
Abstract
Background and Purpose
Nutraceuticals are products being used as food and medicines for the treatment of several chronic diseases. The herbal nutraceuticals are gaining importance because of a wide variety of biological actions due to the presence of phytochemicals, such as alkaloids, glycosides, terpenoids, oils, tannins, and several others, like anthocyanins. These anthocyanins are water-soluble pigments found in plants as secondary metabolites and can be of great therapeutic interest for the management of insulin resistance and non-alcoholic steatohepatitis (NASH).
Materials and Methods
Mouse intraperitoneal macrophages primed with lipopolysaccharide (LPS, 1 ng/mL) were subjected to treatment with Oryza sativa L. (black rice) and Ipomoea batatas L. (purple sweet potato) extracts. The evaluation focused on assessing the impact of these extracts on the production of inflammatory cytokines (interleukin-6 [IL-6] and tumor necrosis factor alpha [TNF-α]). Additionally, C57Bl/6 mice were kept on a diet deficient in methionine and choline (MCD) for 8 weeks and were administered O. sativa L. (black rice) and I batatas L. (purple sweet potato) to investigate their effects on NASH.
Results
Extracts obtained from O. sativa L. (black rice) and I. batatas L. (purple sweet potato) demonstrated a significant reduction in inflammatory cytokines, as indicated by a marked decline in TNF-α and IL-1β levels in LPS-primed macrophages. Mice on the MCD diet showed NASH phenotype, and treatment with the extracts of black rice and purple sweet potato significantly (p < .05) lowered plasma liver enzyme levels and inflammation, as measured by mRNA analysis, and attenuated fibrosis, as measured by histopathology.
Conclusion
Collectively, the findings of the study validated the hypothesis of O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extract could improve inflammation-associated NASH.
Introduction
Particularly in some ethnic groups in the USA, non-alcoholic fatty liver disease (NAFLD) has surpassed chronic hepatitis C infection, the most common cause for the development of cirrhosis and chronic liver disease (Setiawan et al., 2016; Younossi et al., 2011). NAFLD is currently the primary source of abnormal aminotransferase levels. The incidence of NAFLD has been progressively rising over the past 25 years, likely mirroring the increasing incidence of insulin resistance, obesity, and type 2 diabetes across the population (Charlton, 2004; Yilmaz & Younossi, 2014). Approximately 30% of the overall population and a higher proportion, ranging from 60% to 80%, within the type 2 diabetic population are estimated to be affected by NAFLD (McPherson et al., 2015). NAFLD is a spectrum condition that includes non-alcoholic fatty liver (NAFL), a non-progressive hepatic steatosis with low inflammation, NASH, and a progressive variant. In contrast, NASH, an advanced form, requires early intervention. While 15%–20% of NASH patients may develop cirrhosis, recent studies suggest that fibrosis progression is not exclusive to NASH and may also occur in NAFL (Singh et al., 2015). Biologically active lipid peroxidation products and pro-inflammatory cytokines, such as interleukin-6 (IL-6) and tumor necrosis factor alpha (TNF-α), synergistically contribute to the initiation of many hepatic lesions in NASH. Their actions induce hepatic inflammation and fibrosis, ultimately progressing to end-stage liver disease (Alisi et al., 2010). In the development of hepatic steatosis, inflammation plays a crucial role, and toll-like receptor (TLR)-4 signaling is implicated as the pro-inflammatory response for triggering NASH. Acute hepatic injury triggers modulators like transforming growth factor β and platelet-derived growth factor (PDGF), amplifying synthesis of collagen through SMAD and non-SMAD signals, and intersecting with pathways like c-Jun terminal kinase (JNK) to promote fibrosis of liver. Cytokines and growth factors activate signaling pathways, involving transcriptional factors like NF-κB, contributing to liver fibrogenesis (Dela et al., 2005; Yoshida & Matsuzaki, 2012).
Natural remedies are now gaining the attention of researchers to treat several disorders based on the phytocompounds present in them (Pratap & Pattnaik, 2023). Numerous compounds are undergoing testing for NASH. Regrettably, reported trials to date have not shown significant improvements in disease status, with many compounds failing to demonstrate effectiveness. Enhancing the likelihood of success in clinical trials could involve the development of robust preclinical models that accurately reflect the disease in humans. Testing various options for efficacy in both preclinical models and clinical settings may provide a more reliable approach.
Choices for treatment for NASH currently are limited to weight loss, lifestyle, and dietary modifications, as well as the use of lipid-lowering drugs and insulin-sensitizing agents, such as pentoxifylline, vitamin E, pioglitazone, and others. Of all these treatments, pioglitazone has shown improvement in NAFLD score in a few clinical studies. Additionally, pioglitazone also decreases ad-lib fed blood glucose and fasting blood glucose levels by improving insulin peripheral sensitivity in type 2 diabetes patients and is currently used as an anti-diabetic medication. Given the complexity (multifactorial) of NASH, single mechanism-based treatments have limited improvements (Masterjohn & Brubo, 2012). Many scientific documents have suggested the beneficial effects of various polyphenolic compounds in experimental NASH models (Hu et al., 2012; Ramirez-Tortosa et al., 2009). Additionally, the flavonol mixture (silymarin treatment) has demonstrated favorable effects in patients with NAFLD.
Anthocyanins are particularly intriguing for their profusion in a diverse range of edible plants, and their appeal lies in the substantial amounts that can be consumed through plant-based daily diets. Many of the anthocyanins, because of their ability to scavenge free radicals, exhibit marked anti-inflammatory activity. The inflammation is because of the release of inflammatory mediators like prostaglandins, histamine, nitric oxide, and oxygen free radicals by the cyclooxygenase pathway. Examples for some anthocyanins having anti-inflammatory activity include cyanidin, delphinidin, petunidin, malvidin, and pleagonidin (Joven et al., 2012; Schrieber et al., 2011). Black rice extract treatment in rats fed a high-fat diet (HFD) showed significant improvement in elevated plasma glucose and lipid levels, as well as increased liver enzyme activities (aspartate transaminase and alanine transaminase). Black rice extract-treated rats showed amelioration of the liver injury, accompanied by lower inflammatory and apoptotic markers (Laorodphun et al., 2024). Peonidin-3-glucoside and cyanidin-3-glucoside are anthocyanins that have demonstrated promising antioxidant and anti-inflammatory activities to lower the symptoms of liver damage induced by carbon tetrachloride (CCl4) in mice hepatic fibrosis model. Cyanidin-3-glucoside and peonidin-3-glucoside were expressed in large amounts in the bran portion of black–purple rice and may contribute to beneficial effects due to their the antioxidant properties (Laorodphun et al., 2024). Similarly, it is reported that a class of naturally occurring anthocyanins in purple sweet potato color (PSPC) has shown significant improvements in many features of HFD-induced NAFLD in a mouse model (Su et al., 2020).
Therefore, based on this evidence, the present research was carried out to further investigate the theory that extracts from O. sativa L. (black rice) and I. batatas L. (purple sweet potato), which are rich in anthocyanins and could potentially provide protection against steatosis. Additionally, the study sought to examine whether these extracts could mitigate the release of pro-inflammatory cytokines and genes related to fibrosis in a mouse model of NASH caused by feeding a methionine and choline-deficient (MCD) diet. In this study, pioglitazone was used as a comparator since several studies have demonstrated that treatment of NASH with pioglitazone shows a significant ameliorative effect through modulation of insulin resistance, inflammation, apoptosis, and necroptosis pathways.
Materials and Methods
Animals
C57BL/6J male mice of 7–10 weeks of old, weighing between 23 and 26 g body weight were utilized for the experiment. All procedures were followed by the Committee for the Control and Supervision of Experiments on Animals (CCSEA), India. This experimental protocol and procedures performed herein have been approved by the Institutional Animal Ethics Committee (IAEC) of Eurofins Advinus Ltd. (approval no: Renewal 022_June-2021). Mice were provided ad libitum feed containing MCD diet (Cat # A02082002B, Research Diet®, USA), and control mice were fed with standard rodent maintenance diet 1334P formulated by Altromin Spezialfutter GmbH & Co. KG, Im Seelenkamp 20, 32791 Lage, Germany.
Experimental Designs and Protocols
Extraction of O. sativa L. (Black Rice) and I. batatas L. (Purple Sweet Potato)
Extraction of O. sativa L. (black rice): In total, 100 g of black rice was cleaned and de-dusted using a vacuum. The rice was coarsely powdered using a grinder (Preethi Steele Supreme 750), and the powdered rice was extracted with 500 mL of hydroethanol (80:20). The blend was kept in a water bath, and the contents were refluxed for 3 h. The blend was then filtered with Whatman No. 1 filter paper, and the filtrate was heated to evaporate in vacuo to 100 mL and then transferred to an evaporating dish, where it was left to dry at room temperature. The dried sample (~4 g) was further pulverized into powder in a mortar and pestle (Pratap & Pattnaik, 2024a; Tan et al., 2022).
Extraction of I. batatas L. (purple sweet potato): In total, 100 g of skin was peeled and cleaned, and de-dusted using a vacuum. Peel was coarsely powdered using a grinder (Preethi Steele Supreme 750), and the powder was transferred to a 3-L round-bottom flask and extracted with 500 mL of hydroethanol (80:20). The blend was kept in a water bath, and the contents were refluxed for 3 h. The blend was then filtered with Whatman No. 1 filter paper, and the filtrate was heated to evaporate in vacuo to 100 mL and then transferred to an evaporating dish, where it was left to dry at room temperature. The dried sample (~4 g) was further pulverized to powder in a mortar and pestle (Tan et al., 2022).
Proof of Concept Assay in Lipopolysaccharide (LPS)-primed Peritoneal Macrophages Derived from C57BL/6 Mice
C57BL/6 male mice were treated with a single injection of phosphate-buffered saline (PBS) intraperitoneally, and after 10–15 min, mice were anesthetized with isoflurane anesthesia and then sacrificed by cervical dislocation. The peritoneal cavity was cut open, and PBS in the cavity was aspirated using a syringe. This aspirated PBS was transferred to a Falcon tube and centrifuged at 6,000 RPM. Supernatant was discarded, and the pellet containing macrophages was isolated. These macrophages were then placed in Dulbecco’s Minimum Essential Medium (DMEM) containing 10% fetal bovine serum (FBS) and 10 mg/mL gentamicin at 37°C in 5% CO2/95% air atmosphere for culturing. The viability of cultured peritoneal macrophages was assessed by assessing cell morphology under a microscope, and there was ~80% yield from the total collected macrophages. These macrophages were pretreated with different concentrations (0.3–6 mg/mL) of extracts of O. sativa L. (black rice) and I. batatas L. (purple sweet potato) and then stimulated with 1 ng/mL of LPS. Then the macrophages were incubated overnight, and the supernatants were collected. These supernatants were used for the estimation of TNF-α and IL-6 inflammatory cytokines using enzyme-linked immunosorbent assay (ELISA) kits as per the manufacturer’s manual for the respective kit for the procedure (Chinta et al., 2022).
Chronic In Vivo Animal Study
C57BL/6 male mice of 7–10 weeks old weighing between 23 and 26 g were provided ad libitum with MCD, and control mice were provided with a regular chow diet throughout the experimental period. Animals were randomly assigned based on body weight to different treatment groups (n = 6) as shown in Table 1.
Different Animal Groups and Treatments were Administered in a Chronic In Vivo Animal Study.
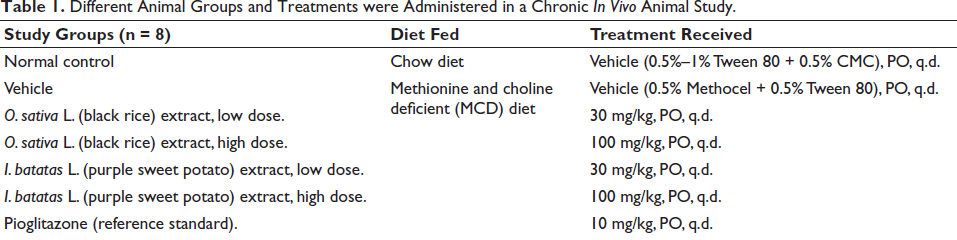
All the animals were treated once daily from day 1, along with initiation of MCD diet feeding in the morning around 9–10
Blood samples, collected in K2EDTA with 20 µL of a 200 mM solution per mL of blood, were placed on ice before plasma collection. Within 30 min of collection, all samples were centrifuged at 5,000 g for 5 min at 4°C ± 2°C. After centrifugation, the separated plasma was kept at −70°C. Samples of plasma were examined for liver enzymes and plasma triglyceride (TG) levels. The liver was collected and weighed. Liver tissue collected was used for the estimation of mRNA biomarkers for various genes, like IL-6, TNF-α, transforming growth factor beta (TGF-β), alpha smooth muscle actin (α-SMA), tissue inhibitor of metalloproteinase-1 (TIMP-1), and collagen 1α (Col1-α), liver TGs, and histopathological analysis (Deng et al., 2013; Gotoh et al., 2011; Marcolin et al., 2012; Weltman et al., 1996).
Plasma Measurements
After 56 days of treatment, plasma samples were collected, and alanine aminotransferase (ALT), aspartate aminotransferase (AST), and TG concentrations were measured with commercially available diagnostic kits (Yao & Vance, 1990).
Histopathological Analysis
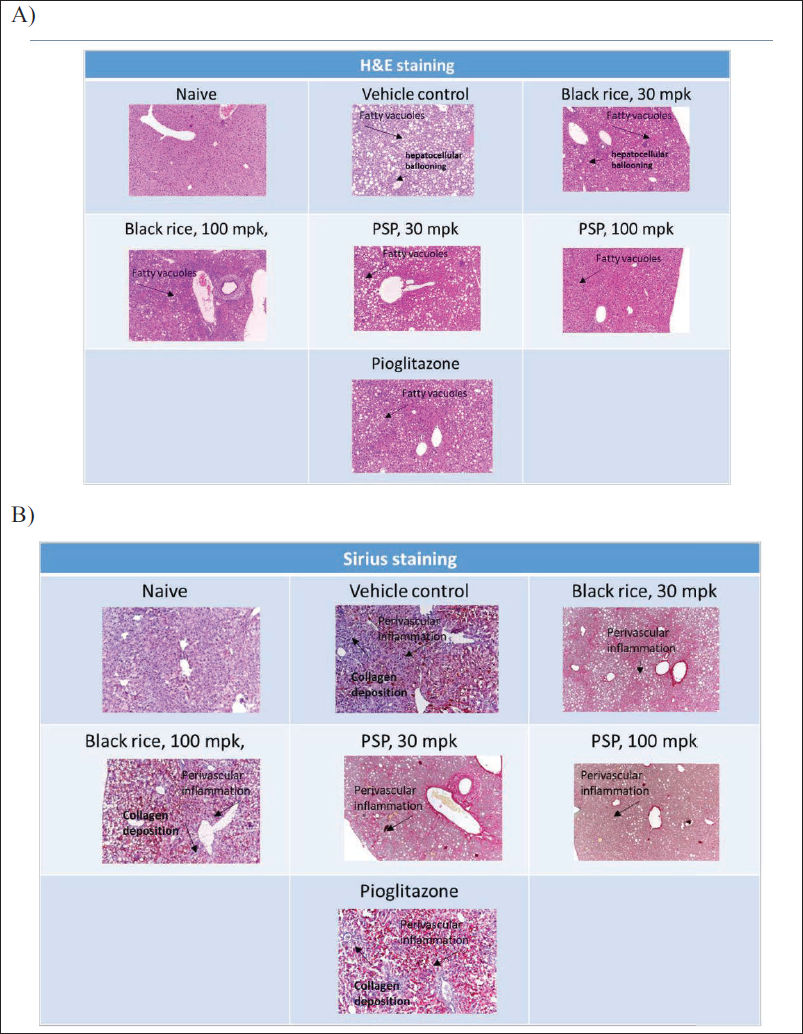
Collected liver samples were assessed for histopathological examination. Histopathological scoring was carried out by a histopathologist blinded to the treatment groups. Microscopic analysis involved staining of liver section slides with hematoxylin-eosin and Sirius red. Blinded slide assessment was conducted by a veterinarian pathologist. The diagnosis of NASH hinged on the histological criterion of steatosis accompanied by ballooned hepatocytes in zone 3 and lobular inflammatory infiltrate (Larter et al., 2008). The slide scoring was carried out as a 0–4 scale for fibrosis (Sirius red stain), a 0–3 scale for inflammation (H&E stain), and a 0–3 scale for degeneration based on intralobular degeneration; H&E stain (Leclercq et al., 2000).
Biomarker Analysis by mRNA Estimations in Liver Tissue
Part of the liver sample collected and stored at the end of the treatment period was used for the estimation of mRNA levels of various genes as markers of fibrosis and inflammation. Liver samples were collected in reagent RNALater® and stored as per the manufacturer’s recommendation until further use. RNA from liver samples was extracted using the RNAeasy kit (Qiagen). mRNA was quantified, and cDNA was prepared using the SuperScript III kit (Invitrogen). qPCR experiments were set up using 100 ng of cDNA, gene expression master mix, Taqman (Applied Biosystems), and a real-time PCR machine, Step One Plus (Applied Biosystems). Ct values were obtained, and data were analyzed using the ∆∆Ct method. All sample data were normalized to the respective GAPDH (housekeeping gene) data. Markers analyzed were TNF-α, IL-6, α-SMA, TGF-β, TIMP-1, and Col1-α for inflammation and fibrosis (Browning & Horton, 2004; George et al., 2003; Marcolin et al., 2012; Richardson et al., 2007; Weltman et al., 1996).
Statistical Analysis
All the values in graphs are presented as mean ± standard error of the mean (SEM). All the datasets generated in this study were analyzed using one-way analysis of variance (ANOVA) with p < .05, and post hoc comparison of Dunnett with control was used.
Results
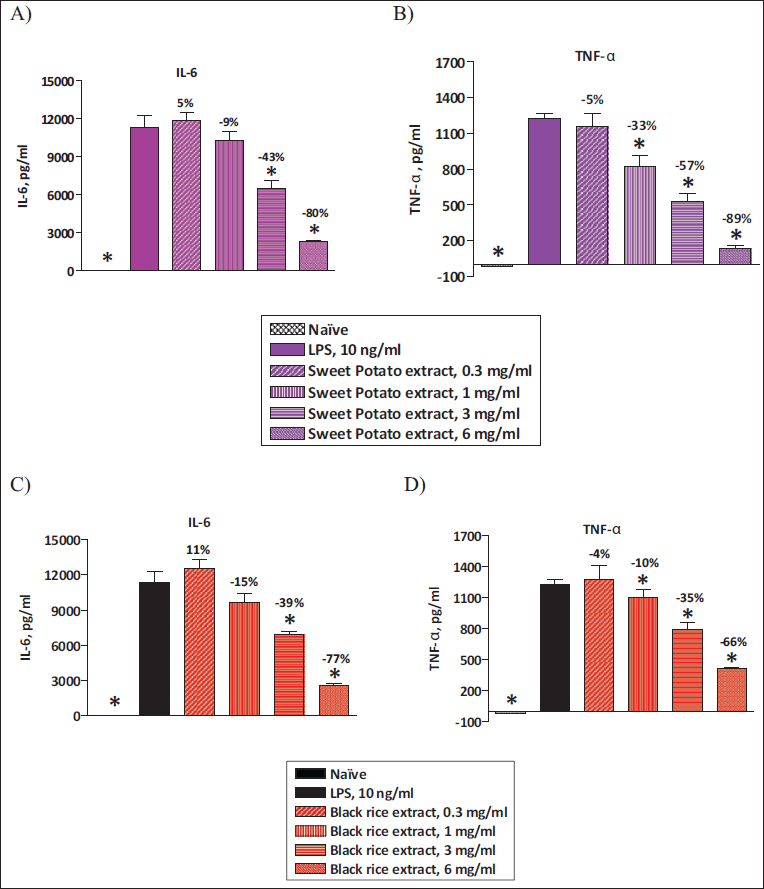
O. sativa L. (Black Rice) and I. batatas L. (Purple Sweet Potato) Extracts Inhibit LPS-induced IL-6 and TNF-α Levels in Intraperitoneal Macrophages (IPMACs)
In order to check if O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts have an inhibitory effect on inflammatory cytokines, we analyzed TNF-α and IL-6 levels by ELISA. Resident peritoneal macrophages were incubated with either vehicle or various concentrations of O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts. Then these macrophages were stimulated with 1 ng/mL of LPS. Both IL-6 and TNF-α levels were significantly elevated in LPS-primed IPMACs. Both O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts showed significant inhibition of IL-6 and TNF-α in LPS-primed macrophages in a dose-dependent manner (Figure 1A–1D). These data suggest that O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts containing anthocyanins have a potential inhibitory effect on inflammatory cytokines.
Effect of O. sativa L. (Black Rice) and I. batatas L. (Purple Sweet Potato) Extracts Treatment on Plasma Liver Enzyme Levels and Plasma TG Levels
At the end of 8 weeks of treatment, animals were fasted for 6–8 h, and blood samples were collected for estimation of plasma liver enzyme levels. Feeding of animals with the MCD diet led to a significant elevation in serum AST and ALT levels in comparison with animals fed with the chow diet. Daily treatment with 100 mg/kg daily doses of O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts significantly improved plasma liver enzyme levels (ALT and AST) in comparison with vehicle treatment animals. Pioglitazone treatment at a 10 mg/kg/day dose also showed a significant improvement in plasma AST and ALT levels in comparison with vehicle treatment. The extent of improvement in plasma liver enzymes is similar to pioglitazone treatment, indicating the potential beneficial effects of extracts (Figure 2A and 2B).
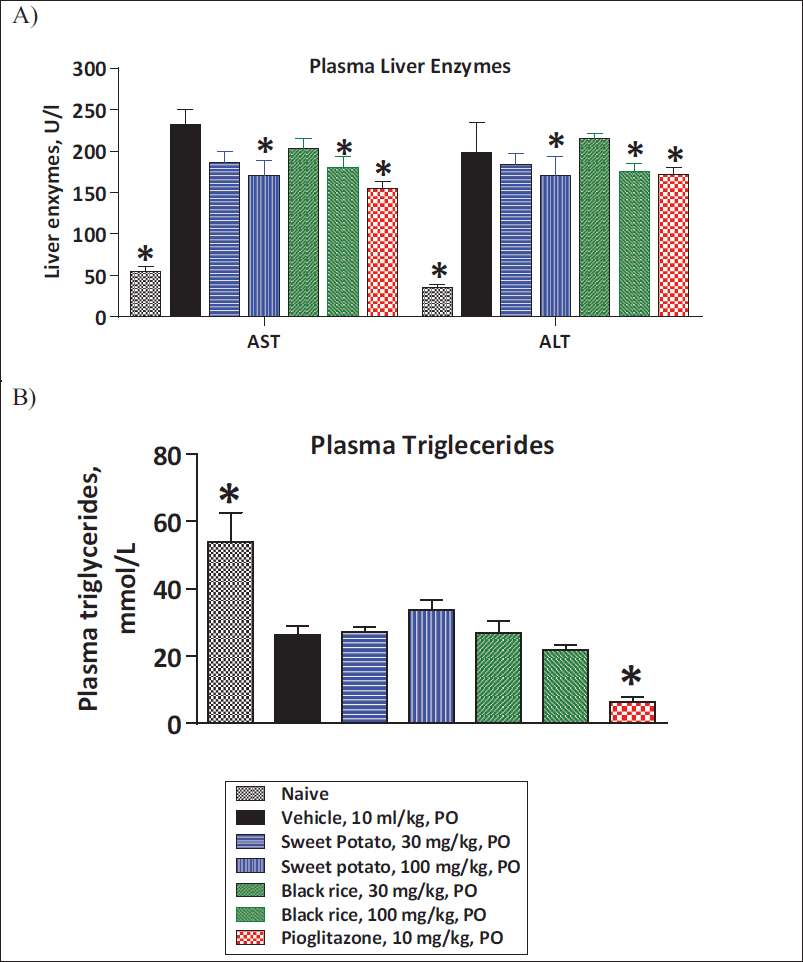
MCD diet feeding in mice caused a significant reduction in plasma TGs as compared to control animals. Treatment with black rice and purple sweet potato extracts displayed no significant change in plasma TG levels when compared to vehicle treatment. However, pioglitazone showed a significant reduction in plasma TG levels in comparison with vehicle treatment (Figure 2A and 2B).
Effect of O. sativa L. (Black Rice) and I. batatas L. (Purple Sweet Potato) Extracts Treatment on Liver TGs, Liver Weight, and Histopathological Analysis
MCD diet feeding for 8 weeks in mice also led to a significant elevation in liver TGs and a significant decrease in liver weight in comparison with control animals. Treatment with O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts did not show a significant change in liver TGs and liver weight as compared to vehicle treatment. However, pioglitazone treatment at a 10 mg/kg/day dose also did not show a significant change in liver TGs and liver weight as compared to vehicle treatment (Figure 3A and 3B; Laghezza et al., 2023, 2025).
The MCD diet is known to cause a significant increase in hepatic fibrosis and increased inflammation in comparison to control animals. In our study, we also observed significant derangement of liver tissue as assessed by gross pathology (Figure 4) and histopathological evaluation by H&E staining. O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts showed a significant reduction in inflammation score as compared to vehicle-treated animals (Figures 3D and 5) and a trend towards reduction in fibrosis (this was not statistically significant; Figure 3C). This indicates the potential beneficial effects of extracts in reducing inflammation and thereby fibrosis. In this study, treatment was carried out for 8 weeks, and the treatment duration may not be sufficient. Longer duration of treatment may show beneficial effects in reducing fibrosis as well (Pratap Singh & Pattnaik, 2024b).
Gross Representative Photographs of Liver at the End of Treatment Showing Macroscopical Changes in the Liver.
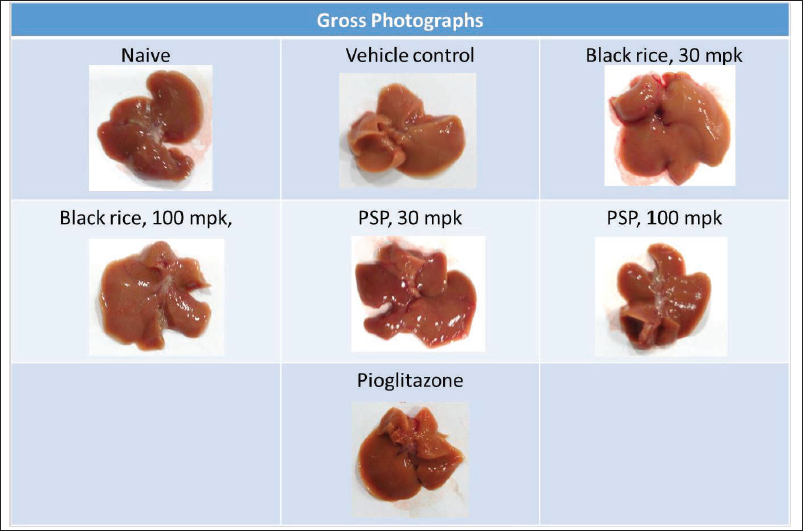
Pioglitazone treatment demonstrated a considerable decrease in fibrosis score and inflammation score as compared to vehicle treatment (Figures 3C, 3D, 3F, and 5). MCD diet feeding is also known to lead to an increase in hepatocyte degeneration and rarefaction as compared to control animals, and it was also observed in this study. O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts did not show significant change in hepatocyte degeneration and rarefaction as compared to vehicle treatment. Pioglitazone considerably declined hepatocyte rarefaction as compared to vehicle treatment (Figures 3E, 3F, and 5).
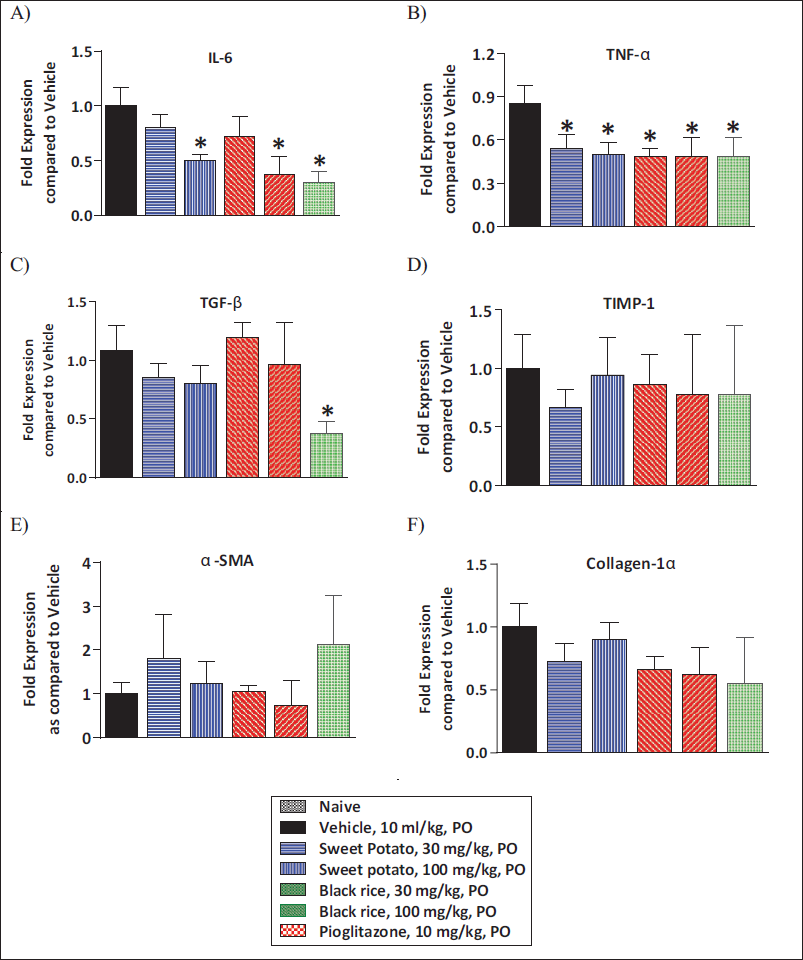
Effect of O. sativa L. (Black Rice) and I. batatas L. (Purple Sweet Potato) Extracts Treatment on Liver mRNA Levels of Various Markers
Part of the liver tissue collected was also used for the estimation of various markers of inflammation and fibrosis by mRNA analysis. MCD diet feeding caused a notable rise in mRNA levels of inflammation-related genes IL-6 and TNF-α as compared to control animals. O. sativa L. (black rice) and I. batatas L. (purple sweet potato) showed significant amelioration in IL-6 and TNF-α levels in comparison with vehicle treatment. Pioglitazone treatment at a 10 mg/kg/day dose showed a significant reduction in IL-6 and TNF-α levels as compared to vehicle treatment (Figure 6A and 6B). The data of fibrosis-related gene expression demonstrated a significant elevation in mRNA levels of Col1-α and α-SMA, TGF-β, and TIMP-1 levels in comparison with control animals. O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts did not show significant reduction in α-SMA and Col1-α, TGF-β, and TIMP-1 levels as compared to vehicle treatment (Figure 6C–6F).
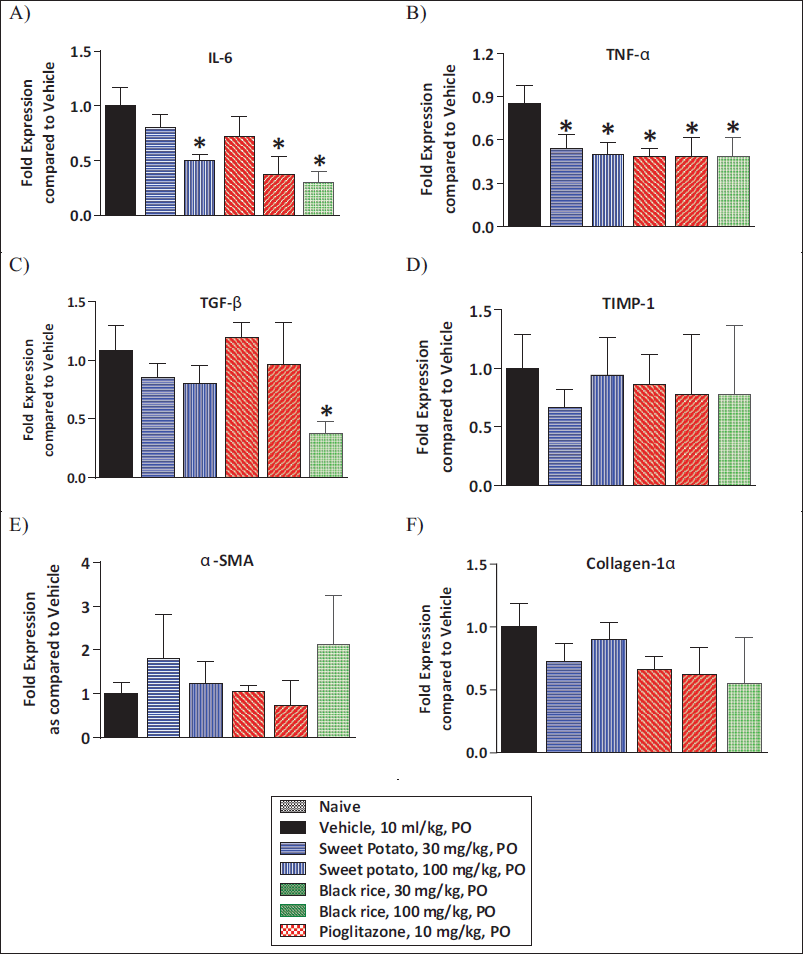
Pioglitazone treatment (10 mg/kg/day) displayed a prominent reduction in TGF-β levels and no change in α-SMA and Col1-α TIMP-1 levels as compared to vehicle treatment (Figure 6C–6F).
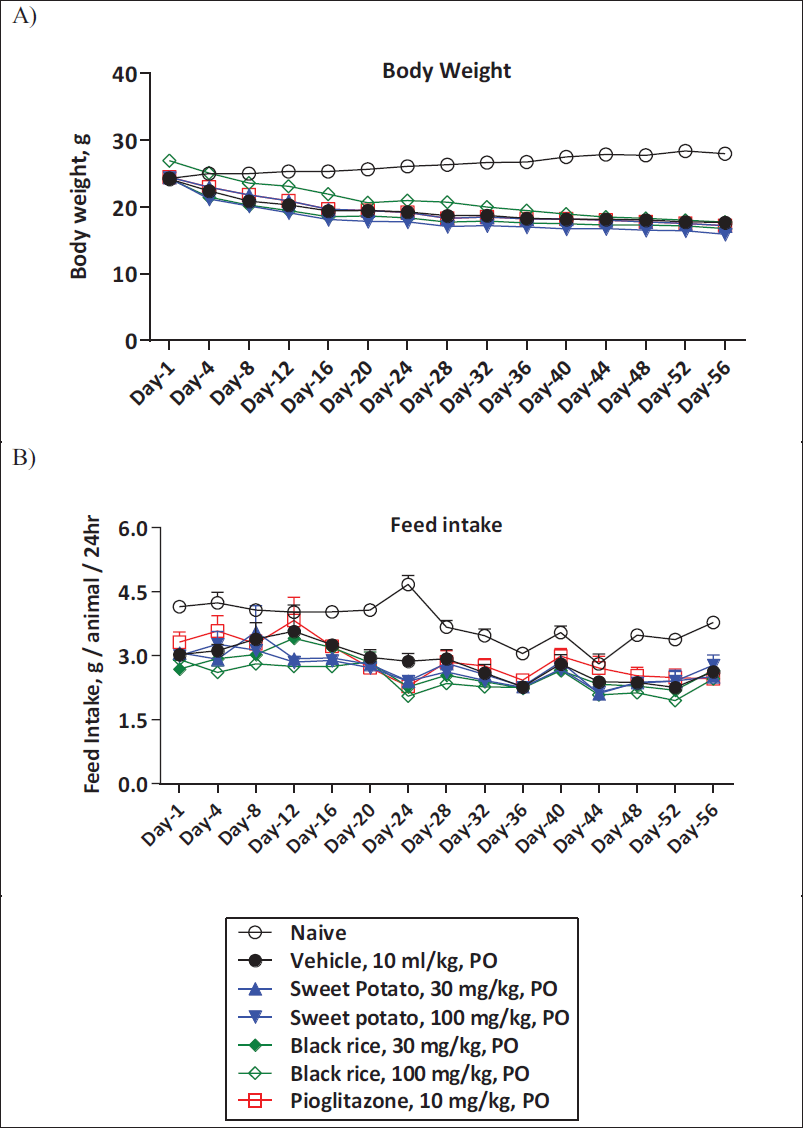
Effect of O. sativa L. (Black Rice) and I. batatas L. (Purple Sweet Potato) Extract Treatment on Body Weight and Feed Intake
MCD diet-fed animals are cachectic and show significant weight loss (muscle and fat weight). In our study, MCD diet-fed animals showed a significant decrease in body weight and feed intake as compared to control animals. O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts demonstrated no prominent effect on body weight and feed intake in comparison with vehicle treatment. Similarly, pioglitazone treatment displayed no significant alterations in body weight and feed intake (Figure 7A and 7B).
Discussion
Inflammation plays a very significant role in the development of insulin resistance, and its implication on hepatic diseases like steatohepatitis is well documented. In our study, we demonstrated the inhibitory effect of these extracts on the release of cytokines in LPS-primed macrophages, supporting our hypothesis.
To further demonstrate the translation of the observed in vitro effects to in vivo correlation, we utilized a diet lacking in important amino acids, such as choline and methionine, which is widely recognized as a method to induce NASH. The MCD diet consists of high fat and high sucrose content (10% fat, 40% sucrose) and is deficient in methionine and choline, which are essential amino acids crucial for hepatic β-oxidation and very low-density lipoprotein (VLDL) production. Choline deficiency further hinders hepatic VLDL secretion, leading to heightened hepatic fat accumulation and diminished VLDL production. Additionally, oxidative stress and shifts in cytokines and adipokines further contribute to liver injury within this model. C57BL/6J mice on MCD diet develop NASH features, including necrosis, hepatic fibrosis, and inflammation. The MCD diet-induced mouse model is widely used as it mimics the histopathological features observed in human NASH, which include inflammation, fibrosis, and stress responses. Mice exposed to the MCD diet exhibit an elevation in inflammatory responses marked by triggering liver macrophages through activation of nuclear factor kappa B (NF-κB), a transcriptional factor pivotal in regulating inflammatory and cell survival responses. Although the MCD diet more accurately reflects the pathobiological mechanisms driving the progression of human NAFLD to advanced NASH, it does not fully mimic the metabolic profile or weight regulation observed in human NASH.
Simultaneously, there is an elevation in TNF-α, IL-6, IL-1β, and TGF-β levels, mirroring various aspects of the disease in humans. This encompasses a histological profile that reflects human fibrotic disorders linked to hepatic lipid accumulation, inflammation, and oxidative stress (Richardson et al., 2007).
Fat building up in the liver is considered the triggering event in the initiation of NASH. According to a number of studies, hepatic steatosis causes increased hepatic lipogenesis. This can be due to either impaired fatty acid oxidation or increased synthesis of TGs, which both raise the amount of TG in the liver. In our study, there was a minimal effect of O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts. This may be due to the shorter duration of treatment, or the doses tested were lower in this study and therefore exerted only a minimal effect in reducing the TG accumulation in the liver.
During MCD diet administration, the pathogenic cascade begins with lipid accumulation and peroxidation, followed by hepatocellular injury and inflammation, activation of hepatic stellate cells (HSCs), and upregulation of profibrotic genes. Two interrelated processes, oxidative stress and the subsequent cytokine release, may further drive fibrogenesis. However, while inflammation has been proposed as a prerequisite for fibrosis development, the chronological and interdependent relationship between inflammation and fibrosis in NASH still requires a more comprehensive understanding. TNF-α, an inflammatory cytokine, significantly contributes to the progression from steatosis to NASH and induces the secretion of various other cytokines and chemokines, such as IL-6 and matrix metalloproteinase 9 (MMP-9) (31). Among these, TGF-β emerges as a key mediator, exerting a pivotal influence on hepatic fibrogenesis by activating HSCs. The activation of HSCs by TNF-α and TGF-β leads to the expression and deposition of alpha-smooth muscle actin (α-SMA) and collagen. In our study, we demonstrate that O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts lower mRNA levels of hepatic TNF-α and IL-6 and have minimal effect in lowering TGF-β mRNA levels that are higher after all MCD feeding. The heightened expression of α-SMA and extracellular matrix-related factors, including collagen I-alpha, in the livers of mice on MCD diet remained unaltered following treatment with O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts. Elevated levels of MMP and TIMP are typically observed in fibrotic diseases. A decline in the TIMP:MMP expression ratio is crucial for impeding matrix degradation and remodeling, favoring the deposition of extracellular matrix. This increased expression of TIMP-1 was unchanged with O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts. Findings from this study indicate that the second hit may have been attenuated by O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts, which only improved liver inflammation but had not much impact on the fibrosis induced by the MCD diet feeding. This may be due to the shorter duration of treatment, and a longer duration of treatment may be required to demonstrate the beneficial effects on fibrosis as well.
After 8 weeks on the MCD diet, mice exhibit pronounced hepatic macro vesicular steatosis, marked intralobular inflammatory foci, elevated hepatic lipid content, and increased serum ALT and AST levels. In our study, MCD diet feeding to mice caused a significant increase in hepatic fibrosis, increased inflammation, an increase in hepatocyte degeneration, and rarefaction as compared to control animals. O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts showed significant reduction in inflammation score and trend towards reduction in hepatic fibrosis, no significant changes in hepatocyte, degeneration, and rarefaction compared to vehicle treatment, confirming the role of these extracts in reducing hepatic inflammation and mild anti-fibrotic effects.
Elevated ALT and AST levels may indicate hepatic steatosis, inflammation, or fibrosis, particularly when they exceed four times the upper normal limit. In our study, the MCD diet also caused a significant elevation in serum AST and ALT levels, and this effect was significantly attenuated with O. sativa L. (black rice) and I. batatas L. (purple sweet potato) extracts treatment. The extent of reduction in plasma liver enzymes was similar to that of the pioglitazone treatment, indicating the beneficial effects of the extracts for NASH treatment.
Several publications have shown beneficial effects of resveratrol, curcumin, or silymarin in various models of liver fibrosis and NSAH. The potential effects of the extracts in this study for O. sativa L. (black rice) and I. batatas L. (purple sweet potato) represent a value addition to the various polyphenolic treatments in reducing the progression and treatment of such diseases.
Conclusion
The MCD diet is a well-established dietary model for investigating NASH.
Rodents fed this diet develop steatohepatitis, necroinflammation, and fibrosis resembling the pathology seen in human NASH. The MCD diet is characterized by reduced glycogen stores, protein peroxidation, increased reactive oxygen species (ROS) generation, mitochondrial DNA damage, and apoptosis, making it one of the most widely accepted models for studying NASH-related inflammation, oxidative stress, and fibrosis. Our results showed that extracts of O. sativa L. (black rice) and I. batatas L. (purple sweet potato) containing large amounts of anthocyanins contribute to the reduction of multiple pro-inflammatory gene pathways, imparting beneficial effects in mice with MCD diet-induced steatohepatitis. Despite the constraints associated with this animal model of NASH and the consideration that pharmacological doses of these extracts were examined, further elucidation of molecular pathways and potential clinical applications is warranted. Further long-term studies at higher doses in another animal models of NAFLD/NASH shall be performed to elucidate their usefulness in the development of pharmacotherapy for treatment in humans.
Footnotes
Abbreviations
ALT: Alanine aminotransferase; ANOVA: Analysis of variance; AST: Aspartate aminotransferase; CCSEA: Committee for the Control and Supervision of Experiments on Animals: Col1: Collagen 1 alpha; ELISA: Enzyme-linked immunosorbent assay; H&E; Hematoxylin and eosin; IAEC: Institutional Animal Ethics Committee; IL-6: Interleukin 6; MCD: Methionine and choline deficient; mg/kg: Milligram per kilogram; Na-CMC: Sodium carboxymethyl cellulose; NASH: Non-alcoholic steatohepatitis; TG: Triglycerides; TIMP-1: Tissue inhibitor of metalloproteinase-1; TGF-β: Transforming growth factor beta; TNF-α: Tumor necrosis factor-alpha; αSMA: Alpha smooth muscle actin.
Acknowledgments
Suhas Tambe conducted the research work, wrote, and authored this article, while P. Sivakami Sundari and Geetha Kannoth Mukundan conceptualized the study and provided guidance and analysis of the research outcomes.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
The study design and procedures were approved by the Institutional Animal Ethics Committee (IAEC) of Eurofins Advinus Limited (approval no.: Renewal 22_June-2021).
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.