
Abstract
Background
Buprofezin, a thiadiazine insecticide globally recognized as a molting inhibitor, is widely used for controlling whitefly and other insect pests.
Purpose
This study investigated the impacts of sub-lethal buprofezin concentrations (0.15, 0.30, and 0.60 mg/L) on adult Clarias gariepinus fish over an 8-day exposure period.
Materials and Methods
The 96-h LC50 was determined to be 2.4 mg/L. During the sub-lethal treatment, genotoxic analysis, hematological examination, biochemical analysis, and histopathological observations were performed.
Results
Genotoxic analysis revealed a gradual, concentration-dependent increase in micronucleus abnormalities, including binucleus, blebbed nuclei, lobed nuclei, and notched nuclei. Hematological examination revealed a considerable increase in white blood cells, neutrophils, monocytes, eosinophils, basophils, and mean corpuscular hemoglobin, alongside a reduction in lymphocytes, mean corpuscular hemoglobin concentration, red blood cell distribution width, and platelets. Biochemical analysis indicated a decline in alkaline phosphatase and total protein levels, while aspartate aminotransferase, alanine aminotransferase, and total glucose levels progressively increased with higher buprofezin doses. Histopathological observations revealed significant alterations in gills, such as cellular proliferation leading to lamellae fusion. The liver exhibited necrosis, vacuolization, hemorrhage, sac formation, enlargement, and rupturing of hepatocytes.
Conclusion
These findings collectively demonstrate that buprofezin induces significant genotoxic, hematological, biochemical, and tissue abnormalities in C. gariepinus, highlighting its potential ecotoxicological risks in freshwater ecosystems.
Introduction
The aquatic environment is currently under significant threat from the indiscriminate release of synthetic xenobiotics due to various human activities, posing substantial risks to non-target organisms. 1 While there is increasing research focus on environmentally safer pesticides, including plant-derived alternatives, synthetic pesticides remain crucial for maintaining agricultural yields by protecting crops from pests and vector-borne diseases, and for controlling weeds.2, 3 However, the growing reliance on pesticides, particularly in developing countries, often occurs without adequate consideration for the environmental consequences of their widespread and sometimes unethical application. 4
Insecticides, after their application, are dispersed into the environment through various pathways, frequently entering aquatic ecosystems via surface water runoff, precipitation, infiltration, and drainage from adjacent areas.4, 5 This contamination ultimately affects fish and other aquatic organisms, which are vital components of food webs. Beyond agricultural runoff, municipal and industrial discharges also contribute to water contamination, introducing highly toxic substances to non-target organisms. 4 Reports globally indicate an increased mortality among fish species in water bodies, often linked to insecticide and pesticide runoff from agricultural activities. The diverse concentrations of pesticides found in different types of wastewater are particularly harmful to aquatic organisms, especially fish, due to their inherent sensitivity to environmental pollutants. 6
Buprofezin (2-tert-butylimino-5-phenyl-3-propan-2-yl-1,3,5-thiadiazinan-4-one) is a thiadiazine pesticide, initially developed by Nihon-Nohyaku, Ltd. (Japan), which functions as a chitin synthesis inhibitor. 7 It is particularly effective against Homoptera pests, such as whiteflies, mealybugs, planthoppers, scales, and leafhoppers, by interfering with cuticle formation during molting in their incomplete growth stages. This action results in nymphal mortality and also suppresses adult oviposition, inhibiting chitin biosynthesis and reducing egg viability. 8 Despite its efficacy in pest control, buprofezin is widely used in paddy fields in various countries, including Malaysia, leading to its transport into water reservoirs, streams, and rivers, thereby causing severe effects on aquatic organisms. 8
The evaluation of micronucleus induction is a proven and effective assay for assessing the genotoxic effects of various compounds in fish. 9 Previous studies on African catfish embryos and larvae have shown that even low concentrations of buprofezin can cause malformations, decreased egg-hatching rates, and reduced larval survival.10, 11 Exposure to sub-lethal concentrations of buprofezin in Nile tilapia (Oreochromis niloticus) has been linked to oxidative stress and inflammatory responses. 9 Similarly, the synergistic effect of fipronil and buprofezin in common carp (Cyprinus carpio) resulted in changes in hematological, biochemical, and deoxyribonucleic acid (DNA) parameters, as well as histopathological alterations in gills, liver, and kidney. 12
Biochemical parameters, such as enzymatic activities (e.g., aspartate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP)) and metabolic indicators (e.g., glucose, total protein), are crucial biomarkers in ecotoxicology research.13–16 Changes in these parameters can signal physiological stress, cellular damage, and organ dysfunction in fish exposed to pollutants. For instance, elevated transaminases (AST, ALT) often indicate liver damage, while altered glucose levels reflect metabolic disruption due to stress. These biochemical alterations provide insights into the internal physiological state of the organism before overt histopathological changes become evident. 13
In Malaysia, catfish are extensively farmed, often in close proximity to agricultural areas, particularly paddy fields, which are characterized by continuous water supply and the regular, extensive use of pesticides, insecticides, herbicides, and fungicides. The African catfish (Clarias gariepinus) is a highly fecund species that is easily maintained under laboratory conditions and serves as a standard species for micronucleus tests, which analyze erythrocyte nuclear abnormalities for assessing genotoxicity in fish toxicology. 17 Teleost erythrocytes are nucleated, making them suitable for validating micronuclei (MN) and clastogenic activity. Furthermore, histopathology provides a functional tool for determining toxicant activity at the tissue level, offering useful information concerning organ damage. 18 Pollutants are known to cause histopathological changes in vital organs such as the gills, liver, and kidneys of fish. 18
Therefore, the current study was designed to comprehensively assess the impacts of buprofezin on freshwater adult African catfish (C. gariepinus). This research aims to fill a critical knowledge gap by being the first to evaluate the combined genotoxic, hematological, biochemical, and histopathological alterations induced by buprofezin in this commercially important and widely farmed species, particularly given its extensive use in Malaysian paddy fields. The findings will provide crucial insights into the ecotoxicological risks of buprofezin, informing environmental management practices and highlighting potential implications for aquatic food chains and human health.
Materials and Methods
Chemicals
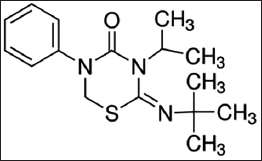
Buprofezin, with 98% purity, was purchased from Sigma–Aldrich International GmbH, Malaysia, and used for this study without further purification (Figure 1).
Chemical Structure of Buprofezin.37, 43
Experimental Animals
Live adult C. gariepinus of both sexes, with an average weight of 200–300 g and a total length between 28 and 33 cm (indicative of sexually mature adult fish), were obtained from a local market in Sungai Petani, Kedah, Malaysia. Fish were acclimatized for 7 days in large holding tanks (500 L) containing continuously aerated dechlorinated tap water. Water parameters during acclimatization were maintained as follows: temperature 26°C ± 2°C, pH 7.0 ± 0.5, dissolved oxygen 6.5 ± 0.5 mg/L. Fish were visually inspected twice daily for signs of stress, disease, injuries, or mortality. During acclimatization, fish were fed commercial fish pellets (40% crude protein) twice daily at 2% of their body weight. Experimental protocols and procedures used in this study were approved by the Human and Animal Ethics Committee, AIMST University, Kedah, Malaysia (AUHAEC/FAS/2017/01). All animal handling adhered to the Organization for Economic Co-operation and Development (OECD) guidelines for fish toxicity testing. 19
Acute Toxicity Evaluation
The median lethal concentration (LC50) of buprofezin for 24, 48, 72, and 96 h was determined following OECD Test Guideline 203. 19 A preliminary screening test was conducted to establish an appropriate range of toxic concentrations. The toxicant concentrations used in the acute study were 1.6, 1.8, 2.0, 2.2, 2.4, and 2.6 mg/L. For each concentration, 10 C. gariepinus fish were placed in separate cemented tanks containing 200 L of water with the respective buprofezin concentration. Feeding was suspended 24 h prior to and throughout the acute toxicity test to minimize interference with toxicant uptake and to prevent water quality degradation.
Chronic Toxicity Evaluation and LC50 Determination
Preliminary observations indicated 100% mortality at buprofezin doses exceeding 2.6 mg/L across all exposure durations (24, 48, 72, and 96 h). The 96-h LC50 value was calculated using probit analysis, 20 and estimated to be 2.4 mg/L of buprofezin. This LC50 value was subsequently used to derive the sub-lethal concentrations for the chronic exposure study. Sub-lethal studies are essential for assessing chronic toxicity under prolonged stress conditions caused by pesticides.
Experimental Design
For the chronic toxicity study, fish were randomly assigned to four experimental groups. Each group comprised 10 fish, and the experiment was performed in triplicate (n = 3), resulting in a total of 30 fish per treatment group across three tanks, and 30 fish for the control group. Each tank contained 200 L of continuously aerated dechlorinated tap water. The following experimental groups were maintained for a period of 8 days:
Group 1: Control (no buprofezin).
Group 2: 0.15 mg/L of buprofezin (1/16th of 96-h LC50).
Group 3: 0.30 mg/L of buprofezin (1/8th of 96-h LC50).
Group 4: 0.60 mg/L of buprofezin (1/4th of 96-h LC50).
The selected sub-lethal concentrations were chosen to represent environmentally relevant exposure levels that fish might encounter in aquatic ecosystems adjacent to agricultural areas in Malaysia, where buprofezin is commonly applied. A stock solution of buprofezin was prepared by dissolving the pure compound directly in dechlorinated tap water. Desired sub-lethal concentrations were then prepared by serial dilution from the stock solution. No organic solvents were used; therefore, a solvent control was not deemed necessary. Feeding was withheld during the entire chronic exposure period to minimize metabolic interference and maintain water quality consistency.
Genotoxic Analysis
Blood samples were collected from the posterior caudal vein of six fish per group (two fish per replicate tank) using a 3 mL syringe at the end of the 1st, 4th, and 8th days of exposure, following OECD Test Guideline 210. 21 One drop of blood was placed near one end of a clean glass slide. Another slide was used to spread the blood drop evenly, creating a thin smear that covered approximately half of the slide. 22 The blood smears were fixed using 100% ethanol for 20 min and subsequently stained with a 10% Giemsa solution for 15 min. Slides were then rinsed with distilled water, air-dried, and examined under a light microscope. MN were identified as circular or ovoid chromatin bodies with the same staining intensity as the main nucleus, but separate from it. Cells containing two nuclei were categorized as binuclei (BN). Blebbed nuclei (BL) were characterized by evagination of the nuclear membrane with the presence of euchromatin. Lobed nuclei (LB) were larger than BL and displayed multiple lobes. Nuclei with vacuoles and those lacking clear nuclear material were identified as notched nuclei (NT). 23 For each slide, 1,000 erythrocytes were examined to determine the frequency of MN and other nuclear abnormalities.
Hematological Analysis
Blood samples were collected from the posterior caudal vein of six fish per group (two fish per replicate tank) using a 3 mL syringe at the end of the 1st, 4th, and 8th days of exposure. Samples were immediately transferred into tubes containing ethylenediaminetetraacetic acid (EDTA) as an anti-coagulant. The following hematological parameters were analyzed using a CELL-DYN 2 3500 1 unit cell dye CD-3500SL analyzer S/NO 31189 P: white blood cell (WBC) count, lymphocyte (LYM) percentage, neutrophil (NUE) count, eosinophil (EOS) count, monocyte (MONO) count, basophil (BASO) count, mean corpuscular hemoglobin concentration (MCHC), mean corpuscular hemoglobin (MCH), mean red blood cell distribution width (RDW), and platelets (PLT).
Biochemical Analysis
At the end of the 1st, 4th, and 8th day of exposure, blood samples were collected from the posterior caudal vein of six fish per group (two fish per replicate tank) and immediately centrifuged at 4,000 rpm for 5 min using a JUAN B41 benchtop centrifuge multi-functional P/N 11175600 to separate the serum. The isolated serum was then used to analyze metabolites (glucose and total protein) and enzymes such as ALP, AST, and ALT using an Abbot Architect CI8200 analyzer. These blood biochemical parameters served as systemic indicators of physiological stress and organ dysfunction, reflecting changes in liver, kidney, and gill function due to their circulating nature and their critical roles in metabolism and detoxification.
Histopathological Analysis
At the end of the 4th and 8th days of exposure, samples of gills, liver, and kidney were collected from three fish per treatment group (one fish per replicate tank) and immediately fixed in 10% neutral buffered formalin. Histopathological changes typically develop over a longer duration compared to acute genotoxic, hematological, and biochemical alterations. Therefore, to observe significant and progressive tissue damage, samples for histopathological analysis were collected on days 4 and 8, focusing on the later stages where more pronounced effects would be evident. To identify the amount of cellular damage caused by buprofezin, gill, liver, and kidney tissues from control and treated fish were subsequently processed. Tissues were dehydrated through a graded ethanol series, cleared in xylene, and embedded in paraffin wax. Sections were cut into 3 µm thickness using a microtome and stained with hematoxylin and eosin (H&E). 24 Stained sections were then examined under a light microscope to identify and document cellular and tissue alterations.
Statistical Analysis
All experiments were performed in triplicate (n = 3), and results were expressed as the mean ± standard deviation (SD). Statistical analysis was carried out using the Statistical Package for the Social Sciences (SPSS) package version 22. Data were subjected to one-way analysis of variance (ANOVA) followed by Duncan’s Multiple Range Test (DMRT) to determine significant differences between groups. A significance level of p < .05 was used for all statistical comparisons.
Results
Preliminary Observations and LC50 Determination
Preliminary screening tests indicated that buprofezin doses beyond 2.6 mg/L resulted in 100% mortality of all test fish within 24, 48, 72, and 96 h. Based on these observations and probit analysis, 20 the 96-h LC50 value for buprofezin in C. gariepinus was estimated to be 2.4 mg/L. This value served as the basis for selecting sub-lethal concentrations for the chronic toxicity study, which are crucial for assessing prolonged stress conditions caused by pesticides.
Micronucleus and Erythrocyte Morphology
Exposure to buprofezin at concentrations of 0.15, 0.30, and 0.60 mg/L induced distinct structural changes and nuclear abnormalities in the red blood cells (RBCs) of C. gariepinus (Table 1). The frequency of MN and other nuclear abnormalities (BN, BL, LB, and NT) significantly increased with higher buprofezin concentrations (0.60 mg/L, showing the most pronounced effects) and longer exposure durations (day 4 and 8, compared to day 1). By day 4 at concentrations of 0.30 and 0.60 mg/L, early morphological changes were observed, including the formation of echinocytes (crenated RBCs with spiky or irregular surfaces) and frequent acanthocytes. In addition, teardrop-shaped RBCs with pointed apices were also noted.
Micronucleus Profiles of Clarias gariepinus Exposed to Sub-lethal Toxicity Levels of Buprofezin.
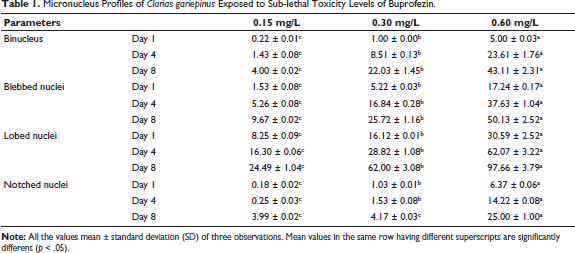
By day 8, at 0.15 and 0.60 mg/L, morphological changes such as sickle cells (SK) of various shapes (ellipsoidal, bean-shaped, and true sickles), swelled cells (SC) with water-filled cytoplasm, and one or more MN per cell were observed. Exposure to 0.30 mg/L on day 8 showed discrete MN, SK, joined RBCs, and SC. Fish treated with 0.60 mg/L on day 8 exhibited severely altered cells with widespread degeneration of RBCs. At 0.15 mg/L on day 8, common RBC malformations included altered, joined, and irregular shapes, along with pale cytoplasm. The most frequent and severe changes were noticed in fish treated with 0.30 and 0.60 mg/L on day 4, where cells joined together in chain-like formations, showing signs of karyolysis (dissolution of the cell nucleus). Cell nuclei with loss of their central dye, RBCs with visible vacuoles, and numerous SK were prevalent in fish treated with 0.30 and 0.60 mg/L on day 8 (Figure 2).

Hematological Parameters
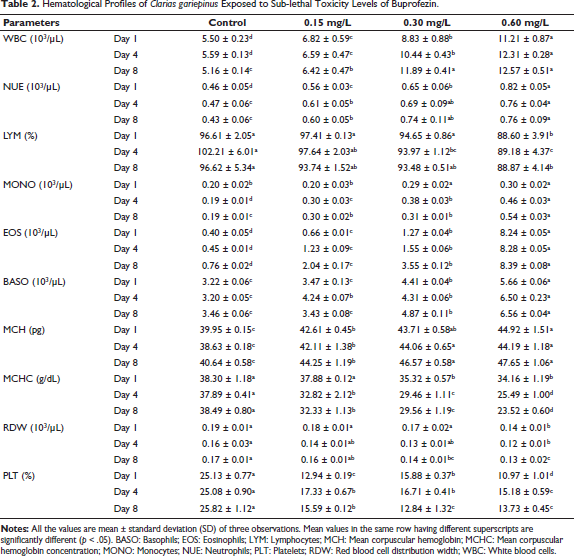
The hematological profile of C. gariepinus demonstrated significant alterations in response to buprofezin exposure (Table 2). A gradual increase was observed in the levels of WBC, NUE, MONO, EOS, BASO, and MCH, progressing from the lower concentration (0.15 mg/L) to the higher concentration (0.60 mg/L) across the exposure period. Conversely, the levels of LYM, MCHC, mean RDW, and PLT gradually decreased with increasing concentrations of buprofezin and longer exposure durations. These changes indicate a dose- and time-dependent impact on the fish’s immune response and erythropoietic system.
Hematological Profiles of Clarias gariepinus Exposed to Sub-lethal Toxicity Levels of Buprofezin.
Biochemical Parameters
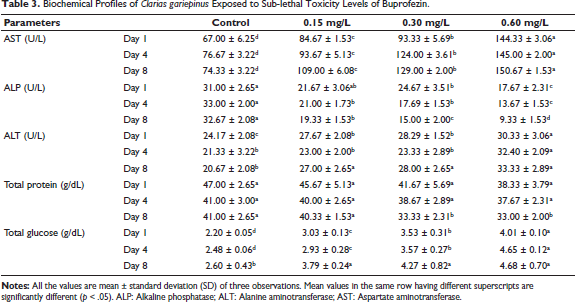
Exposure to buprofezin induced significant dose- and time-dependent changes in the biochemical parameters of C. gariepinus (Table 3). The levels of AST, ALT, and total glucose increased progressively from the lower concentration (0.15 mg/L) to the higher concentration (0.60 mg/L) of buprofezin across the exposure period. In contrast, the levels of ALP and total protein decreased as the concentration of buprofezin increased and with prolonged exposure. These alterations suggest metabolic disruption and potential organ damage.
Biochemical Profiles of Clarias gariepinus Exposed to Sub-lethal Toxicity Levels of Buprofezin.
Histopathology of Gill
Normal Structure in Control Fish
Sections of the gill from control fish exhibited a normal and regular cellular pattern, characterized by intact gill rakers, a healthy gill arch, well-structured primary and secondary filaments, clear sinuses, venules, pseudo-brachial lamellae, robust cartilaginous support, the ceratobranchial bone of the arch, distinct nuclei, and a normal mucous epithelium lining the membrane, along with clearly defined branches of the afferent and efferent arterioles (Figure 3A).
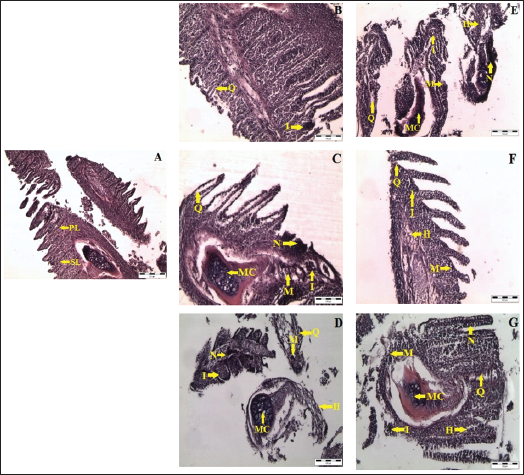
Alterations Due to Buprofezin Exposure
Exposure to buprofezin at various concentrations and durations resulted in significant histopathological changes in the gill tissues. Observed alterations included a marked reduction in the length or complete loss of the primary and secondary lamellae, often accompanied by clavate formation (club-shaped lamellae). Within the respiratory epithelium, widespread necrosis and hyperplastic accumulation of cells, alongside separated epithelial cells, were prominent in the clavate secondary lamellae. Ballooning dilation, irregular capillary circulation, and the presence of free erythrocytes within the secondary lamellae were particularly noted at 0.30 and 0.60 mg/L on both day 4 and day 8 (Figure 3C, 3D, 3F, 3G). Accumulation of hemocytes was evident beneath the tips of the primary lamellae and within the epithelium of secondary lamellae. Furthermore, cystic structures were observed in the secondary lamellar epithelia. In the primary lamellae, proliferation and thickening of the cartilage-perichondrium, along with deformation and alteration in the cartilage cells, were present, especially at 0.60 mg/L on day 4, and at 0.15 and 0.30 mg/L on day 8 (Figure 3D–3F).
Histopathology of the Kidney
Normal Structure in Control Fish
Kidney sections from control fish displayed normal architecture, including well-defined renal corpuscles with clear, healthy glomeruli, and intact proximal and distal convoluted tubules lined by healthy epithelial cells, demonstrating normal renal function (Figure 4A).
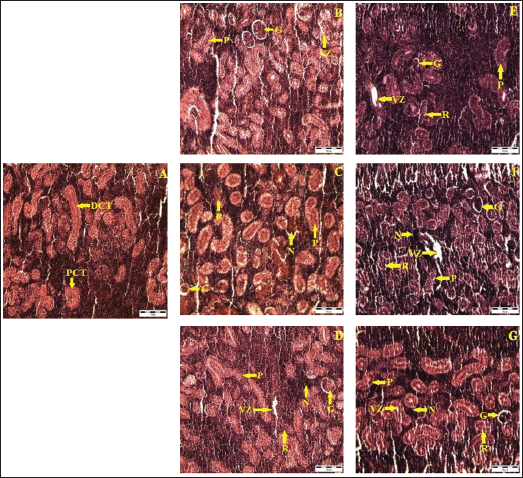
Alterations Due to Buprofezin Exposure
Exposure to buprofezin led to significant structural damage in the renal tissue. A noticeable decrease in renal cell numbers was observed in both proximal and distal collecting tubules. The tubular cells exhibited hypertrophy, and several renal tubules lost their original shape. Severe vacuolation, resulting from the degeneration of the cytoplasm, was frequently seen. Hyperplasia, degeneration, and necrosis, which could progress to complete tissue necrosis, were prominent, particularly at 0.60 mg/L on day 4 and consistently across all treated groups (0.15, 0.30, and 0.60 mg/L) on day 8 (Figure 4D–4G). The cuboidal epithelial cells lining the tubules showed complete vacuolation with cytoplasmic degeneration, and further nuclear division with improper structure was evident in fish exposed to 0.15, 0.30, and 0.60 mg/L on day 8.
Histopathology of Liver
Normal Structure in Control Fish
A cross-section of the liver from control fish demonstrated a normal central vein, regular cellular patterns of hepatic plates, healthy hepatocytes, well-defined spaces of Disse, and an intact biliary epithelium, indicating normal liver architecture and function (Figure 5A).
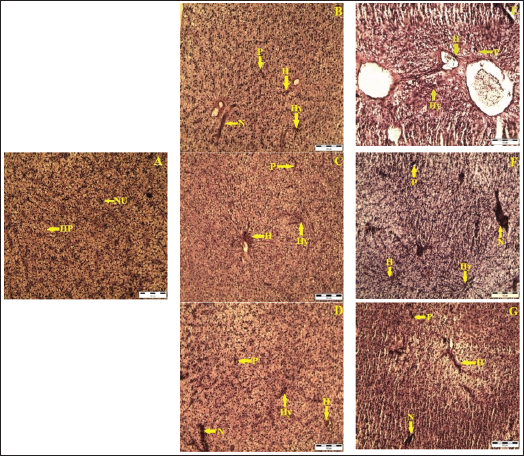
Alterations Due to Buprofezin Exposure
Exposure to buprofezin induced significant histopathological abnormalities in the liver. Lesions, signs of malignancy, widespread necrosis, accumulation of pigments, inclusion bodies, and inflammation were observed, particularly at 0.30 and 0.60 mg/L on day 4 (Figure 5C, 5D). Sections from fish treated with 0.30 mg/L on day 4 showed slight lesions, inclusion bodies, inflammation, necrosis, malignancy, and pigments. Additionally, hypertrophy, further necrosis, and vacuolation were evident. Empty spaces resulting from marked hypertrophy, necrotic hepatocytes, and pyknotic cells (cells with condensed nuclei) were clearly visible. Severe hypertrophy, hemorrhage, extensive necrotic hepatocytes, pyknosis, and generalized vacuolation were increasingly observed across all treated groups (0.15, 0.30, and 0.60 mg/L) on day 8 (Figure 5E–5G).
Discussion
The aquatic environment faces increasing threats from anthropogenic xenobiotics, with pesticides posing a significant risk to non-target aquatic organisms like fish. 1 This study provides a comprehensive assessment of the genotoxic, hematological, biochemical, and histopathological impacts of buprofezin, a widely used thiadiazine insecticide, on adult freshwater African catfish (C. gariepinus). While previous studies have examined aspects of buprofezin toxicity in fish larvae or other species,9–12 this research uniquely integrates a range of biomarkers in adult C. gariepinus, a commercially important species often farmed near agricultural areas with high pesticide usage in Malaysia. Our findings reveal a clear dose- and time-dependent toxicological profile, indicating that buprofezin induces significant physiological and cellular damage.
Genotoxic Effects
The micronucleus test is a well-established and effective assay for evaluating genotoxic effects in fish, providing insights into chromosomal damage and nuclear abnormalities.23, 25 Our results demonstrate a significant increase in MN and other nuclear abnormalities, such as BN, BL, LB, and NT, in the erythrocytes of C. gariepinus exposed to buprofezin. This increase was directly correlated with both higher concentrations and longer exposure durations. The frequency of LB, a common indicator of genotoxic damage, showed a substantial rise under these conditions, thereby indicating significant genetic damage accumulation (Table 1). These nuclear alterations suggest clastogenic and aneugenic effects, potentially arising from the failure of normal cell division, damage to erythrocyte chromosomes, or interference with DNA replication.23, 26 The presence of MN specifically points to chromosomal breakage or spindle apparatus dysfunction, leading to the exclusion of chromosome fragments or whole chromosomes from the main nucleus during cell division. Other abnormalities, like blebbed and LB, while not direct MN, serve as complementary indicators of genetic instability and cellular distress caused by the toxicant.23, 26 The observed increase in these abnormalities over time strongly indicates an accumulation of genotoxic damage rather than adaptation, suggesting a persistent assault on the cellular genetic machinery. This aligns with findings in Barbonymus gonionotus exposed to quinalphos 25 and C. carpio exposed to fipronil and buprofezin, where genetic damage was pronounced. 12
Hematological Alterations
Hematological parameters are sensitive and reliable indicators of physiological stress and immune status in fish exposed to environmental pollutants. In the present study, buprofezin exposure induced significant alterations in the hematological profile of C. gariepinus, with a clear dose- and time-dependent pattern (Table 2). We observed a gradual increase in WBC count, NUE, MONO, EOS, BASO, and MCH, from lower to higher buprofezin concentrations. The elevated WBC count, particularly the increase in NUE, MONO, and EOS, suggests an immunological response to the stress and potential infection induced by the pesticide.27–30 This is likely a protective mechanism, where the fish’s immune system attempts to cope with the xenobiotic stress by increasing circulating leukocytes. However, a sustained increase can also indicate chronic inflammation or an overwhelmed immune system.29–33 Conversely, LYM count, MCHC, RDW, and PLT gradually decreased with increasing buprofezin concentrations. A decrease in LYM often suggests an immunosuppressive effect or a compensatory response due to damage to circulating LYM. 31 The reduction in MCHC is indicative of variations in erythrocyte size and hemoglobin content, often signaling microcytic anemia, 34 while decreased RDW can reflect altered erythropoiesis. A significant, dose-dependent decrease in platelet count was observed, suggesting impaired thrombopoiesis or increased platelet destruction. 28 This finding is consistent with other studies where toxic substances caused anemia and affected platelet production in aquatic animals.28, 31 The overall hematological picture points towards a compromised immune system and developing anemic conditions, making the fish more susceptible to diseases and further environmental stressors. The changes generally became more pronounced with increased exposure duration, further suggesting cumulative toxic effects rather than adaptation.
Biochemical Changes
Biochemical parameters, including enzyme activities and metabolite levels, serve as crucial biomarkers for assessing organ dysfunction and metabolic disturbances in fish exposed to toxicants. 1 In C. gariepinus, exposure to buprofezin resulted in significant dose- and time-dependent alterations in key biochemical indicators (Table 3). AST and ALT levels showed a progressive increase with higher buprofezin concentrations and longer exposure durations. For instance, AST showed a substantial increase from control levels to those observed at the highest concentration and longest exposure. Similarly, ALT demonstrated an increase under the same conditions. These enzymes are commonly found in the liver, kidney, gills, and muscle, and their elevated plasma levels typically indicate organ dysfunction or tissue injury, particularly hepatocellular damage.13, 31, 35 This suggests that buprofezin is causing cellular leakage and damage to vital organs involved in metabolism and detoxification. Conversely, ALP activity and total protein levels decreased as buprofezin concentrations increased. ALP decreased significantly, particularly at the highest concentration and longest exposure duration. This reduction in ALP suggests a disruption in metabolic pathways, potentially reducing substrate availability in cycles like the Krebs’ cycle. 13 Total protein levels also declined, particularly on day 8 at higher concentrations. Decreased protein content often indicates an adaptive response to stress, where proteins may undergo proteolysis to meet energy demands, or there might be impaired protein synthesis due to liver damage.36, 37 This suggests an energy crisis and altered protein metabolism in C. gariepinus under buprofezin stress. Conversely, total glucose levels showed a gradual increase, correlating with both rising buprofezin concentrations and extended exposure durations. This hyperglycemia is a classic stress response in fish, where cortisol and catecholamines stimulate glycogenolysis and gluconeogenesis to provide immediate energy for coping mechanisms. Such an increase in plasma glucose levels indicates significant physiological stress prompted by buprofezin. The continuous elevation suggests a sustained stress response rather than a return to homeostasis, reinforcing the chronic toxic effects.
Histopathological Alterations
Histopathological examination is a powerful tool for directly assessing tissue-level damage caused by toxicants and provides crucial information about the affected organs. 35 Our study revealed significant histopathological changes in the gills, kidney, and liver of C. gariepinus, reflecting the cumulative impact of buprofezin exposure.
Gills
The gills are primary organs of gaseous exchange and osmoregulation, making them highly susceptible to waterborne pollutants. 14 Buprofezin exposure induced severe alterations in gill architecture. We observed excessive mucus secretion, lamellar fusion, reduction, and swelling in the epithelial cells of secondary lamellae, as well as hyperplasia and edema, particularly at higher concentrations and longer durations (days 4 and 8) (Figure 3). These changes are consistent with typical responses to irritants, where excess mucus acts as a protective barrier but simultaneously impairs oxygen uptake. Lamellar fusion reduces the effective surface area for gas exchange, leading to respiratory distress.9, 38 Hyperplasia of epithelial cells further thickens the diffusion barrier, exacerbating respiratory problems. These findings are similar to those reported for Mystus vittatus exposed to phenol 38 and other fish species exposed to various pesticides. 9 The increasing severity of these lesions from day 4 to day 8 indicates progressive damage.
Kidney
The kidney plays a vital role in excretion and osmoregulation, making it vulnerable to toxicants. 39 C. gariepinus exposed to buprofezin exhibited significant structural damage in the renal tissue, including inflammation of tubules, tubular necrosis, vacuolation, shrinkage, and damage to the glomeruli (Figure 4). Hypertrophy of tubular cells and loss of the original tubule shape were also evident. Furthermore, desquamation of the tubular epithelium and dilation of the lamina were observed. These findings align with other studies showing histological damage and functional interference in fish kidneys due to various pesticides.36, 40 The degenerative changes, especially vacuolation and necrosis, indicate direct cytotoxicity to renal cells and impaired excretory function. The progression of these lesions over the 8-day period suggests that the kidney’s capacity to detoxify or excrete buprofezin metabolites may be overwhelmed.
Liver
The liver is the primary organ for metabolism and detoxification of xenobiotics, rendering it highly susceptible to chemical-induced damage.41, 42 Our histopathological analysis revealed prominent abnormalities in the liver of buprofezin-exposed C. gariepinus. These included lesions, malignancy, necrosis, pigments, inclusion bodies, inflammation, enlargement of hepatocytes, rupture of hepatocytes and their nuclei, space formation, and aggregation of nuclei (Figure 5). These severe changes, particularly necrosis and hypertrophy, indicate significant hepatocellular damage and dysfunction. Necrosis reflects irreversible cell death, while hypertrophy can be an initial adaptive response to increase metabolic capacity, but in this context, it appears linked to subsequent degeneration. The presence of vacuolation suggests lipid accumulation or mitochondrial swelling, indicative of metabolic disruption. These findings are consistent with observations in C. carpio treated with copper nanoparticles, and Nile tilapia exposed to pendimethalin, 21 where similar hepatic lesions were reported due to toxicant exposure. The severity of liver damage, increasing with both concentration and duration, underscores the liver’s role as a major target organ for buprofezin toxicity. The sudden withdrawal and utilization of glycogen from liver cells to fulfill energy requirements during stress conditions could also contribute to the observed structural changes. 2 The integrated analysis of genotoxic, hematological, biochemical, and histopathological biomarkers demonstrates a clear cascade of toxic effects. Genotoxic damage at the cellular level likely contributes to overall physiological stress, reflected in hematological and biochemical imbalances. These systemic disturbances, in turn, manifest as observable structural damage in vital organs like the gills, kidneys, and liver. Importantly, in all observed parameters, the toxicological effects were more pronounced with increasing buprofezin concentration and, critically, with longer exposure durations. This suggests that C. gariepinus did not adapt to the chronic sub-lethal exposure; instead, the cumulative damage intensified over the 8-day study period, indicating a progressive rather than transient toxic response.
Conclusion
This study demonstrates that buprofezin exposure induces significant adverse effects on adult African catfish (C. gariepinus). We observed erythrocyte micronucleus abnormalities, indicating genotoxic stress, along with considerable alterations in hematological and biochemical parameters that signify systemic physiological disruption and compromised immune function. Furthermore, histopathological examination revealed severe damage to vital organs, including gills, kidneys, and liver, which are critical for respiration, excretion, and detoxification, respectively. These findings collectively suggest that buprofezin, even at sub-lethal concentrations, poses a substantial threat to the health of aquatic organisms. Given that C. gariepinus is a commercially important species often farmed near agricultural areas where buprofezin is widely used, these results have important implications for environmental management. Therefore, suitable controlled and regular use of insecticides, coupled with continuous monitoring, is highly recommended to acquire the beneficial effects of pest control without polluting the environment and without leaving harmful residues in food and water sources, thereby protecting aquatic life and human health.
Footnotes
Acknowledgments
The authors would like to acknowledge the administrative and technical support provided by the Faculty of Applied Sciences, AIMST University, and the School of Biological Sciences, Universiti Sains Malaysia, Penang, during this study.
Abbreviations
Authors Contribution
Suresh V. Chinni: Conceptualization, validation, visualization, project administration. Kasi Marimuthu: Conceptualization, validation, software, data curation, writing—review & editing, supervision. Gobinath Ramachawolran: Methodology, formal analysis, investigation, writing—original draft preparation. Sreeraman Subramaniam: Validation, resources. All authors have read and agreed to the published version of the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The experimental protocols and procedures utilized in this study received approval from the Human and Animal Ethics Committee, AIMST University, Kedah, Malaysia, under reference number AUHAEC/FAS/2017/01. All animal handling and experimental practices adhered strictly to the guidelines established by the OECD for fish toxicity testing.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Publication fee was supported by RCSI & UCD Malaysia Campus (RUMC).
Informed Consent
Not applicable.