time polymerase chain reaction (PCR) using SYBR Green Master Mix (Primer Design, UK) and ABI 7500 detection system (Applied Biosystems). Relative mRNA levels were determined using the ΔCt method. Values were expressed relative to endogenous β-actin expression.
Statistical analysis
Statistical differences were analyzed by one-way analysis of variance (ANOVA) with a significance level of P < 0.05. Kaplan–Meier survival curves were analyzed for statistical difference between groups using the log-rank test. Statistical analysis and graph generation were conducted using GraphPad Prism (Version 5).
Results
Phenotype of BMDCs
In order to determine the maturation status of BMDCs before injection, the expression of MHC-II and co-stimulatory molecules (CMs) were analyzed by flow cytometry. Approximately 70% of mature DCs expressed mouse-specific DC marker CD11c and also mature DCs expressed a higher level of MHC-II and CMs as compared with immature DCs (Figure 1).
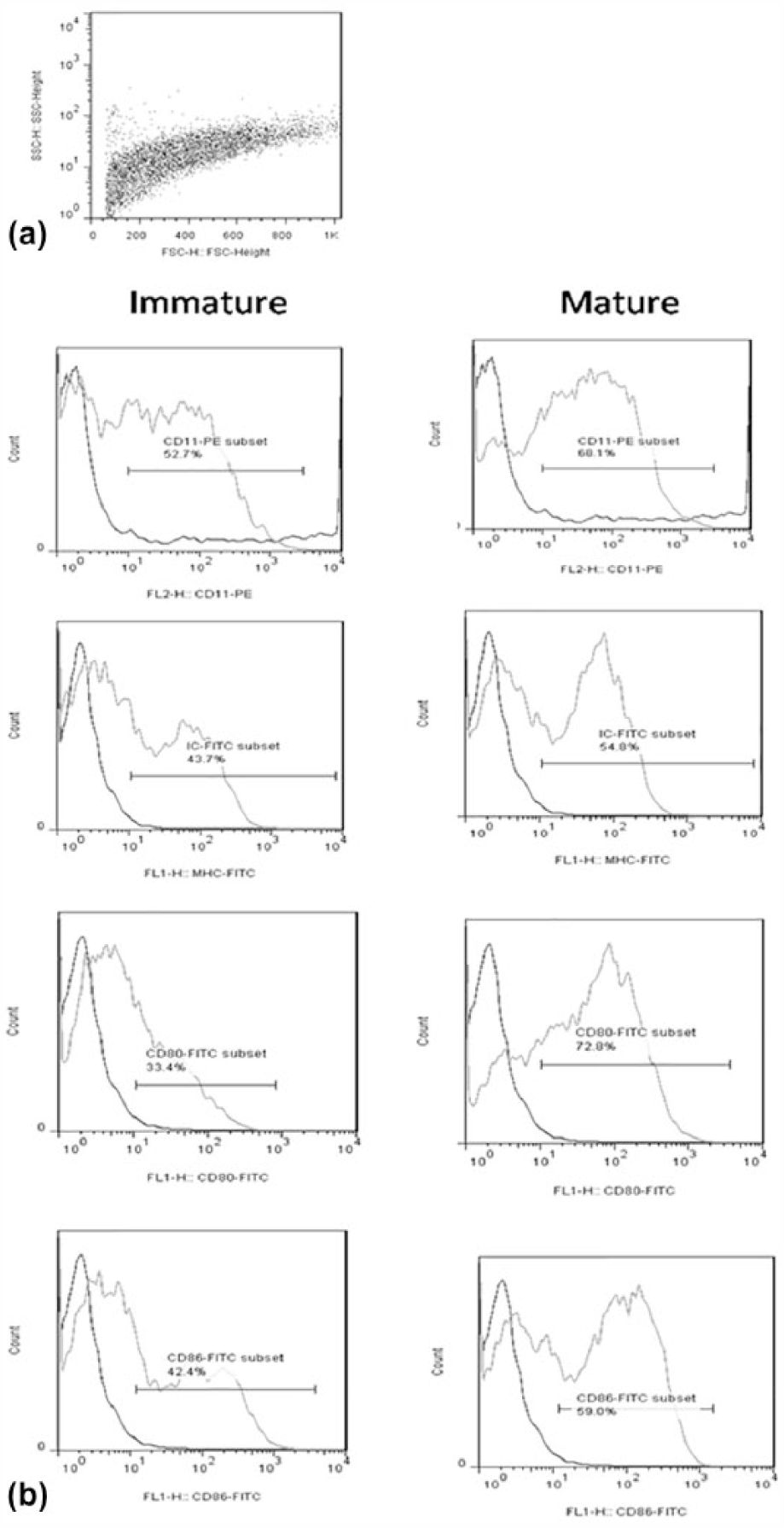
Phenotype of Dendritic cells on day 7. (a) Forward scatter (FSC) versus side scatter (SSC) dot plots of differentiated DCs. (b) Expression of CD11c, CD86, CD80, CD40, and MHC-II molecules (gray line) compared with isotype control (black line) in immature(without LPS) and mature DC.
Antitumor effects of DC, SCH, and APCP administration
A schematic of the experimental procedure is shown in Figure 2(a). To examine the antitumor effect of the SCH, APCP, and DC combination therapy, tumor size and growth rate of mice were measured. Treatment of the established tumors with DC/APCP and DC/APCP/SCH induced a significant reduction of the tumor size compared with the control groups (Figure 2(b)). As shown in Figure 2(c), treatment with DC/APCP and DC/APCP/SCH significantly improved survival rates compared with control groups.

Antitumor effects of DC, SCH, and APCP administration. (a) Schematic diagram of experiments. BALB/c mice bearing 4T1 tumors were left untreated or, on days 5, 7, 9, 12, 14, and 16, received SCH, APCP, and/or DC vaccine (106 cells, subcutaneously around the tumor site, on day 6 of tumor inoculation) alone or in combination. (b) Mean ± SEM of eight mice per group is shown (n = 8, P < 0.05 by one-way ANOVA). (c) Survival of the animals in each group was monitored and the respective Kaplan–Maier curves are given (n = 5; *significant at P < 0.05).
Improvement of tumor-specific immune responses by administration of DC, APCP, and SCH combination therapy
To address whether DC therapy combined with SCH and APCP treatment could stimulate a cytotoxic T lymphocyte (CTL) response, splenocytes were isolated from mice one day after the final injection, and their cytotoxic activity against 4T1 tumor cells was determined by granzyme measurement. As shown in Figure 3(a), DC therapy combined with SCH and APCP resulted in a significant elevation of granzyme level compared with the control group and all instances of single administration. To investigate the type of generated immune response, IFN-γ and IL-10 level were determined. The combination of DC with both drugs significantly increased the production of IFN-γ (Figure 3(b)) and reduced IL-10 (Figure 3(c)), although the reduction of the latter was not significant.
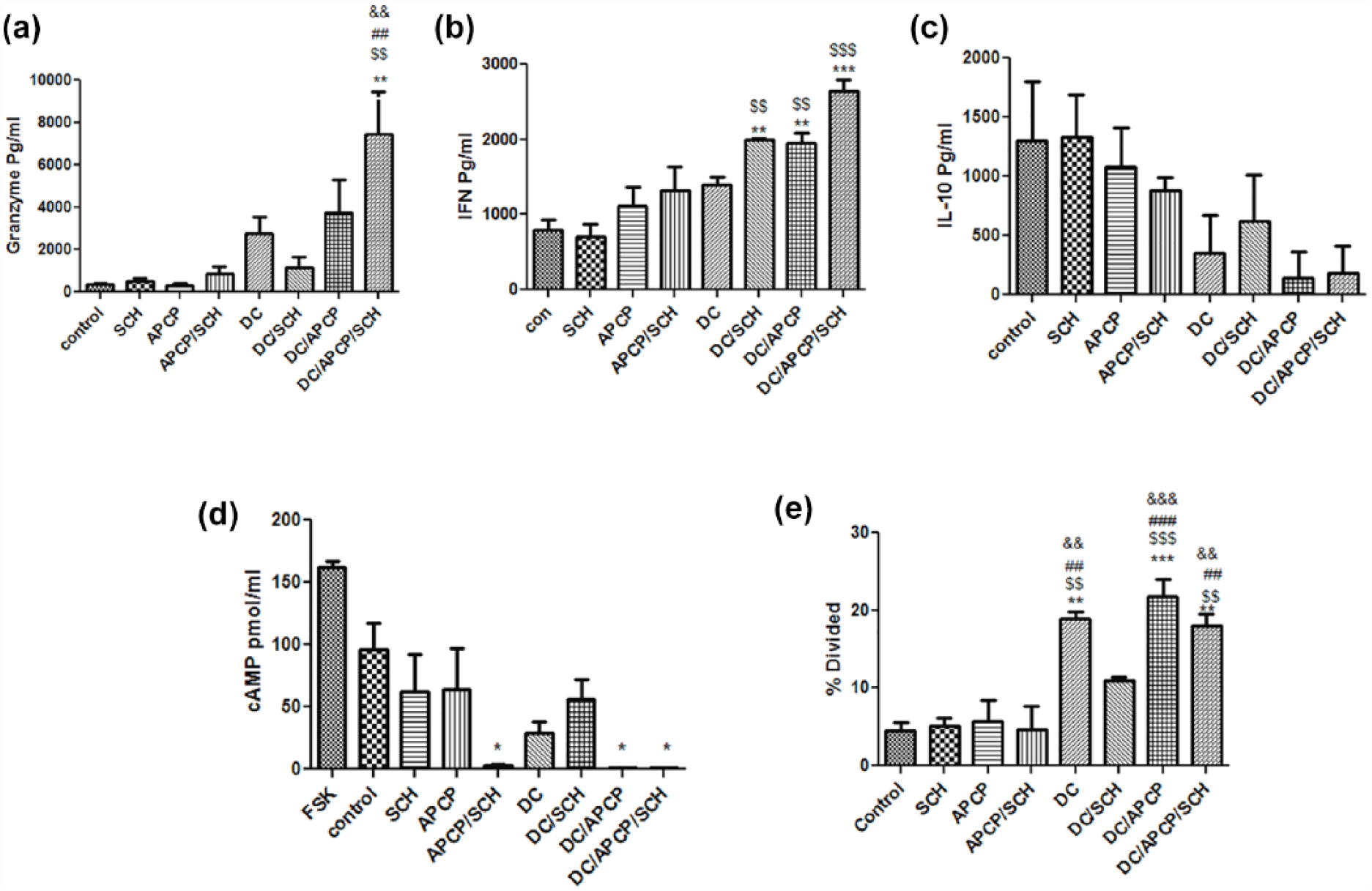
Tumor-specific immune responses induced by administration of combination therapy. Splenocytes obtained from treated mice were cultured with tumor Ag for 78 h, and then supernatants were collected and the concentration of (a) granzyme, (b) IFN-γ, (c) IL-10, (d) cAMP were determined. Forskolin was used as a positive control because it increases cAMP levels and (e) 2 × 106 CFSE-labeled splenocytes isolated from tumor-bearing mice were treated in vitro with tumor lysate in CM (80 µg/ml, 5 d) and the proliferation ability of CD3-stained cells, reported as percentage of proliferation, was evaluated by flow cytometry. Bars indicate mean ± SEM. Average values from three different experiments have been shown
As stated earlier, adenosine A2 receptors stimulate cAMP production, a known immunosuppressive mediator. 34 To demonstrate whether the inhibition of the CD73–adenosine pathway reduces cAMP production, we cultured splenocytes of treated mice with tumor Ag and then cAMP levels were determined. As seen in Figure 3(d), cAMP generation was sharply reduced in groups receiving DC/APCP, DC/APCP/SCH, or DC/SCH.
To assess the effect of the treatments on specific T-cell responses, CFSE-labeled splenocytes of treated mice were cultured with tumor Ag, and the CFSE dilution was determined. As shown in Figure 3(e), mice treated with DC vaccine (with the exception of DC/SCH) showed an augmented proliferation response compared to other groups.
Effect of combination therapy on the number of MDSCs and T cells
In previous studies, T cells and MDSCs were shown to express CD73 and CD39, enabling these cells to generate adenosine from ATP;24,25 therefore, we analyzed the frequency of these cells in the spleens of treated mice. We observed that combination therapy increased the number of CD11b+ Gr-1+ and FOXP3+ CD4+ cells. However, this elevation did not reach statistical significance (Figure 4(a) and (b)).

Combination therapy showed no effect on the number of splenic MDSCs and Treg cells. The number of (a) CD11b+ Gr1-PE+ cells and (b) FOXP3+ CD4+ cells were assessed by flow cytometry in the spleens of mice. Bars indicate mean ± SEMs. Average values from three different experiments are shown.
Administration of combination therapy with DC, APCP, and SCH is associated with decreased VEGF production
VEGF is a key mediator of tumor angiogenesis. Splenocytes from treated mice were isolated and VEGF levels were measured in response to stimulation with tumor Ag. Figure 5(a) shows that in mice that received DC/APCP/SCH alone, and those receiving a combination of SCH and APCP, cells produced lower levels of VEGF compared to control mice.

Combination therapy decreased VEGF production. (a) Splenocytes obtained from treated mice were cultured with tumor antigen. On day 3, supernatants were collected and the concentration of VEGF was determined. (b) For assessing gene expression of VEGF in the tumor tissues, specific mRNA transcripts were quantified by real-time PCR. Bars indicate mean ± SEMs. Average values from three different experiments are shown.
Real-time PCR analysis also showed that the gene expression levels of VEGF in tumor tissues of mice treated with DC/APCP, DC/APCP/SCH, and SCH/APCP were lower than in other groups (Figure 5(b)).
Discussion
Immunotherapy is a type of cancer treatment that attempts to improve the ability of an individual’s immune response to reject the tumor immunologically. 35 However, the production of immunosuppressive factors such as adenosine in the tumor microenvironment reduces the efficacy of this method. Therapies that target immunosuppressive mechanisms are effective tools that enhance anticancer immune responses, leading to better immune surveillance.32,36
We combined DC immunotherapy with an adenosine receptor antagonist, and a CD73 inhibitor, as a novel combination strategy in mouse breast tumor model (4T1). It has been shown that the 4T1 cell line, which shows behavior comparable with triple negative breast cancer in humans, expresses high amounts of CD73. Previous experiments showed that adoptive transfer of CD8+T cells combined with ZM241,385—the A2 receptor antagonist—led to a reduction in the number of metastatic nodules in CL8-1 melanoma model. 37 In a study conducted by Jin et al., 38 blockade of the A2A adenosine receptor with the antagonists SCH58261 and caffeine promoted tumor-specific immunity and improved the efficacy of adoptive T-cell therapy. A combination of SCH58261 and adoptive T-cell therapy was also shown to significantly improve survival in tumor-bearing mice. 38 The aim of A2A adenosine receptor blockade is to interfere with adenosine receptor signaling in the tumor environment in order to induce tumor-specific immune responses. In this study, a combination of the CD73 inhibitor APCP and the adenosine receptor antagonist SCH with DC injection led to delays in tumor growth and improved survival in tumor-implanted mice. In addition, this combination induced specific immune responses that likely cause the inhibition of tumor growth.
Tregs are potent immunosuppressive cells that induce the progression of cancer through their ability to inhibit antitumor immunity and provoke angiogenesis. 39 Different drugs developed for other therapeutic indications (such as anti-angiogenic factor and tyrosine kinase inhibitors) have recently been used to inhibit Tregs. 40 Tregs have been shown to express CD73 and CD39, and therefore, they actively produce extracellular adenosine and block the activation of effector cells through A2AR.41–43 In addition, MDSCs prevent immune responses by releasing adenosine in a CD73-dependent manner. The blockade of adenosine receptors, in previous studies, led to reductions in tumor growth and immune suppression mediated by MDSCs in a mouse model of melanoma.25,44 In our study, the administration of an adenosine antagonist or CD73 inhibitor as single agents or in combination with DC therapy did not change the number of Treg and MDSC populations. It seems that inhibition of adenosine in the tumor model does not alter the numbers of these suppressive cells. Because of interference with adenosine suppression, it seems likely that the inhibitory functions of these cell types are impaired. In this experiment, production of IFN-γ by splenocytes co-cultured with tumor antigen significantly increased in groups treated with DC, SCH, or APCP, individually and in combination. Moreover, the reduction of IL-10 was observed in DC vaccine-treated mice. Inhibition of cAMP occurred in mice treated with DC/APCP and DC/APCP/SCH. Moreover, an analysis of CTL function and the proliferative capacity of specific T cells in mice that received DC/APCP/SCH showed an augmented response compared with other groups.
VEGF acts as a key mediator of angiogenesis in cancer. Several studies have demonstrated that adenosine enhances VEGF expression.45–47 Novitskiy et al. showed that adenosine-differentiated DCs express high levels of angiogenic, proinflammatory, immunosuppressive, and tolerogenic mediators, including VEGF, IL-8, IL-6, IL-10, cyclooxygenase-2 (COX-2), tumor growth factor (TGF)-β, and indoleamine-pyrrole 2,3-dioxygenase (IDO). 48
Another study showed that CD73 causes endothelial cells to form tube-like structures and facilitates tumor angiogenesis. 11 It has been shown that targeted blockade of CD73 with a mAb significantly decreased tumor VEGF levels and suppressed tumor angiogenesis. 49 In our study, inhibition of adenosine combined with a DC-based vaccine reduced VEGF production. Interestingly, the combination of SCH and APCP had an additive effect.
Altogether, it seems that inhibition of CD73 activity, both individually and, especially, in combination with an adenosine receptor antagonist, enhances the potency of DC vaccines to induce more robust antitumor immune responses, facilitates retarded tumor growth, and prolongs the survival of tumor-bearing mice. This approach could be used for DC-based antitumor vaccines, and could be evaluated in clinical trials as a treatment for certain cancers in humans, particularly in cases where tumors express high amounts of surface CD73.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
This study was conducted in strict accordance with regulations formulated by the Institutional Animal Care and Use Committee. The protocol was approved by the Committee on Animal Experiments of Tehran University of Medical Sciences.
Funding
This study was supported by a grant from Tehran University of Medical Sciences (Grant number: 92-03-87-23034).