
Abstract
Background
Cerebral ischemic stroke, a devastating neurological condition, arises from the disruption of blood flow to the brain, resulting in oxygen and nutrient deprivation and subsequent neuronal damage. Ischemic stroke is responsible for a significant number of all stroke incidences, highlighting the urgency for effective treatments.
Purpose
The present study aimed to investigate the therapeutic efficacy of gigantol in rats with cerebral ischemia–reperfusion injury (CI–RI).
Materials and Methods
The CI–RI was induced in experimental rats, and gigantol was administered intragastrically at 10 and 20 mg/kg concentrations for 7 days prior to CI–RI induction and for an additional 14 days. The levels of brain edema and infarction in the experimental rats were assessed. The concentrations of antioxidants, oxidative stress markers, inflammatory cytokines, and apoptotic proteins were assessed using the kits.
Results
The current findings demonstrated that gigantol treatment significantly reduced brain edema and infarction levels in CI–RI-induced rats. The gigantol treatment considerably reduced pro-inflammatory cytokine levels and alleviated oxidative stress by enhancing antioxidant concentrations in the brains of CI–RI rats. The gigantol treatment also regulated the levels of apoptotic proteins in the rats with CI–RI.
Conclusion
The current study revealed the neuroprotective effects of gigantol in rats subjected to CI–RI due to its antioxidant, anti-inflammatory, and anti-apoptotic properties. These findings may promote the advancement of gigantol as a potential salutary candidate to treat brain damage due to ischemic stroke.
Introduction
Cerebral ischemic stroke, a prevalent cerebrovascular disease, poses a significant threat to global health due to its elevated morbidity and mortality rates. A stroke, in general, is defined as an abrupt neurological deficit resulting from impaired blood flow to the brain. Ischemic stroke, specifically, develops from a blockage in a cerebral blood vessel, leading to a decrease or blockage of blood flow to the brain, and it constitutes approximately 85% of all stroke cases (Li et al., 2024). This deprivation of oxygen and nutrients initiates several cellular events ending in infarction, or cell necrosis, within the affected area. The complications of ischemic stroke are wide-ranging, impacting motor function, sensory perception, cognitive abilities, and emotional well-being. Prompt diagnosis and intervention are crucial for minimizing brain damage and improving patient outcomes (Donkor, 2018). The burden of ischemic stroke is substantial, affecting millions globally and placing a heavy strain on healthcare systems. Stroke is a foremost cause of permanent disability, with up to 50% of survivors experiencing motor, cognitive, or speech impairments. The economic impact of stroke is also significant, encompassing healthcare costs, rehabilitation expenses, and lost productivity (Ma et al., 2021). Globally, the burden is also higher in low- and middle-income nations, where preventive methods, acute treatment, and rehabilitation are often restricted. The aging global population further exacerbates the burden of stroke, as the cases of stroke increase with age. Projections estimate a substantial rise in stroke-related deaths and disabilities in the coming decades, underscoring the need for potential protective strategies and efficient stroke care worldwide (Pandian et al., 2018).
The causes of ischemic stroke are diverse, but they primarily revolve around factors that compromise cerebral blood flow. Atherosclerosis, the buildup of plaque within arterial walls, is a major contributor. These plaques can narrow the arteries, impeding blood flow and potentially rupturing to form thrombi that occlude vessels. Lifestyle factors like smoking, obesity, less physical activity, and unhealthy diets also contribute to the high risk of stroke. Addressing these modifiable risk factors through lifestyle changes and medical management is crucial for stroke prevention (Bushnell et al., 2014). The pathogenesis of stroke comprises a complex interplay of cellular and molecular events that unfold after the blockage of blood flow. The initial event is a reduction in nutrients and oxygen delivery to the affected brain tissue, resulting in energy failure and disruption of cellular function. Within seconds of ischemia onset, neurons become depolarized, releasing the excitatory neurotransmitter glutamate into the extracellular space (Su et al., 2022). Excessive glutamate accumulation triggers excitotoxicity, overstimulating receptors on neighboring neurons and leading to cellular damage. The ischemic cascade progresses through the formation of free radicals, inflammation, and the activation of apoptotic pathways, ultimately leading to neuronal apoptosis. Microglia are rapidly activated after ischemia, contributing to neuroinflammation (Khatri et al., 2012). Reperfusion, while essential for saving the penumbra, can paradoxically cause additional damage, known as reperfusion injury. This phenomenon involves oxidative stress, inflammation, and disruption of the blood–brain barrier (BBB). Targeting oxidative stress presents a rational therapeutic approach, given its role in the complexities of oxygen deprivation (Sun et al., 2018). Moreover, inflammation, triggered by ischemia, exacerbates neuronal damage following strokes. The complex pathogenesis of stroke underscores the importance of a multifaceted approach to treatment, targeting various points in the ischemic cascade to minimize brain damage and improve outcomes (Wu et al., 2020).
Current clinical interventions primarily focus on restoring blood flow to the affected brain region through thrombolysis and mechanical thrombectomy. Beyond recanalization strategies, neuroprotective approaches aim to mitigate the cascade of cellular and molecular mechanisms that result in neuronal death in the ischemic penumbra. Despite the advancements in acute stroke therapy, a considerable number of patients still suffer from long-term disabilities, emphasizing the need for more effective and safe alternative therapies (Kaesmacher et al., 2024). Gigantol is a bibenzyl compound extracted from many species of orchids belonging to the genus Dendrobium, such as Dendrobium aurantiacum. Gigantol exhibits several pharmacological properties, including anti-inflammatory (Won et al., 2006), anti-cancer (Yu et al., 2018), anti-cataractogenic (Fang et al., 2015), anti-colitic (Yu et al., 2024), and hepatoprotective (Xue et al., 2020) effects. Nonetheless, the neuroprotective impacts of gigantol in cerebral ischemia–reperfusion injury (CI–RI) model have not been previously investigated. Therefore, this study was planned to assess the therapeutic efficacy of gigantol in CI–RI-induced rats.
Materials and Methods
Experimental Rats
Adult male Sprague-Dawley rats were employed in the present study. The rats were caged in sterilized polypropylene cages under regulated laboratory environments, maintaining a temperature of 22°C–26°C and humidity of 50%–60%. A 12-h cycle of alternating illumination and darkness was established. During the entire study period, all rats were provided with free access to a conventional rodent meal and purified drinking water. Before the commencement of the research, all rats had a 7-day acclimation phase in a laboratory environment.
Middle Cerebral Artery Occlusion-induced Cerebral Ischemia–Reperfusion Injury Model
The middle cerebral artery occlusion (MCAO) procedure was performed on the experimental rats as previously outlined (Longa et al., 1989). All rats were sedated with 10% chloral hydrate (i.p.), after which an intraluminal suture was introduced from the external carotid artery into the internal carotid artery. After 2 h, the suture was removed, and blood flow was restored. The sham-operated group received an identical operation without arterial ligation. Rats were arbitrarily distributed to four groups: Group I: Sham rats; Group II: MCAO-induced CI–RI rats; Group III: MCAO-induced CI–RI + gigantol (10 mg/kg)-treated rats; Group IV: MCAO-induced CI–RI + gigantol (20 mg/kg)-treated rats. Gigantol was given intragastrically for 7 days before the MCAO operation and for 14 days, commencing 4 h post-reperfusion, until the animals were euthanized. A comparable amount of 10% Tween-80 was administered in both the sham and MCAO groups.
Assessment of Brain Edema
The brain edema was assessed using the wet/dry weight technique. Following the scarification of the animals, brains were excised and weighed to obtain wet weight. After a 72-h desiccation phase at 70°C, the brains were reweighed to assess their dry mass. The volume of brain edema was assessed by an equation: (wet weight − dry weight)/wet weight × 100%.
Assessment of Cerebral Infarction
The brain tissues were sliced to a thickness of 2 µm and were submerged in a 0.5% 2,3,5-triphenyltetrazolium chloride (TTC) for 10 min, and thereafter subjected to paraformaldehyde fixation. The tissues were then assessed for pigmentation as follows: The color white represented the infarcted region, whereas red signified the non-infarcted area, which was assessed using ImageJ software.
Analysis of Oxidative Stress and Inflammatory Biomarkers
The levels of oxidative stress indicators were evaluated in the brain tissue homogenates. The tissues were homogenized in a tris buffer supplemented with a protease inhibitor solution for 15 min. The homogenate was then centrifuged for 20 min at 10,000 rpm. The resulting supernatant was employed to test the amounts of catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), nitrate, and malondialdehyde (MDA) using commercial kits. The matrix metalloproteinase-2 (MMP-2) and MMP-9 in the brain tissue homogenates were also measured using commercial kits. The manufacturer’s specifications (MyBioSource, USA) were followed for the estimates, which were performed in triplicate.
Assessment of Inflammatory Cytokines
The levels of inflammatory cytokines, especially tumor necrosis factor-alpha (TNF-α), interleukin (IL)-6, IL-1β, and IL-10 in the brain tissue homogenates were quantified in the experimental rats using assay kits. The analysis was performed in accordance with the methods established by the manufacturer (Elabscience, USA).
Analysis of Apoptotic Proteins and Inflammatory Biomarker Levels
The concentrations of apoptotic proteins and inflammatory biomarkers were evaluated in the brain homogenates. The supernatant was utilized to assess the quantities of apoptotic proteins Bax, Bcl-2, and caspase-3 levels using commercial test kits. The evaluations were performed in triplicate following the manufacturer’s procedure (Elabscience, USA). The levels of inflammatory markers nuclear factor kappa B (NF-κB), cyclooxygenase-2 (COX-2), and prostaglandin E2 (PGE2) in rat brain tissue homogenates were evaluated using commercial test kits. The tests were conducted in triplicate as per the manufacturer’s specifications (Abcam, USA).
Statistical Analysis
The values are analyzed using GraphPad Prism. The results were illustrated as the mean ± SD of triplicate measurements, and evaluated by one-way analysis of variance (ANOVA) and Tukey’s post hoc tests. The significance was established with a p value of less than .05.
Results
Effect of Gigantol on Brain Edema and Infarction Levels in the Cerebral Ischemia–Reperfusion Injury-induced Rats
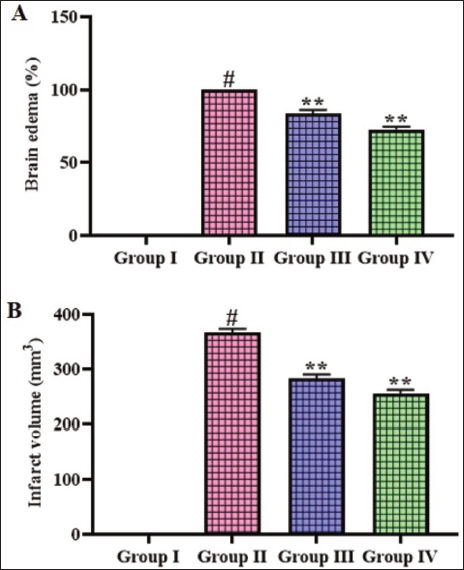
Figure 1 illustrates the effects of gigantol on the brain edema and infarction levels in the CI–RI-induced rats. The rats with MCAO-induced CI–RI had significantly (p < .01) elevated levels of brain edema and infarction when compared with the sham group. Nonetheless, the treatment of gigantol at a concentration of 10 and 20 mg/kg, respectively, displayed significant (p < .05) decrease in both edema and infarction levels in the CI–RI-induced rats.
Effect of Gigantol on the Brain Edema and Infarction Levels in the Cerebral Ischemia–Reperfusion Injury (CI–RI)-induced Rats. The Results were Expressed as the Mean ± Standard Deviation (SD) of Triplicates. The Data were Statistically Analyzed by One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Test Using GraphPad Prism. The Symbol “#” Indicates Statistical Significance (p < .01) in Comparison to the Sham Group (Group I). The Symbol “**” Indicates Statistical Significance (p < .05) in Comparison to the CI–RI-induced Group (Group II).
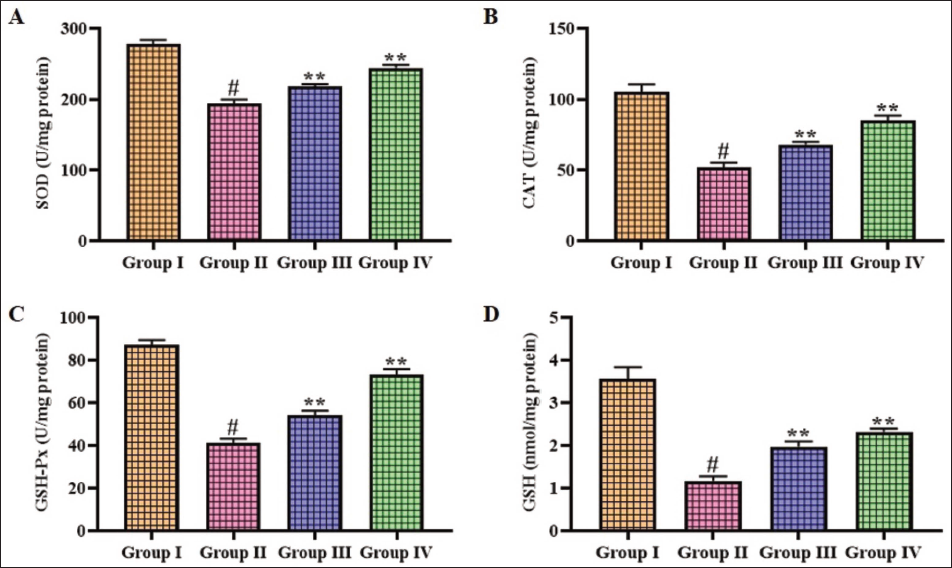
Effect of Gigantol on Antioxidant Levels in Brain Tissues of Cerebral Ischemia–Reperfusion Injury-induced Rats
The concentrations of antioxidants in the brain tissues of experimental rats were analyzed, and the results are illustrated in Figure 2. The current findings illustrate that the SOD, CAT, GPx, and GSH levels were significantly (p < .01) reduced in the brains of CI–RI-induced rats compared to the sham group. Fascinatingly, the gigantol treatment (10 and 20 mg/kg) concentrations markedly elevated (p < .05) the concentrations of these antioxidants in the brains of CI–RI rats.
Effect of Gigantol on the Antioxidant Levels in the Brain Tissues of Cerebral Ischemia–Reperfusion Injury (CI–RI)-induced Rats. The Results were Expressed as the Mean ± Standard Deviation (SD) of Triplicates. The Data were Statistically Analyzed by One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Test Using GraphPad Prism. The Symbol “#” Indicates Statistical Significance (p < .01) in Comparison to the Sham Group (Group I). The Symbol “**” Indicates Statistical Significance (p < .05) in Comparison to the CI–RI-induced Group (Group II).
Effect of Gigantol on the Nitrate, Malondialdehyde, and Matrix Metalloproteinase-2/9 Levels in the Cerebral Ischemia–Reperfusion Injury-induced Rats
Figure 3 depicts the impact of gigantol treatment on the concentrations of nitrate, MDA, and MMP-2/9 in the brains of CI–RI-induced rats. The rats with CI–RI had increased concentrations of MDA, nitrate, and MMP-2/9 in their brains compared to the sham group. Remarkably, the 10 and 20 mg/kg dosages of gigantol treatment effectively decreased (p < .05) the nitrate, MDA, and MMP-2/9 concentrations in the brains of CI–RI-induced rats.
Effect of Gigantol on the Nitrate, Malondialdehyde (MDA), and Matrix Metalloproteinase (MMP)-2/9 Levels in the Brain Tissues of Cerebral Ischemia–Reperfusion Injury (CI–RI)-induced Rats. The Results were Expressed as the Mean ± Standard Deviation (SD) of Triplicates. The Data were Statistically Analyzed by One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Test Using GraphPad Prism. The Symbol “#” Indicates Statistical Significance (p < .01) in Comparison to the Sham Group (Group I). The Symbol “**” Indicates Statistical Significance (p < .05) in Comparison to the CI–RI-induced Group (Group II).

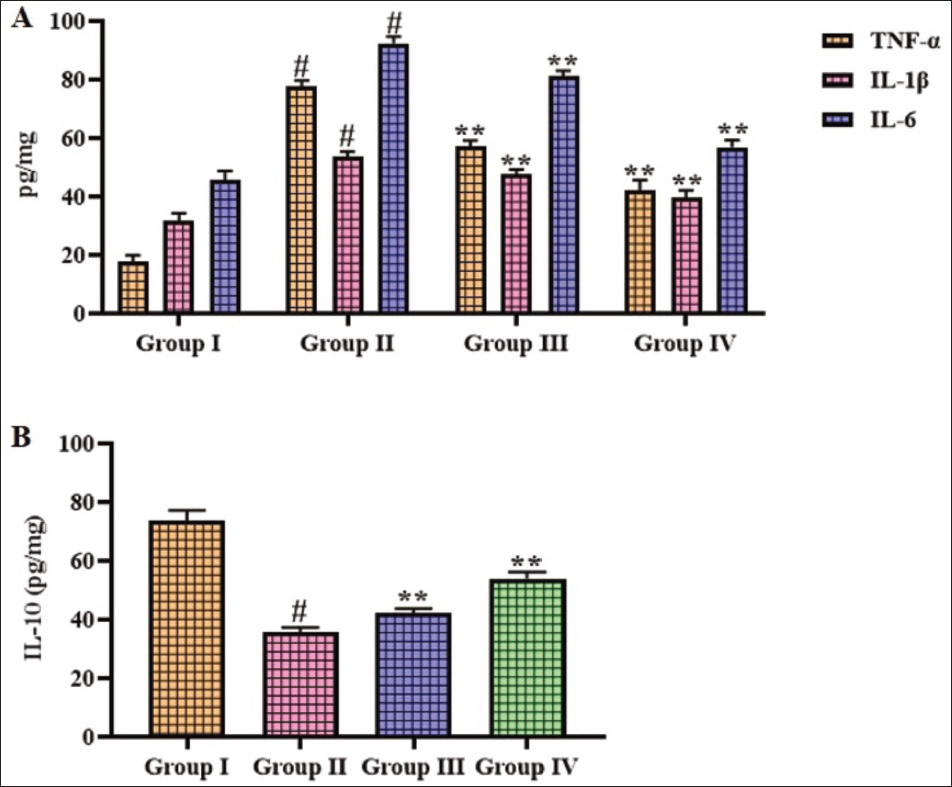
Effect of Gigantol on Inflammatory Cytokines in Brain Tissues of Cerebral Ischemia–Reperfusion Injury-induced Rats
Figure 4 displays the inflammatory cytokines in the brain tissues of experimental rats. Elevated levels of IL-6, IL-1β, and TNF-α, along with diminished IL-10 levels, were seen in the brain tissue homogenates of the CI–RI-induced rats. Interestingly, treatment with 10 and 20 mg/kg of gigantol markedly reduced (p < .05) the pro-inflammatory cytokines and augmented IL-10 in the brain tissues of CI–RI-induced rats.
Effect of Gigantol on the Inflammatory Cytokine Levels in the Brain Tissues of Cerebral Ischemia–Reperfusion Injury (CI–RI)-induced Rats. The Results were Expressed as the Mean ± Standard Deviation (SD) of Triplicates. The Data were Statistically Analyzed Using One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Test in GraphPad Prism. The Symbol “#” Indicates Statistical Significance (p < .01) in Comparison to the Sham Group (Group I). The Symbol “**” Indicates Statistical Significance (p < .05) in Comparison to the CI–RI-induced Group (Group II).
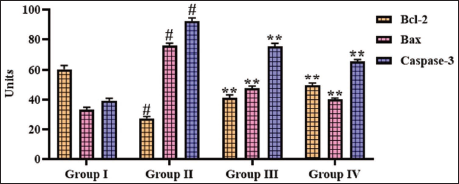
Effect of Gigantol on the Apoptotic Protein Levels in the Cerebral Ischemia–Reperfusion Injury-induced Rats
The impact of gigantol treatment on the apoptotic proteins in the brains of experimental rats was evaluated, with findings depicted in Figure 5. The Bax and caspase-3 concentrations were augmented and subsequently (p < .01) Bcl-2 was reduced in the brains of CI–RI-induced rats relative to the sham group. Whereas the administration of 10 and 20 mg/kg of gigantol considerably reduced (p < .05) both Bax and caspase-3 and augmented Bcl-2 in the brains of the CI–RI-induced rats.
Effect of Gigantol on the Apoptotic Protein Levels in the Brain Tissues of Cerebral Ischemia–Reperfusion Injury (CI–RI)-induced Rats. The Results were Expressed as the Mean ± Standard Deviation (SD) of Triplicates. The Data were Statistically Analyzed by One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Test Using GraphPad Prism. The Symbol “#” Indicates Statistical Significance (p < .01) in Comparison to the Sham Group (Group I). The Symbol “**” Indicates Statistical Significance (p < .05) in Comparison to the CI–RI-induced Group (Group II).
Effect of Gigantol on Inflammatory Markers in Brain Tissues of Cerebral Ischemia–Reperfusion Injury-induced Rats
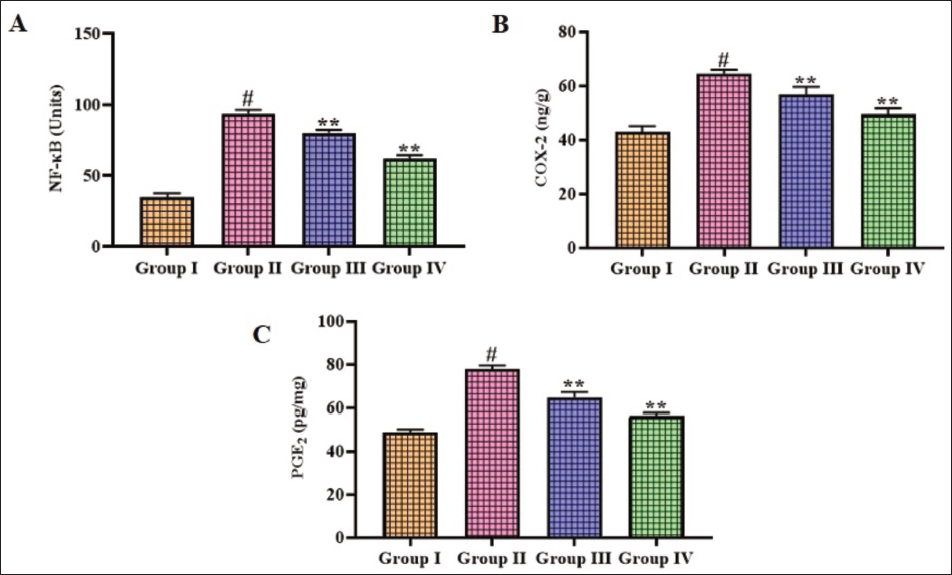
Figure 6 illustrates the concentrations of inflammatory markers, including COX-2, NF-κB, and PGE2, in the brain tissues of rats. The brain homogenates of CI–RI-induced rats displayed increased COX-2, NF-κB, and PGE2 concentrations in comparison with the sham group. Remarkably, the gigantol treatment substantially (p < .05) diminished the concentrations of COX-2, NF-κB, and PGE2 in the brains of CI–RI-induced rats.
Effect of Gigantol on the Inflammatory Marker Levels in the Brain Tissues of Cerebral Ischemia–Reperfusion Injury (CI–RI)-induced Rats. The Results were Expressed as the Mean ± Standard Deviation (SD) of Triplicates. The Data were Statistically Analyzed by One-way Analysis of Variance (ANOVA) and Tukey’s Post Hoc Test Using GraphPad Prism. The Symbol “#” Indicates Statistical Significance (p < .01) in Comparison to the Sham Group (Group I). The Symbol “**” Indicates Statistical Significance (p < .05) in Comparison to the CI–RI-induced Group (Group II).
Discussion
Cerebral ischemic stroke, a critical neurological condition, arises from the blockage of blood delivery to the brain, initiating a complex mechanism that culminates in neuronal injury and functional deficits. The primary pathological event in stroke is brain ischemia, resulting from blockage of blood vessels and consequent neuronal injury (GBD 2016 Neurology Collaborators, 2019). Oxidative stress, defined by a disproportion between the ROS generation and the antioxidant systems, plays an imperative role in the onset of cerebral stroke, exacerbating neuronal damage and contributing to the progression of the disease. Reactive oxygen species (ROS) are produced during I/R, overwhelming the endogenous antioxidant systems and triggering oxidative damage to cellular components. The brain’s higher oxygen utilization, high lipid content, and limited antioxidant systems render it more susceptible to oxidative stress. Mitochondrial dysfunction, excitotoxicity, and neuroinflammation are key mechanisms that contribute to cellular injury following ischemia, with oxidative stress acting as a central mediator in this cascade (Duan et al., 2021). During ischemia, the disruption of the mitochondrial electron transport chain leads to augmented superoxide radical generation, which further exacerbates oxidative stress. MDA, a lipid peroxidation end-product, serves as a widely used indicator of oxidative stress in ischemic stroke. SOD, CAT, GPx, and GSH are key antioxidants that counteract oxidative stress by scavenging ROS and maintaining redox balance. Under normal circumstances, cells maintain a balance between the accumulation and removal of ROS, ensuring their appropriate role in cell signaling and homeostasis (Ramana et al., 2017).
Oxidative stress develops when the ROS generation overwhelms the cellular antioxidant defense systems, leading to cellular damage (Li et al., 2022). SOD catalyzes the dismutation of superoxide radicals into H2O2 and O2, playing a crucial role in mitigating oxidative stress. CAT, another crucial antioxidant that catalyzes the decomposition of H2O2 into H2O and O2, further reducing oxidative stress (Pawluk et al., 2024). GPx, a selenium-containing enzyme, decreases H2O2 and lipid hydroperoxides by utilizing GSH as a substrate, thereby protecting cells from oxidative damage. GSH is a major intracellular antioxidant that directly scavenges ROS and serves as a substrate for GPx. These antioxidant enzymes act synergistically to neutralize ROS and maintain cellular redox balance, protecting neurons from oxidative damage (Orellana-Urzúa et al., 2020). Maintaining antioxidant levels and redox equilibrium within the brain is a crucial aspect of neuroprotection. Therapeutic strategies aimed at enhancing antioxidant defenses and reducing oxidative stress may hold promise to improve outcomes in stroke patients. Understanding the crucial interplay between oxidative stress and antioxidant mechanisms is essential to developing potential neuroprotective methods (Lochhead et al., 2024). The current findings evidenced that the concentrations of antioxidants were markedly diminished and the MDA level was augmented in the brains of CI–RI rats. Captivatingly, the gigantol markedly increased the concentrations of these antioxidants and diminished MDA in the brains of CI–RI rats, which witnesses its antioxidant capacity.
MMPs play an essential role in degrading the extracellular matrix (ECM) and are implicated in several physiological and pathological mechanisms, including cerebral ischemic stroke. Within the MMP family, MMP-2 and MMP-9 have been the subject of intense investigation due to their prominent roles in the context of stroke, particularly in relation to BBB integrity, edema development, and neuronal injury (Wysocka et al., 2023). Cerebral stroke initiates several mechanisms that result in neuronal apoptosis and neurological deficits. In the normal brain, MMPs participate in the maintenance of the ECM, synaptic plasticity, and neurogenesis. However, in the setting of cerebral ischemia, the expression and activity of MMP-2 and MMP-9 are markedly augmented, contributing to the breakdown of the BBB that maintains brain homeostasis. Specifically, the ability of MMPs to digest the basal lamina of capillaries raises the possibility of hemorrhagic conversion of ischemic tissue (Wang et al., 2024). The disruption of the BBB results in the immune cell infiltrations, exacerbating neuroinflammation and contributing to secondary brain injury. Furthermore, MMP-2 and MMP-9 can directly degrade ingredients of the neurovascular unit, like tight junction proteins and ECM components, resulting in vasogenic edema and high intracranial pressure (Saleem et al., 2021). The present results highlighted that CI–RI-induced rats had increased concentrations of MMP-2 and MMP-9 in their brains. Whereas, the gigantol considerably diminished the MMP-2/9 in the brains of CI–RI-induced rats.
Inflammation is recognized as an imperative player in the onset of cerebrovascular diseases, particularly stroke, which arises from arterial occlusion. The interplay between the immune response and the ischemic brain profoundly influences the fate of brain tissue and the survival of stroke patients. The post-ischemic inflammatory cascade is a complex phenomenon involving a multitude of cell types, signaling molecules, and pathways that can either exacerbate or ameliorate neuronal injury (Levinson et al., 2025). Inflammation is defined by the stimulation of microglia, leading to the subsequent generation of inflammatory mediators. Understanding the intricacies of the inflammatory response in stroke is essential to developing potential interventions focused on minimizing brain damage and enhancing neurological recovery (Cheng et al., 2022).
TNF-α, a pro-inflammatory cytokine, contributes to neuronal injury by promoting apoptosis, increasing BBB permeability, and enhancing the generation of other inflammatory markers. Elevated levels of TNF-α are indicative of acute and chronic neuroinflammation (Bai et al., 2023). IL-6, which can exhibit both pro- and anti-inflammatory properties, is upregulated in the ischemic brain and contributes to neuronal damage by promoting excitotoxicity and oxidative stress. IL-1β plays a crucial role in amplifying the inflammatory response by stimulating microglia and astrocytes, promoting the influx of immune cells, and increasing the production of other inflammatory markers. IL-10 is an anti-inflammatory marker that neutralizes the pro-inflammatory cytokines by suppressing microglia activation, inhibiting the pro-inflammatory cytokine production, and enhancing neuronal survival (Doll et al., 2014). The balance between pro- and anti-inflammatory cytokines determines the outcome of ischemic stroke, with an excessive or prolonged pro-inflammatory response leading to exacerbated brain damage and poor neurological outcomes. Targeting inflammatory cytokines represents a hopeful therapeutic method to mitigate brain damage and enhance neurological outcomes in cerebral stroke (Zhang et al., 2019). Here, we found increased levels of IL-6, IL-1β, and TNF-α, accompanied by diminished IL-10, in the brain tissues of rats induced with CI–RI. Whereas, the treatment with gigantol diminished the pro-inflammatory cytokines and elevated IL-10 in the brain tissues of CI–RI-induced rats, which highlights its anti-inflammatory activity.
Apoptosis, or programmed cell death, plays an important role in the pathophysiology of ischemic stroke, contributing significantly to the expansion of the infarct core and the exacerbation of neurological outcomes. While necrosis has long been known as the primary mode of cell death in the ischemic core, apoptosis emerges as a critical mechanism in the penumbral region, where cells are subjected to moderate ischemia (Mao et al., 2022). This region, characterized by salvageable tissue, becomes a battlefield where pro-survival and pro-death signals intricately interact, determining the fate of neurons and ultimately influencing the extent of neurological damage. Understanding the molecular mechanisms governing apoptosis in the context of cerebral ischemia is important to develop effective neuroprotective strategies focused on minimizing brain injury and increasing patient outcomes (Tuo et al., 2022).
The apoptotic pathway involves a highly regulated series of events composed of a family of cysteine proteases (caspases). These caspases act as executioners, dismantling the cell in a controlled and organized manner. Among the key players in the apoptotic cascade are the Bcl-2 family proteins, which act as gatekeepers, regulating the permeabilization of the mitochondrial membrane, an imperative step in the activation of apoptosis (Yan et al., 2021). The Bcl-2 family has both pro-apoptotic (Bax) and anti-apoptotic (Bcl-2) proteins, whose interplay determines the cell’s susceptibility to apoptosis. The intricate balance between pro- and anti-apoptotic proteins is crucial in determining the cell’s fate. In cerebral stroke, the disruption of this balance contributes significantly to neuronal apoptosis (Peng et al., 2022). In this study, we noted that the Bax and caspase-3 concentrations were increased, and subsequently, Bcl-2 was diminished in the brains of CI–RI-induced rats. Notably, the treatment of gigantol markedly decreased both Bax and caspase-3 and increased the Bcl-2 in the brains of the CI–RI-induced rats. These findings highlight that the gigantol inhibited the neuronal cell apoptosis during CI–RI in rats.
NF-κB, a transcription factor, plays an essential role in controlling the gene expression that participates in inflammation, immunity, and cell survival. In the context of cerebral stroke, NF-κB activation is triggered by various stimuli, including oxidative stress, hypoxia, and inflammatory cytokines. NF-κB migrates to the nucleus upon activation, where it interacts with specific DNA sequences and facilitates the transcription of target genes that encode pro-inflammatory cytokines and chemokines (Rubin et al., 2021). The stimulation of NF-κB in microglia and neurons leads to the generation of pro-inflammatory markers that attract immune cells and amplify inflammatory reactions. The dynamic interactions between astrocytes, microglia, and neurons highlight the importance of the neurovascular unit in stroke pathology. This, in turn, exacerbates neuronal damage and contributes to the progression of the ischemic cascade (Zhu et al., 2016). COX-2, an inducible enzyme that synthesizes the prostaglandins, is upregulated in response to inflammatory stimuli, including those present during ischemic stroke. COX-2 catalyzes the conversion of arachidonic acid to PGH2, a precursor for various prostaglandins, including PGE2 (Lacroix et al., 2015). PGE2, a bioactive lipid mediator, exerts diverse effects on various cell types in the brain. The stimulation of brain endothelial cells can induce alterations in the BBB, leading to the discharge of several mediators into the brain. PGE2 can modulate neuronal excitability, synaptic transmission, and neuroinflammation, thereby influencing the outcome of cerebral stroke (Li et al., 2021). The intricate interplay between NF-κB, COX-2, and PGE2 contributes significantly to the onset of cerebral stroke. Targeting these key mediators represents a hopeful therapeutic option for mitigating the detrimental effects of inflammation and improving outcomes following ischemic stroke (Maida et al., 2024). The current results showed increased levels of COX-2, NF-κB, and PGE2 in the brains of CI–RI-induced rats. Captivatingly, the gigantol treatment substantially reduced the concentrations of COX-2, NF-κB, and PGE2 in the brain tissues of CI–RI-induced rats.
Conclusion
The present work revealed the neuroprotective effects of gigantol in rats subjected to CI–RI, attributed to its anti-inflammatory, antioxidant, and anti-apoptotic properties. The treatment of gigantol markedly decreased brain edema and infarction levels. Furthermore, gigantol treatment significantly reduced inflammation and oxidative stress, and enhanced antioxidant activity in rats with CI–RI. The gigantol treatment considerably regulated the apoptotic protein levels in the CI–RI-induced rats. These findings may promote the advancement of gigantol as a potential salutary candidate to alleviate and/or prevent brain damage due to ischemic stroke. Moreover, further research is needed to clarify the precise therapeutic potentials of gigantol in relation to ischemic stroke.
Abbreviations
Footnotes
Declaration of Conflict of Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
This work has been approved by the institutional animal ethical committee by the Seventh Department of Encephalopathy, Xi’an TCM Hospital of Encephalopathy, Xi’an, China.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Informed Consent
Not applicable.