
Abstract
The sonic hedgehog signaling pathway is critical for cell proliferation, cell differentiation, and organ development during embryogenesis. Aberrant activation of the sonic hedgehog pathway has been associated with a variety of human cancers. Currently, there is no target molecular drug that clinically affects sonic hedgehog activation in patients with gastric cancer. In this review, we will focus on the current clinical treatment options for advanced gastric cancer and discuss the sonic hedgehog signaling pathway. We also review our understanding of the role of sonic hedgehog signaling in advanced gastric cancer progression. Finally, we will describe current known molecular pathways that crosstalk with the sonic hedgehog pathway in cancer stem cells.
Introduction
Gastric (stomach) cancer is a disease in which tumor cells form in a mucous membrane of the stomach. Nearly all gastric cancers are adenocarcinomas. The other types of gastric cancers are gastrointestinal stromal tumors, gastrointestinal carcinoid tumors, and lymphomas. Gastric cancer is typically diagnosed at an advanced stage or metastasis stage because there are no early symptoms or signs. Consequently, with over 800,000 deaths each year worldwide, gastric cancer is the second leading cause of cancer death.1,2 Despite some advances in treatment, the prognosis of patients with gastric cancer remains poor.3,4
As a component of the hedgehog (Hh) family, sonic hedgehog (Shh) was originally identified in Drosophila melanogaster, and the Shh signaling pathway is highly expressed in mammals.5,6 As shown in Figure 1, upon binding to its Patched (PTCH), Shh can increase the activity of the Smoothened (SMO) and the glioma-associated oncogene homolog (GLI) transcription factor to subsequently regulate the expression of target genes that control cell growth and survival in several tissues. 7 Numerous reports have shown that the Shh signaling pathway is overexpressed in various cancer types, including colon cancer, neuroblastoma, and lung cancer.8–15 In addition, recent data have indicated that the Shh pathway is correlated with the initiation of gastric cancers.16–18

Hedgehog signaling pathway. Inactivated signaling (left) occurs in the absence of Shh ligands, wherein PTCH inhibits SMO resulting in GLI1 sequestration in the cytoplasm by SuFu. In the presence of Shh (right), PTCH suppression of SMO is abrogated, resulting in the nuclear accumulation of GLI1 and activation of target genes that promote several oncogenic properties in tumor cells.
Recent data have supported the hypothesis that cancer stem cells (CSCs) exist in several solid tumors, including colorectal cancer, breast cancer, lung cancer, and gastric cancer. 19 CSCs have the potential for self-renewal as well as metastasis and recurrence.20,21 Various signaling makers and biomarkers have been applied to detect CSCs, including those with gastric cancer, prostate cancer, and breast cancer.19,22,23 Targeting CSCs has been suggested for treatment rather than traditional therapies because standard chemotherapy and radiotherapy target the differentiated tumor mass, resulting in cancer recurrence. 24 The identification of the CSC portion of a tumor may provide a new therapeutic option based on targeting in this small population. In this review, the characteristic properties of gastric cancer and gastric cancer stem cells (GCSCs) are reviewed with regard to surface markers and Shh signaling.
Core tip
Gastric cancer is the fifth most common malignancy worldwide with the highest incidence rates in Latin America and Eastern Asia. Although the incidence of gastric cancer has been decreasing, it is the second leading cause of cancer mortality; thus, there is a need for targeted therapy for more effective gastric cancer treatment. One of the pathways reported to be dysregulated is the Shh signaling pathway. This review focuses on how this aberrant activation of the Shh signaling pathway is crucial for gastric cancer tumorigenesis. In addition, we summarize the Shh signaling pathway inhibitors being investigated for gastric cancer treatment. Herein, we discuss that targeting of the Shh signaling pathways in gastric cancer or gastric cancer stem cells may be a promising approach for developing novel drugs to treat gastrointestinal cancer.
Characterization of Hh signaling pathway
Hh pathway was first identified in the Drosophila. 25 This pathway has major seven components which includes Hh, PTCH, SMO, GLIs, fused (Fu), Costal 2 (Cos 2), and suppressor of fused (SuFu). Also, Hh protein types are Shh, Indian hedgehog (Ihh), and desert hedgehog (Dhh) in mammals. 25 Hh targets the PTCH receptor (a 12-span transmembrane protein), and it acts as a central negative regulator of the Hh signaling pathway by suppressing the SMO in the absence of Hh protein. If Hh protein binds with the PTCH, SMO releases GLIs including GLI1, GLI2, and GLI3. And then, GLIs join the nucleus to regulate the transcription factors. It has been previously reported that GLI1 was amplified in human brain tumor.26–28 GLI target molecules have been found to affect cell proliferation,29–32 cell survival,30,33 epithelial–mesenchymal transition (EMT),34,35 and stem regeneration30,36 (Figure 2). If Hh is absent in the cells, GLI proteins become the GLI-Cos 2-Fu-SuFu complex and joins the cytoplasm. 37 And, GLIs separate from the nucleus and its target genes are activated. The other way, when Hh is present in the cell, Hh binds to PTCH and it acts as an activator to SMO. Consequently, GLIs become inactivated, discharged from the GLI-Cos 2-Fu-SuFu complex, and join the gene promoter sites to adjust the transcription factor–binding sites. However, recent studies represent that GLI can be regulated by Hh-independent mechanisms. For instance, the Hh pathway is modified by the RAS-MEK/AKT signaling pathway in cancer cells. 38
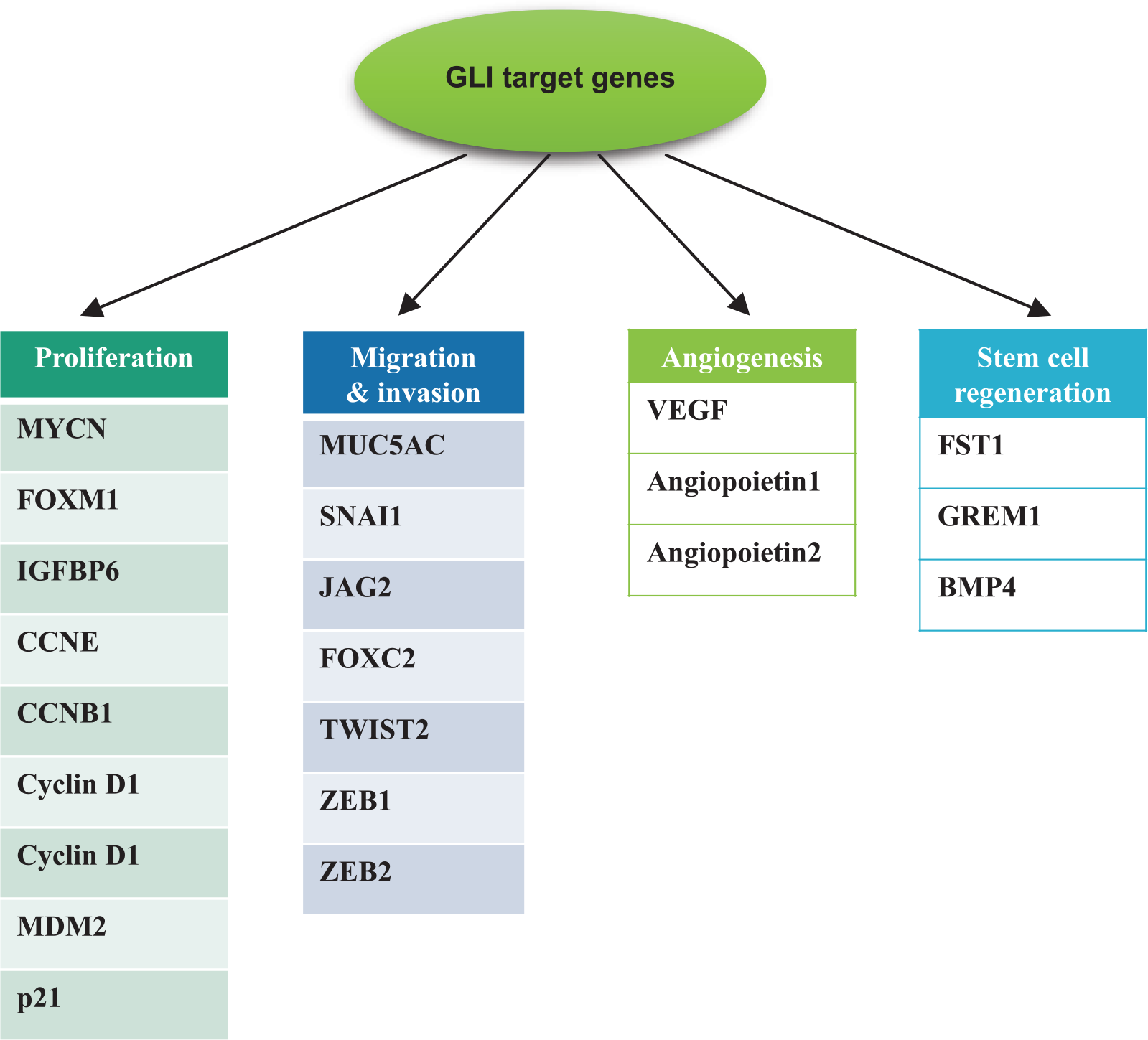
GLI target genes. Several GLI target genes have been identified to affect cell proliferation (MYCN, CCND1, CCND2, and FOXM1), migration and invasion (EMT) (FOXC2, SNAI1, and TWIST2), angiogenesis (VEGF, ANGP1, and ANGP2), stem regeneration (JAG2 and FST), and cell survival (BCL2 and CFLAR).
Hh in stomach cancer
The expression of Shh protein occurs in parietal cells of the normal adult gastric corpus (Table 1). However, there is controversy regarding how Hh signaling occurs. Most studies have shown that the gastric epithelium secretes Shh ligand and activates Hh receptors in the primitive mesenchyme. For instance, the physiology of mice for PTCH and GLI shows that robust expression occurs in the gastric mesenchyme.39,40 Berman et al. 41 reported that PTCH-LacZ is expressed in gastric parietal cells. However, other studies have not followed their results. They also reported that resected human tumor samples overexpress PTCH, GLI, Shh, and Ihh. 41 Additionally, Fukaya et al. 42 detected the expression of the three Hh ligands (Shh, Ihh, and Dhh) and downstream signaling molecules of tissues in the human and mouse gastric epithelium by both immunohistochemistry and real-time polymerase chain reaction (RT-PCR) analyses (Table 1). Shh is a central factor in the differentiation of progenitor cells into various gastric cell origins rather than growth and for function in parietal cells. Specifically, lack of Shh increases the stem cell compartment at the expense of terminally differentiated cells.40,43
Expression of hedgehog-related molecules in the stomach and gastric cancers.
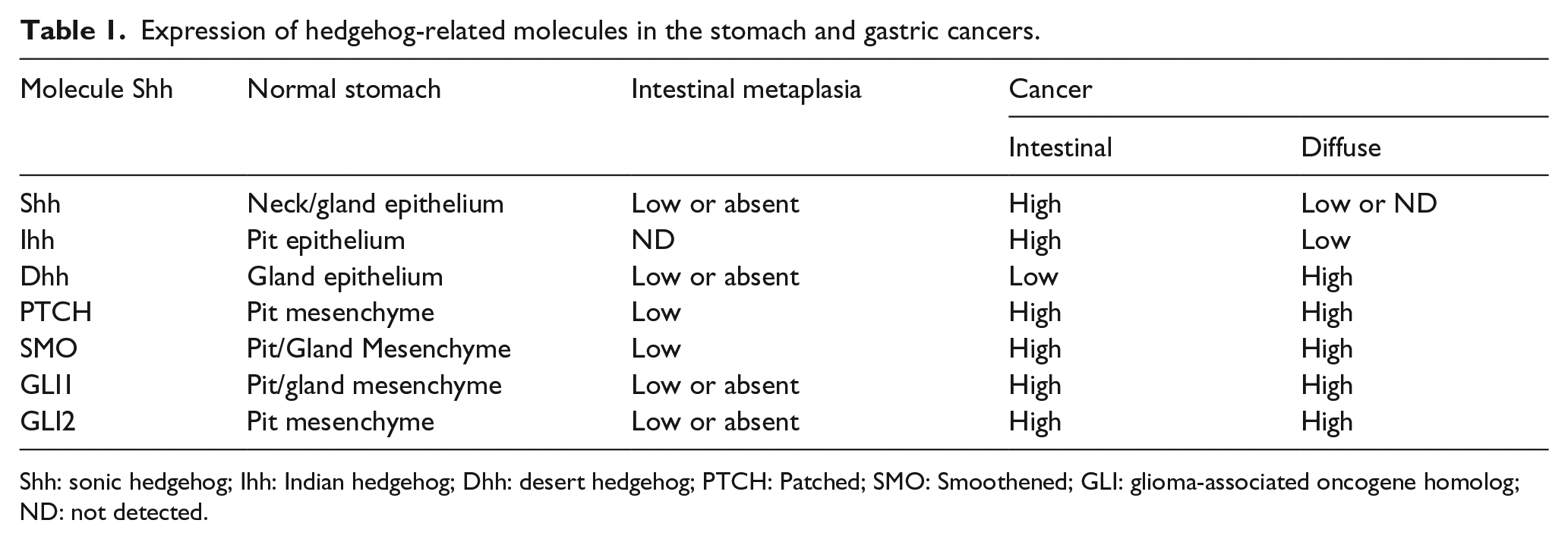
Shh: sonic hedgehog; Ihh: Indian hedgehog; Dhh: desert hedgehog; PTCH: Patched; SMO: Smoothened; GLI: glioma-associated oncogene homolog; ND: not detected.
Gastric cancers are distinguished into two main types: intestinal and diffuse. 44 The intestinal type is thought to trigger chronic inflammation, followed by parietal cell atrophy, intestinal metaplasia, dysplasia, gastric mucous cell hyperplasia, and cancer. 45 In contrast, the diffuse type is thought to cause deregulation of precursor cells or stem cells.46,47 Moreover, the familial form of diffuse cancer is caused by mutations in E-cadherin. Their cell origin is different, and the two types of gastric cancer show different Hh ligand expression (Table 1). The analysis of gastric cancer and metaplastic tissue samples showed that the three Hh ligands, PTCH, SMO, and the three GLI transcription factors were only weakly expressed or absent. In the intestinal type, Shh and Ihh expressions were detected. However, the expression of SMO, GLI 1, and 2 was unusual but very weak when detectable. 48 Shh, Ihh, and Dhh expression was diverse in diffuse gastric cancer types, but in contrast to the intestinal types, diffuse cancers expressed high levels of PTCH, SMO, and GLI (Table 1). Some studies have reported that Ihh expression was low in diffuse cancer cells, while Shh expression was not detected in epithelial cells nearby the cancer, but rather were expressed in adjacent fibroblast cells.42,49
Hh in GCSCs
The stochastic clonal evolution model has been proposed to explain gastric cancer growth. This model states that gastric cancer cell characteristics are altered by mutations, resulting in a population of frequently multiplexing cells. In contrast, the theory of CSC argues that only CSCs can promote tumor growth and self-renewal. 50 Because gastric carcinoma shows histological heterogeneity, CSCs may explain this multifunction observed in gastric cancer. 51 Studies of the origin of GCSCs are currently being conducted, and numerous recent studies have indicated that progenitor cells, gastric stem cells, or bone marrow–derived cells (BMDCs) are candidate GCSCs.52–54 Houghton et al. 52 demonstrated that BMDCs migrate and repopulate to contribute to dysplasia, metaplasia, and gastric carcinoma. Recent studies have provided compelling evidence that Helicobacter pylori infection induces an increase in BMDCs in gastric epithelial cells, which then participate in gastric cancer development. 55 Although most patients with advanced gastric cancer or metastatic gastric cancer require chemotherapy, the development of chemotherapy resistance is a major problem in the therapy of gastric cancer patients. Because the survival of CSCs is better than that of differentiated tumor cells or proliferating progenitor cells following administration of powerful anticancer therapies, 56 it may be essential to understand CSC chemo-resistance mechanisms before the development of promising anti-cancer reagents aimed at reducing chemo-resistance. Several signals (Wnt, Shh, Notch, and tumor necrosis factor beta (TNF-β)) are known to be related to the stemness of GCSCs and targeting of their signal pathways, which may play a pivotal role in CSC resistance and provide novel therapeutic modalities for advanced gastric cancer.57–60
The signaling pathway in GCSCs
The dysregulation of several major signal pathways may be related to gastric cancer and the self-renewal of GCSCs as stemness function regulators. 61 The Wnt/β-catenin, Notch, transforming growth factor beta (TGF-β), and Shh pathways in gastric cancer signaling are involved in cell proliferation, cell differentiation, and angiogenesis. Among these, Shh expression levels are the highest in the gastric unit. 62 The Shh signaling pathway is dysregulated in gastric cancer,41,63 and abnormal activation of the Shh pathway positively correlates with aggressive and poorly differentiated gastric cancer. 42 We previously reported that TGF-β-induced stimulation of the ALK5-Smad3 pathway plays an important role in gastric cancer metastasis through the Shh signaling pathway. 64 The Shh signaling pathway is critical for the maintenance of cancer stemness in advanced gastric cancer. 18 CD44+/CD24+ stem-like cells from a gastric cancer cell line showed upregulated messenger RNA (mRNA) expression of the Shh signaling molecules PTCH and GLI compared to the non-tumorigenic CD44−/CD24− cell population. 65 The Shh pathway may provide a therapeutic approach for targeting GCSCs for gastric cancer therapy.18,66 Cell surface markers for GCSC have been reported by many research groups19,22,23 (Figure 3). CD44, a transmembrane glycoprotein, is a downstream target of the Wnt/β-catenin signaling pathway. 67 CD44 is associated with cell signaling and migration and shows low expression in the gastric antrum. It has multiple isoforms, among which CD44H exhibits high affinity for hyaluronate and CD44 variants (CD44v) show metastatic properties. Chen et al. 68 suggested that the existence of CD44+ cells within gastric cancer tumors explains chemoresistance. In addition, Takaishi et al. 67 found that CD44+ cells reproduce morphologically and have a sphere-forming ability from the original gastric cancer tissues, demonstrating that CD44+ expression is a potential biomarker of GCSCs. Furthermore, Nishii et al. 21 reported that side population cells with high CD44 expression are likely to undergo metastasis on GCSCs. Han et al. 69 also reported that 500 epithelial cell adhesion molecule+/CD44+ cells from gastric cancer tissues have the potential to form xenograft tumors in mice. Other well-studied markers include CD 24, reported that the CD44+CD24+ subpopulation of gastric cancer cell line AGS is composed of GCSCs, 65 CD90, reported that a cell population with the CD90 phenotype enriched in sphere-forming cells from human gastric cancer tissues, 70 aldehyde dehydrogenase 1 (ALDH1), an enzyme in the ubiquitous ALDH family, identified as a marker of GCSCs by Katsuno et al., 71 and leucine-rich repeat–containing G protein–coupled receptor 5 (Lgr5). Lgr5+ cells are reported to coexist with CD44+, CD133+, and ALDH1+ cells not only in adjacent normal gastric mucosa but also in gastric cancer.72,73

Molecular properties of gastric cancer stem cells (GCSCs). GCSCs have the ability to self-renew and differentiate into diverse daughter cells, including progenitor cells and differentiated tumor cells. GCSCs show the characteristic biomarkers and stemness-maintaining signaling pathways. Moreover, GCSCs show crosstalk networks with stromal cells (myofibroblasts), which may secrete factors that regulate stemness and cancer cell differentiation in the tumor microenvironment.
Targeted agents inhibiting Hh signaling pathway
As described above, because Hh signaling is known to contribute to carcinogenesis and cancer progression, considerable efforts have been made to target the pathway. Various inhibitors of the Hh signaling pathway have been developed and evaluated in preclinical/clinical studies. Hh signaling is targeted at four major levels of the pathway: (1) inhibition of SMO, (2) disruption of receptor–ligand interaction, (3) blocking ligand processing, and (4) targeting GLI (Figure 4).

Molecular sites targeted by Hh signaling pathway inhibitors. These inhibitors target different components of Hh signaling, including Shh, Smo, and GLI1. These encompass natural compounds, their chemical derivatives, a monoclonal antibody, and chemicals identified from screening libraries.
SMO inhibitor
SMO is regarded as the most potential druggable target in this pathway. The plant-derived compound cyclopamine has been focused on as the first targeted agent to inhibit downstream signaling transduction by targeting of SMO. Cyclopamine exerts its action by binding directly to the heptahelical bundle of SMO and inhibits the activation of GLIs. 74 The results of previous studies evaluating the efficacy of cyclopamine revealed its considerable toxicity. Two previous studies using in vivo mice models of osteosarcoma and rhabdomyosarcoma showed no significant effect on the inhibition of tumor growth or metastasis.75,76 Warzecha et al. 76 reported skin toxicities such as scrubby coat and skin ulcerations in the mouse models, which prevented the authors from continuing the experiments to achieve a therapeutic dose of cyclopamine. Hirotus et al. reported cell cycle arrest of osteosarcoma in vitro by cyclopamine and inhibition of osteosarcoma growth as well as a survival benefit in vivo by knockdown of SMO. Difficulty in obtaining a therapeutic dose of cyclopamine because of the serious side effects was also reported in this study. 77 The weak potency of cyclopamine as well as other limitations such as poor solubility and acid sensitivity has impeded clinical development of the agent as a targeted agent for treating various cancers. Numerous efforts to overcome these limitations have been attempted, and in vitro screens have been performed, identifying a distinct chemical class of molecules. A small molecule with improved solubility and acid stability, vismodegib (GDC-0449), was identified through a high-throughput screen of small molecules 78 and was approved by the Food and Drug Administration for the treatment of recurrent, locally advanced, or metastatic basal cell carcinoma. Saridegib (IPI-926) is another novel semisynthetic cyclopamine analogue with better potency and metabolic stability compared with those of cylopamine. 79 Other developed SMO inhibitors with various chemical backbones include BMS-833923, LDE-225 (erismodegib), PF-0449913, LY2940680, and LEQ506.
Agents disrupting receptor–ligand interactions
5E1 (ch5E1) is a murine–human chimeric monoclonal antibody with anti-tumor activity that inhibits Hh signaling by binding to Shh. 80 Robotnikinin is another Hh signaling inhibitor discovered by small-molecule microarray-based screens. The ability of robotnikinin to inhibit the activation of Hh signaling by binding to the extracellular Shh protein was demonstrated in cell lines in a previous study. 81
Agents blocking ligand processing
A new agent with a different mechanism of action was reported by Petrova et al. A high-throughput screen revealed a small molecule inhibitor targeting Hh acyltransferase-mediated Shh palmitoylation, a hydrophobic modification process of native Shh. The authors also demonstrated that this small-molecule inhibitor inhibits Hh signaling in vitro.82,83
GLI inhibitors
GLI-mediated transcription is an attractive target in terms of inhibiting the activation of Hh signaling at the final step, enabling tumors to overcome resistance when the signaling pathway is activated downstream of SMO. A cellular screen identified two small-molecule inhibitors of GLI-mediated transcription, GANT 61 and GANT 58. Both GANT 61 and GANT 58 were revealed to act at the nuclear level. Although the precise mechanism of how these small-molecule antagonists inhibit GLI-mediated transcription has not been clarified, the authors suggested posttranslational modification of GLI1 by GANT 61 resulting in the prevention of DNA binding or destabilization of the GLI1-DNA complex. Moreover, inhibition of tumor cell growth was shown in an in vivo xenograft mice model of human prostate cancer cells. 84 Hyman et al. reported four Hh pathway inhibitors (HPIs) that impede the Hh signaling downstream of SMO with different mechanisms of action from GANT by performing high-throughput cellular screening, although the precise mechanisms of how each HPI inhibits Hh signaling remain to be clarified. HPI-1 is thought to inhibit signaling by targeting a posttranslational modification of GLI or interaction between GLI and co-factor. In addition, interference of GLI2 processing and trafficking was suggested as the mechanisms of HPI-2 and HPI-3 in the report. HPI-4 was thought to be the only agent that acts by perturbing ciliogenesis, although the mechanism by which HPI-4 perturbs ciliogenesis was not clarified. 85
Treatment of gastric cancer by targeting the Hh pathway
Only two clinical trials of agents targeting Hh signaling have been performed in patients with gastric cancer. A prospective, multicenter, randomized phase II clinical trial was performed to determine the efficacy of vismodegib, an SMO inhibitor, in combination with FOLFOX chemotherapy in patients with advanced gastric and gastroesophageal junction (GEJ) adenocarcinoma. A total of 124 patients with treatment-naïve, locally advanced, or metastatic gastric or GEJ adenocarcinoma were randomly assigned to receive FOLFOX plus vismodegib or FOLFOX plus placebo. The study failed to meet its primary endpoint, progression-free survival (PFS). The results showed that PFS was not significantly improved between the vismodegib group and placebo group (11.5 vs 9.3 months; p = 0.34). 86 Although no significant difference in PFS was observed between these two groups, a prolonged tendency in PFS was observed in the vismodegib group. A limited number of patients were analyzed in this study, which may explain the statistically unreliable results because the tendency for prolonged PFS was observed in patients who received vismodegib. Additionally, there was a lack of a biomarker for effective therapy with the targeted agent. A recent paper describing the role of Hh signaling in CD44 immunopositive gastric cancer cells reported the importance of CD44 expression, a marker of GCSCs, as a biomarker for sensitivity to the targeted agent. In the report, clinical gastric tumor samples from the phase II clinical trial were analyzed for the expression of CD44. The authors reported that tumor samples expressing high levels of CD44 from patients who received chemotherapy and vismodegib were related to enhanced survival. Additionally, high expression of CD44 in tumors from patients who received chemotherapy was found to be associated with poor survival, suggesting the potential role of CD44 as a biomarker in treatment with targeted agents against Hh signaling. 87 A phase 1 clinical trial to determine the maximum tolerated dose of BMS-833923 used in combination with cisplatin and capecitabine in patients with treatment-naïve, metastatic gastric, GEJ, or esophageal adenocarcinoma was completed in 2013, but the results have not yet been reported (NCT00909402).
Limitations to gastric cancer treatment
Although gastric cancer has been traditionally classified into two major types, differentiated type and undifferentiated type, based on its biologic behaviors and the appearance of the gastric cancer tissue, the biologic heterogeneity of gastric cancer is widely accepted. Tumor heterogeneity of gastric cancer cells has been regarded as one of causes for poor response to several targeted agents and obstacles in development of those agents in the treatment of gastric cancer. A previous study classified gastric cancer into four molecular subgroups: tumors positive for Epstein–Barr virus, microsatellite unstable tumors, genomically stable tumors, and tumors with chromosomal instability. 88 However, prospective or retrospective studies should be performed to validate the usefulness of this classification before application in the clinical field. One solution for achieving clinical benefits using targeted agents against the Hh signaling pathway in the treatment of patients with gastric cancer is the selection of a biomarker based on well-clarified mechanisms to identify patients expected to benefit from targeted agents. Another limitation of targeted agents in cancer treatment is resistance. Several small-molecule drugs have been specifically designed to target Hh signaling and are undergoing clinical trials. Unfortunately, apparent drug resistance was observed in these clinical trials, preventing clinical development. This drug resistance was considered to be associated with the overexpression of GLI2 and point mutations in SMO, which maintained constitutive activation of Hh signaling in solid tumors. Different effects of the targeted agents on different organs should also be considered. Shh signaling promotes epithelial cell proliferation in the stomach but inhibits proliferation in the intestine because of its negative feedback effects on the canonical Wnt/β-catenin signaling pathway. In order to overcome these limitations, well-designed clinical trials are essential to determine appropriate administration doses to avoid serious adverse effects, use of well-studied biomarkers in order to maximize the efficacy of the targeted agents, and combination therapy with chemotherapeutic agents or biologic agents to overcome resistance.
Conclusion
Abnormally activated Hh signaling is known to contribute to carcinogenesis by promoting the proliferation, invasion, and migration of tumor cells in a variety of cancers. In addition, the progression of cancers or resistance to treatment has been reported to contribute to the ability of activated Hh signaling to maintain and differentiate CSCs. Considering the various roles of the Hh signaling pathway in cancer progression of gastric cancer, targeting this signaling cascade is expected to result in the development of promising therapies. We reviewed the role of the Hh signaling pathway in gastric cancer, various agents targeting this pathway, and clinical trials performed in patients with gastric cancer. There are many limitations to introduce targeted agents in the clinical field, and no clinical benefit of these agents has been demonstrated. Because the selection of patients using appropriate biomarkers is important, this is also the case in the introduction of targeted agents for the treatment of patients with gastric cancer. For the limited therapeutic agents available for treating metastatic gastric cancer, the development of an effective new targeted agent is urgent. Optimal biomarkers based on pre-clinical data for treatment with agents targeting Hh signaling in patients with metastatic gastric cancer are expected to be introduced in future studies.
Footnotes
Acknowledgements
D.-H.L. and S.-y.L. equally contributed to this study as first authors.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Brain Korea (BK) 21 Plus Program and the grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI) and funded by the Ministry of Health and Welfare, Republic of Korea (grant number: HI15C1665).