
Abstract
Background
Cadmium is a hepatotoxic heavy metal. Cadmium is known to induce ceramide generation and ferroptosis. Edible insects have recently been touted as a potential functional food. Various studies are being conducted in this regard.
Purpose
This study was designed to investigate whether two types of insects, Tenebrio molitor larvae (TM) and Protaetia brevitarsis larvae (PB), can inhibit cadmium-induced hepatotoxicity.
Materials and Methods
TM and PB powder were extracted using 70% ethanol and diluted with phosphate-buffered saline. TM and PB extracts were cotreated with cadmium in Hep3B cells. To investigate the mechanism, desipramine (an acid sphingomyelinase (ASMase) inhibitor), ferrostatin-1 (a ferroptosis inhibitor), and GW4869 (a neutral sphingomyelinase (NSMase) inhibitor) were used with cadmium. Anti-ferroptosis mechanisms of the TM and PB extracts were demonstrated using Western blot, enzyme-linked immunosorbent assay, MTT cell viability assay, real-time polymerase chain reaction analysis, and 2,2′-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) radical scavenging assay.
Results
Cadmium increased ceramide generation and ferroptosis, concomitant with increased ASMase and NSMase expressions. To understand which mechanism is the upstream pathway, ferrostatin-1, desipramine, and GW4869 were treated with cadmium. Cadmium-induced hepatotoxicity was partially inhibited by ferrostatin-1, which did not affect ASMase expression and ceramide levels. Desipramine, but not GW4869, reduced cadmium-induced cell death and ferroptosis by reducing ceramide levels. Therefore, ceramide generation by ASMase can be an upstream pathway of ferroptosis in cadmium-induced cell death. Interestingly, TM and PB extracts reduced cadmium-induced ceramide increase and ASMase expression, leading to reduced ferroptosis and cell death. TM and PB extracts also had antioxidant properties due to their ability to scavenge reactive oxygen species.
Conclusion
TM and PB have an anti-ferroptosis mechanism against cadmium, which is due to inhibition of ceramide generation by reducing ASMase expression. Therefore, TM and PB can inhibit cadmium-induced hepatotoxicity as functional foods.
Introduction
Cadmium is a toxic heavy metal that is released into the environment through various industrial activities, primarily from industrial and agricultural sources (Genchi et al., 2020). It is also found naturally in soil, minerals, and water and is particularly obtained as a byproduct of zinc production from ZnS (Charkiewicz et al., 2023). The half-life of cadmium in the human body is approximately 16–30 years (Charkiewicz et al., 2023). Cadmium induces various toxicities in the brain, heart, lungs, liver, kidney, bones, and reproductive system and has carcinogenic effects due to increased cell death (apoptosis, necrosis), epigenetic changes, DNA damage, oxidative stress, and abnormal DNA methylation (Charkiewicz et al., 2023). Cadmium induces reactive oxygen species (ROS) generation and mitochondrial dysfunction, leading to loss of mitochondrial membrane potential and caspase-3 activation (Gobe & Crane, 2010). Cadmium exposure exhibited hepatotoxicity and lipid accumulation in the liver (Xie et al., 2024). Inhibition of acid sphingomyelinase (ASMase) resulted in reduced cadmium-induced apoptosis and oxidative stress, thereby mitigating the associated liver damage (Xie et al., 2024).
Sphingolipids are essential lipids that play a crucial role in the structure of cell membranes, characterized by a sphingoid backbone (Park & Park, 2020). Palmitoyl-coenzyme A (CoA) and serine, via the de novo synthesis pathway, are combined with several enzymes in order to form ceramide (Park & Park, 2020). After synthesis, ceramide can be metabolized into a variety of substances, including sphingomyelin when phosphocholine is attached. Sphingomyelin is a component of cell membranes, and it can be hydrolyzed by Sphingomyelinases (SMase) to produce ceramide and phosphocholine (Truman et al., 2014). The activity of SMase is dependent on pH and cellular location, and it can be broadly divided into neutral sphingomyelinase (NSMase) and ASMase (Adada et al., 2016). The NSMase is highly expressed in the plasma membrane and plays a role in ceramide production in response to various signals. In contrast, the ASMase is predominantly located in lysosomes and regulates ceramide production by several stimuli, such as lipopolysaccharides (LPS), infections, alcohol, and Cu2+ (Adada et al., 2016).
Ferroptosis is an iron-dependent atypical form of cell death in which the cell membrane is disrupted by the accumulation of lipid peroxidation products (Xie et al., 2016). It is a mechanism that differs from necroptosis, pyroptosis, and apoptosis, in which cell death is triggered by accumulating iron and ROS (Xie et al., 2016). One of the main characteristics is the breakdown of the antioxidant system due to the degradation or deficiency of glutathione peroxidase 4 (GPX4), and the failure of GPX4 to reduce lipid peroxides leads to the accumulation of lipid peroxides, resulting in the breakdown of cell membranes and cell death (Xie et al., 2016). Erastin, a ferroptosis inducer, binds to voltage-dependent anion channel (VDAC)2 and VDAC3 and increases ROS generation (Xie et al., 2016). Furthermore, it also reduced glutathione (GSH) levels by inhibiting cysteine/glutamate antiporter system (Xie et al., 2016). Both effects increase ferroptosis.
In recent years, edible insects have emerged as a future food source. Edible insects are more environmentally friendly, producing less methane and carbon dioxide, and contain a variety of bioactive compounds with potential health benefits (Devi et al., 2022). In this study, we studied the effects of edible insects on the cadmium-induced hepatotoxicity.
Materials and methods
Materials
Tenebrio molitor (TM) and Protaetia brevitarsis (PB) larvae were obtained from Edible-bug (Busan, Korea) and GreenBugs Co-op. (Sokcho, Korea), respectively. The powder forms of TM and PB larvae were extracted using 70% ethanol at 4°C for 24 h. Extracts were diluted to a concentration of 200 mg/mL using 1× phosphate-buffered saline (PBS) and kept at −20°C for further use. Cadmium (CdCl2), desipramine, GW4869, ferrostatin-1, 2,2′-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS), vitamin C, and anti-α-tubulin antibody (T9026) were purchased from Sigma–Aldrich (St. Louis, MO, USA). Erastin was obtained from Adooq Bioscience (Irvine, CA, USA). Anti-Slc7a11 antibody (12691) was obtained from Cell Signaling Technology (Beverly, MA, USA). Anti-Glutathione Peroxidase 4 (GPX4) antibody (ab125066) was purchased from Abcam (Cambridge, MA, USA). Anti-Mouse-Horseradish Peroxidase (HRP; 115-036-003) and anti-rabbit-HRP (111-035-003) antibodies were obtained from Jackson Laboratory (Bar Harbor, ME, USA).
Hep3B Cell Culture and Cadmium Treatment
Hep3B cells from an 8-year-old black male with liver cancer were cultured in Dulbecco’s modified Eagle’s medium (DMEM, Welgene, Korea) supplemented with 10% fetal bovine serum (Welgene) and 1% penicillin and streptomycin (Capricorn, Germany) at 37°C in a CO2 incubator. Hep3B cells were subcultured every 2 days. For cadmium treatment, Hep3B cells were seeded to approximately 60%–70% confluency and treated with 1 or 10 µM of CdCl2 for 24 h.
Western Blotting
Total proteins from Hep3B cells are extracted using radio-immunoprecipitation assay (RIPA) buffer (50 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, and 0.1% sodium dodecyl sulfate [SDS]) with the addition of protease and phosphatase inhibitors (Sigma–Aldrich) at a ratio of 200:1. In total, 50 µg of protein samples are loaded equally into each lane, separated using SDS-polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred to Nitrocellulose (NC) membranes (Bio-Rad Laboratories, Hercules, CA, USA) through a wet transfer electrophoretic method. The membranes were incubated in a blocking buffer (5% bovine serum albumin in tris-based saline with 0.1% Tween-20 (TBST)) at 4°C. After blocking, the membranes were treated with primary antibody in TBST (1:1,000) overnight at 4°C, then with secondary antibodies (1:10,000) at room temperature for 1 h. Protein bands were visualized using the EzWestLumi Plus Reagent (ATTO Corp., Tokyo, Japan) and analyzed with the ChemiDoc MP Imaging System (Bio-Rad Laboratories).
MTT Cell Viability Assay
Hep3B cells were plated at a density of 3 × 104 cells per well in a 96-well plate, allowed to adhere for 18 h, and treated with 1 or 10 µM of CdCl2 for 20 h. In some cases, Hep3B cells were treated with 1 µM ferrostatin-1, 2 µM desipramine, or 10 µM GW4869 for 24 h, followed by 1 µM of CdCl2 for another 20 h. Finally, MTT solution was added to achieve a final concentration of 0.5 mg/mL to each well, and the cells were incubated for additional 4 h. After the media was removed, 200 µL of dimethyl sulfoxide was added to dissolve the purple formazan crystals. The resulting solution’s absorbance was measured at 540 nm to determine cell viability.
Malondialdehyde (MDA), GSH, Ceramide, and ROS Measurement
The levels of MDA, GSH, and ceramide in Hep3B cells were measured using Lipid Peroxidation (MDA) Assay Kit (Abcam, Cambridge, MA, USA), GSH Assay Kit (Biovision, Milpitas, CA, USA), and Human Ceramide ELISA Kit (MyBioSource, San Diego, CA, USA), respectively. ROS was measured using DCFDA/H2DCFDA - Cellular ROS Assay Kit (Abcam).
Real-time Polymerase Chain Reaction (PCR)
Quantitative PCR was performed as previously described (Kim et al., 2023). The primers are summarized in Supplementary Table S1.
ABTS Radical Scavenging Assay
In total, 7.4 mM ABTS and 2.6 mM potassium sulfate were mixed in a ratio of 1:1. After 24 h, the mixture was diluted to an absorbance of 0.7 at 760 nm. Mixed 500 µL of the diluted solution with 100 µL of various concentrations of TM and PB extracts to obtain values. Vitamin C was used as a positive control due to its antioxidant property. The absorbance was measured using a spectrophotometer at 760 nm. The result was shown as median inhibitory concentration.
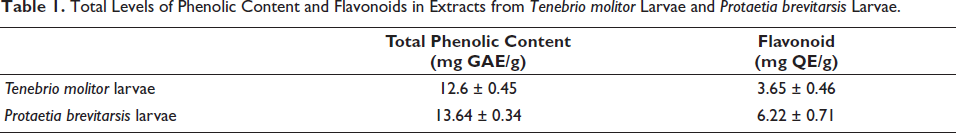
Total Phenolic Content and Flavonoid Content
Total phenolic content (TPC) was performed using the Folin–Ciocalteu method (Kim et al., 2025), and flavonoid content was performed as previously described (Cho et al., 2024). The results of TPC and flavonoid content were represented as mg of gallic acid equivalent per gram (mg GAE/g) and mg of quercetin equivalent per gram (mg QE/g), respectively.
Statistical Analyses
All experiments were repeated independently in triplicate, and values were expressed as mean ± standard error of the mean (SEM). Statistical significance was calculated using Tukey’s post hoc test (GraphPad Prism 6.0; GraphPad Software, San Diego, CA). A statistical value of p < .05 was considered indicative of statistical significance.
Results
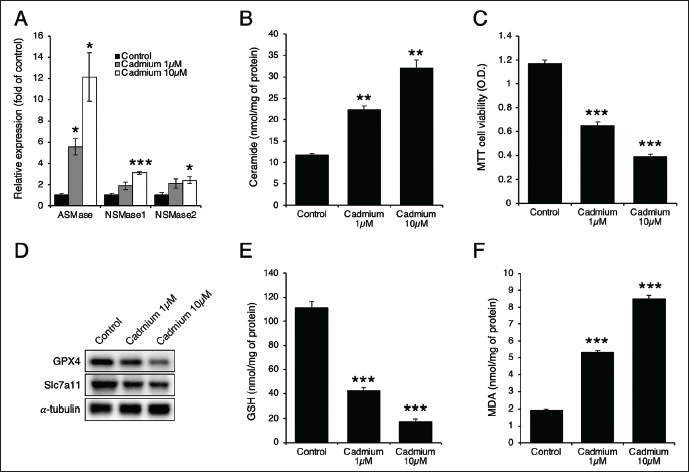
Cadmium Dose-dependently Increased Ferroptosis and Ceramide Generation
A previous study has reported that cadmium induces liver injury by increasing ceramide generation through ASMase (Xie et al., 2024). To explore this, we treated Hep3B cells with 1 or 10 µM of cadmium. The treatment dose-dependently increased the expression of ASMase, NSMase1, and NSMase2, with the most significant increase observed in ASMase (Figure 1A). Additionally, cadmium treatment led to an increase in ceramide levels (Figure 1B) and a decrease in cell viability (Figure 1C). Furthermore, cadmium treatment dose-dependently reduced the levels of GPX4 and soluble carrier family 7 member 11 (Slc7a11) proteins (Figure 1D), decreased GSH levels (Figure 1E), and increased MDA levels (Figure 1F), indicating that cadmium induces ceramide generation and ferroptosis.
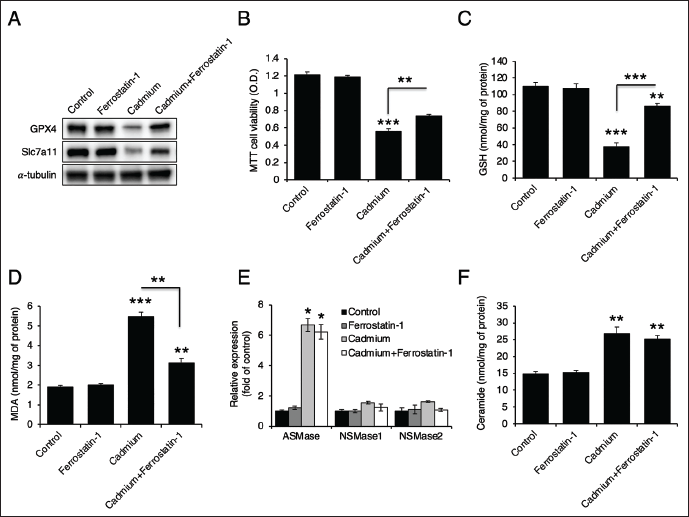
Ferroptosis Inhibition Partially Reduced Cadmium-induced Toxicity
Since cadmium treatment induces ferroptosis (Figure 1), we administered ferrostatin-1, a ferroptosis inhibitor (Miotto et al., 2020). Ferrostatin-1 restored the levels of GPX4 and Slc7a11 (Figure 2A). It also partially reversed cadmium-induced cell death (Figure 2B) and recovered GSH levels (Figure 2C). Additionally, ferrostatin-1 reduced cadmium-induced MDA levels (Figure 2D). However, it had no effect on ASMase expression or ceramide levels (Figure 2E and 2F). These findings suggest that ferroptosis is not upstream of ceramide generation.
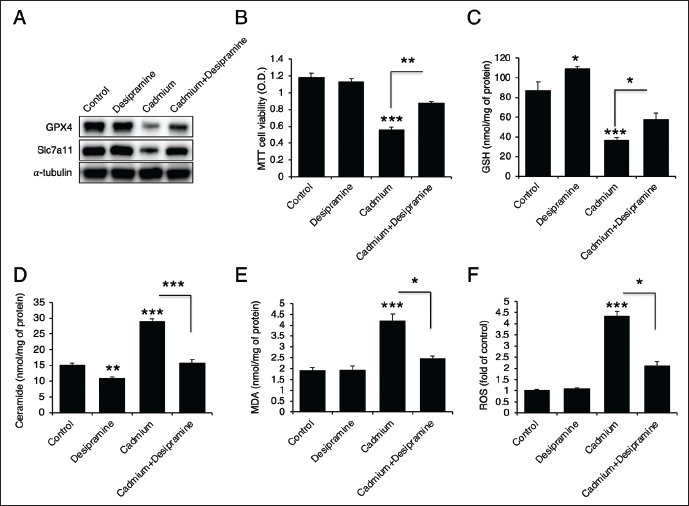
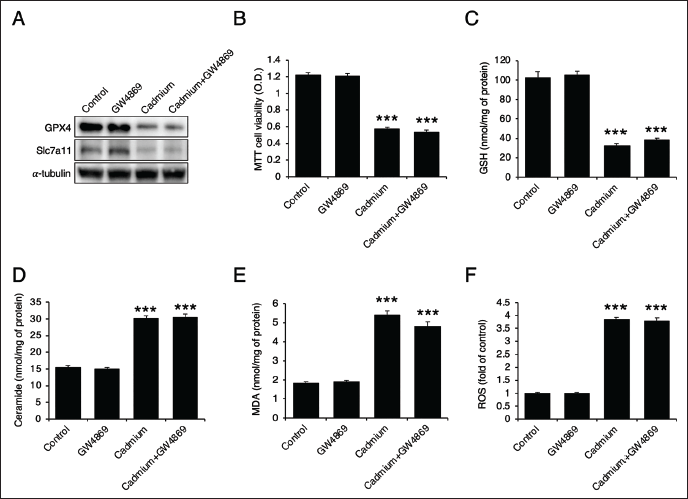
Cadmium Induces Ferroptosis via ASMase
To determine whether ceramide generation is upstream of ferroptosis, we utilized desipramine (an ASMase inhibitor) and GW4869 (an NSMase inhibitor). Desipramine not only recovered the protein levels of GPX4 and Slc7a11 (Figure 3A), but also enhanced cell viability and GSH levels (Figure 3B and 3C). It effectively inhibited ceramide generation (Figure 3D) and reduced both MDA levels and ROS generation (Figure 3E and 3F). These results indicate that ASMase inhibition mitigates cadmium-induced cell death and ferroptosis. On the other hand, GW4869, the NSMase inhibitor, failed to restore the levels of GPX4 and Slc7a11 (Figure 4A) and had no effect on cadmium-induced cell death (Figure 4B). Additionally, it did not recover the levels of GSH, ceramide, MDA, or ROS (Figure 4C–4F), suggesting that NSMase does not play a significant role in cadmium-induced cell death and ferroptosis.
TM Extract Prevents from Cadmium-induced Cell Death and Ferroptosis by Reducing the Expression of ASMase
Given that previous studies have demonstrated the inhibitory effects of extracts from certain edible insects (PB and Gryllus bimaculatus larvae) on ceramide generation (Park et al., 2022, 2024), we treated TM extract with cadmium. Initially, MTT assay was conducted over a range of concentrations to evaluate cytotoxicity induced by the extracts in the Hep3B cells. Both PB and TM extracts exhibited no detectable cytotoxicity at concentrations below 1,000 µg/mL (Supplementary Figure S1). Given that concentrations of 200 µg/mL were previously used in our anti-inflammatory and anti-sarcopenic studies (Oh et al., 2025; Park et al., 2024), we selected 200 µg/mL as the working concentration for the present experiments. Remarkably, TM extract downregulated ASMase expression (Figure 5A) and reduced ceramide generation (Figure 5B) in cadmium-treated Hep3B cells. It also inhibited cadmium-induced cell death (Figure 5C), partially restored GSH and MDA levels (Figure 5D and 5E), reduced ROS generation (Figure 5F), and improved the protein levels of GPX4 and Slc7a11 (Figure 5G). These findings suggest that TM extract mitigates cadmium-induced cell death and ferroptosis.
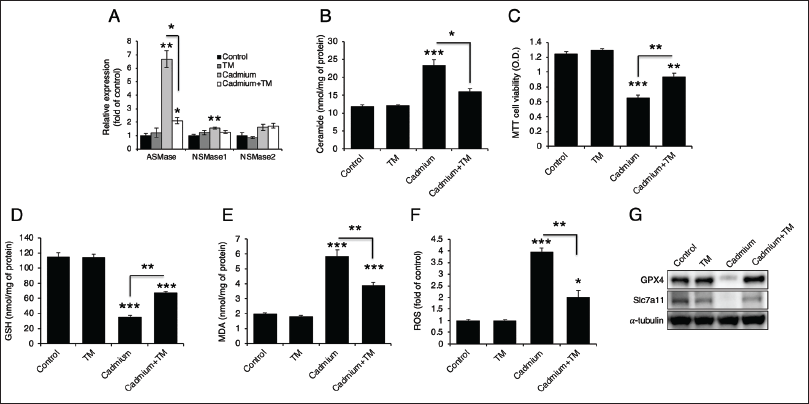
PB Extract Prevents from Cadmium-induced Cell Death and Ferroptosis by Reducing the Expression of ASMase
Because previous research demonstrated that PB extract inhibited LPS-induced ASMase expression (Park et al., 2024), we hypothesized that it might also suppress cadmium-induced ASMase expression. We treated PB extract with cadmium in Hep3B cells. Consistent with the results observed with TM, PB extracts inhibited cadmium-induced ASMase expression (Figure 6A) and ceramide generation (Figure 6B). Additionally, it reduced cadmium-induced cell death (Figure 6C) and restored GSH (Figure 6D) and MDA levels (Figure 6E). It also reduced ROS production (Figure 6F) and recovered the protein levels of GPX4 and Slc7a11 (Figure 6G).
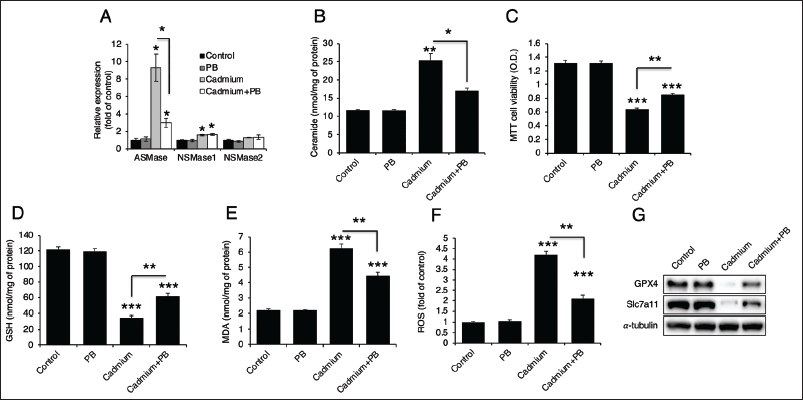
TM Extract and PB Extracts Prevent from Erastin-induced Ferroptosis and Cell Death Due to Their Antioxidants Properties
Because both TM and PB extracts were effective in inhibiting cadmium-induced ferroptosis, we treated them in combination with erastin, a ferroptosis inducer (Thayyullathil et al., 2021), in Hep3B cells. Erastin significantly downregulated both GPX4 and Slc7a11 protein levels, but TM and PB extracts were able to recover these levels (Figure 7A). Erastin treatment also led to increased ASMase expression, ceramide generation, and MDA levels along with decreased cell viability and GSH levels (Figure 7B–7E). TM and PB extracts effectively reversed these effects, restoring ASMase expression, cell viability, and biochemical markers (Figure 7B–7E), suggesting that these extracts can mitigate erastin-induced ferroptosis and cell death.
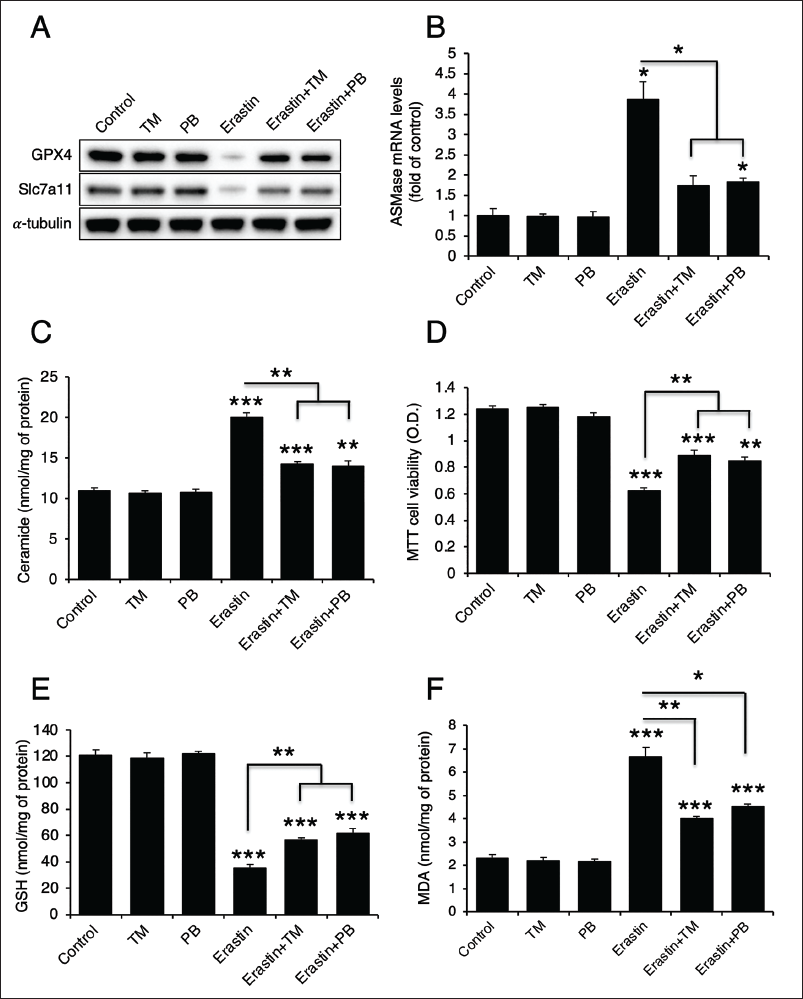
Although TM and PB extracts inhibited cadmium-induced ferroptosis, they did not alter the protein levels of GPX4 and Slc7a11. This led us to hypothesize that these extracts possess antioxidant properties. To confirm this, we performed an ABTS radical scavenging assay. Both TM and PB extracts exhibited dose-dependent antioxidant effects (Figure 8). Furthermore, we quantified the levels of TPC and flavonoids in these extracts, and both TM and PB extracts were found to contain appreciable amounts of these bioactive components (Table 1).
Total Levels of Phenolic Content and Flavonoids in Extracts from Tenebrio molitor Larvae and Protaetia brevitarsis Larvae.
Discussion
Cadmium is a hepatotoxic heavy metal, with chronic exposure leading to itai-itai disease (Inaba et al., 2005). Even at low doses, cadmium can cause liver damage by accumulating in the liver, ultimately resulting in liver fibrosis (Souza-Arroyo et al., 2022). In this study, we investigated the mechanism of acute cadmium toxicity in the liver. Previous studies have highlighted the impact of cadmium on sphingolipid metabolism (Lee et al., 2007; Xie et al., 2024; Xu et al., 2023). Cadmium induces ceramide production, which subsequently activates calpains and caspase-3 in the kidney proximal tubule cells (Lee et al., 2007) and increases ceramide and sphingomyelin contents in mouse midbrain (Xu et al., 2023). A recent study revealed that cadmium-induced ceramide generation is due to activated ASMase (Xie et al., 2024). Our findings also showed that cadmium activates ASMase, leading to increased ceramide production and ferroptosis. Furthermore, desipramine, an ASMase inhibitor, effectively inhibited cadmium-induced cell death and ferroptosis. On the other hand, ferrostatin-1, a ferroptosis inhibitor, partially inhibited cadmium-induced cell death and ferroptosis, but did not affect ASMase expression or ceramide levels. Additionally, erastin, a ferroptosis inducer, was found to activate ASMase, which plays an important role in GPX4 degradation (Du et al., 2023; Thayyullathil et al., 2021). Therefore, ceramide production driven by increased ASMase expression could be an upstream regulator of ferroptosis and play a significant role in cadmium-induced hepatotoxicity.
Cadmium causes hepatotoxicity through multiple mechanisms, including the generation of ROS, mitochondrial dysfunction, and ferroptosis (Rikans & Yamano, 2000). ROS can trigger ferroptosis by promoting lipid peroxidation, a process regulated by the GPX4 and Slc7a11 proteins (Xie et al., 2016). GPX4 is crucial in preventing lipid peroxidation by maintaining oxidized glutathione (GSSG) levels. The ferroptosis inducer erastin degrades GPX4, leading to reduced GSSG levels and increased lipid peroxidation (Xie et al., 2016). The cystine/glutamate antiporter, Slc7a11, also plays a critical role in this process by importing cystine, which is then reduced to cysteine and used for GSH synthesis (Lee & Roh, 2022). Depletion of Slc7a11 accelerates ferroptosis by reducing GSH levels. Extracts of TM and PB, known for their antioxidant properties (Ganguly et al., 2020; Lee et al., 2024; Oh et al., 2022; Yoon et al., 2023), protect against cadmium- and erastin-induced ferroptosis, likely by reducing ROS levels. Although TM and PB extracts do not affect the basal expression of GPX4, SLC7A11, or ASMase, our findings demonstrate that they effectively restore these levels following cadmium exposure. This observation suggests that the extracts act primarily under conditions of oxidative or toxic stress rather than altering homeostatic sphingolipid or ferroptosis-related pathways. The restoration of GPX4 and SLC7A11 is likely attributable, at least in part, to the antioxidant capacity of the extracts, which reduces intracellular ROS accumulation and thereby prevents the ROS-mediated degradation or suppression of these ferroptosis-regulating proteins. In addition, TM and PB extracts markedly attenuate the cadmium-induced upregulation of ASMase, a key enzyme that promotes ceramide generation and contributes to ferroptosis signaling. By suppressing this aberrant ASMase activation, the extracts may limit excessive ceramide accumulation and mitigate downstream cell-death pathways. Taken together, these findings indicate that both antioxidant activity and ASMase-modulatory effects cooperate to confer protection against cadmium-induced cytotoxicity and ferroptosis. This dual mode of action highlights the potential of TM and PB extracts as multifaceted modulators of oxidative stress and sphingolipid-mediated cell-death mechanisms. Other edible insects, such as Gryllus bimaculatus and Allomyrina dichotoma larvae, have also demonstrated antioxidant properties (Kim et al., 2020; Suh et al., 2010), suggesting that most edible insects may possess anti-ferroptosis effects. Because edible insects, including TM and PB, contain phenolic content and flavonoids (Nam et al., 2021; Nino et al., 2021; Yoon et al., 2020), they may contribute to antioxidant activity.
Both TM and PB, which are edible insects, have demonstrated beneficial effects on various diseases. TM is the most extensively studied, showing reductions in adipogenesis and obesity (Seo et al., 2017) and in the development of fatty liver (Ham et al., 2021). It also exhibits anti-inflammatory and antioxidant properties (Oh et al., 2022; Yoon et al., 2023). Fermented PB has been shown to protect against ethanol-induced hepatotoxicity (Lee et al., 2024), and protein hydrolysates from PB have demonstrated anti-obesity and hepatoprotective effects (Lee et al., 2022). Recently, we discovered that PB extract reduced LPS-induced ASMase expression in Hep3B cells (Park et al., 2024). Therefore, both TM and PB can be considered hepatoprotective agents that offer resistance against various toxic substances. Although TM and PB extracts exhibit protective effects against cadmium-induced hepatotoxicity, their development as therapeutic agents beyond functional food applications remains challenging. This limitation arises from the compositional complexity of the extracts, which contain a diverse array of proteins, lipids, phenolic content, and flavonoids, making it difficult to determine which specific component mediates the observed protective effects. Therefore, future studies should prioritize the isolation and identification of the active bioactive compounds responsible for mitigating cadmium-induced hepatotoxicity.
Edible insects are emerging as a crucial future food resource, essential for reducing carbon emissions and promoting environmental sustainability (Devi et al., 2022). They are rich in various antioxidants, which have the effect of scavenging ROS and inhibiting ferroptosis. In particular, their ability to inhibit ceramide synthesis by downregulating ASMase expression is expected to have a protective effect against various liver diseases, such as fatty liver, Wilson’s disease, and cadmium-induced liver fibrosis. Therefore, edible insects, such as TM and PB, hold much promise as a major hepatoprotective functional food in the future.
Footnotes
Abbreviations
ASMase: Acid sphingomyelinase; GPX4: Glutathione peroxidase 4; GSH: glutathione; LPS: Lipopolysaccharides; MDA: Malondialdehyde; NSMase: Neutral sphingomyelinase; PB: Protaetia brevitarsis larvae; ROS: Reactive oxygen species; Slc7a11: Soluble carrier family 7 member 11; SMase: Sphingomyelinase; TM: Tenebrio molitor larvae; TPC: Total phenolic content; VDAC: Voltage-dependent anion channel.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declaration of Conflict of Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval and informed consent
Not applicable.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Supplementary Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.