
Abstract
Background
Meditation is known to modulate neural activity and enhance cognitive and emotional well-being. While previous studies have largely reported increases in alpha and theta activity, the neurophysiological effects of structured sound-based meditation practices remain underexplored.
Purpose
This study investigated the neurophysiological and subjective effects of rhythmic sound meditation (RSM) compared to a resting state (RS) condition. We aimed to identify EEG spectral patterns and changes in subjective alertness during this meditation protocol.
Methods
Fifteen healthy adults (mean age = 24.8 ± 3.6 years) participated in two 26-minute sessions (RSM and RS) in a randomised, counterbalanced design. EEG data were recorded using a 64-channel actiCHamp system, and power spectral density was analysed across delta, theta, alpha, beta, and gamma bands. Subjective alertness was assessed before and after the sessions using the Stanford Sleepiness Scale (SSS). Statistical analyses were conducted using Yuen’s paired t-test with FDR correction.
Results
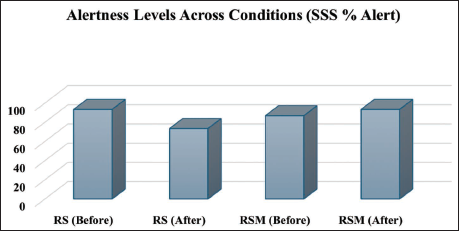
RSM reduced EEG power across all frequency bands, particularly in prefrontal and frontocentral regions, compared to RS (p < .05, FDR-corrected). Power reductions were observed across delta, theta, alpha, beta, and gamma bands, with the most pronounced effects in frontal areas. SSS scores showed increased alertness post-RSM (93.3% alert) versus decreased alertness post-RS (73.3% alert).
Conclusion
RSM induced distinct neurophysiological changes, characterised by widespread EEG power suppression in frontal regions and heightened subjective alertness. These findings support its potential for promoting altered states of consciousness, contributing to the neuroscientific understanding of sound-based meditation.
Introduction
Meditation is a way to mentally train oneself, aiming to cultivate essential psychological skills like concentration and sustained attention. Meditation techniques are associated with better mental health, including enhanced mood and decreased anxiety, depression, and stress levels.1, 2 Functional and structural neuroimaging studies revealed neuroplastic changes induced by meditation, including increased cortical thickness and grey matter density in critical brain areas responsible for self-awareness, attention, and emotional regulation. These areas include the prefrontal cortex, anterior cingulate cortex, hippocampus, and insula.3–5 Moreover, meditation is associated with decreased activity in the default mode network (DMN), while enhancing connectivity within executive control networks, indicating better cognitive integration and focused attention during meditative states.6, 7
Electroencephalography (EEG) offers an insightful perspective on the dynamic brain states caused by meditation, tracking changes in neural oscillatory activity across particular frequency bands. For instance, different meditative techniques exhibit increased theta and alpha activity, attributed to a state of relaxed alertness, focused internal attention, and a decrease in mind-wandering.8, 9 However, lower beta and gamma power is attributed to decreased cognitive effort and reduced sensory processing, highlighting a shift toward states of deep internal focus and passive awareness or concentrated internal focus.10, 11
However, the EEG signatures of meditation are not uniform across practices. While focused attention and open monitoring practices typically show increase in alpha and theta power,9, 12 states characterised by ‘mental emptiness’ or reduced cognitive processing demonstrate opposite patterns. Hinterberger et al. (2014) observed that experienced meditators achieving a conscious state of ‘thoughtless emptiness’ exhibited widespread decreases in EEG power across all frequency bands, including delta, theta, alpha, beta, and gamma. 13 Similarly, Duda et al. (2024) reported that alpha power decreased significantly during meditation compared with rest, particularly in novice practitioners. 14 This reduction in oscillatory activity has been interpreted as reflecting decreased cortical engagement with task-irrelevant cognitive processes, rather than a passive or hypoactive state.15, 16
The divergence in EEG patterns appears to depend on the meditation technique’s cognitive demands and phenomenological targets. Sound-based practices involving rhythmic auditory stimulation may engage distinct neural mechanisms compared to silent meditation forms. Moreover, power suppression during meditation may reflect enhanced neural efficiency and reduced mind-wandering, analogous to task-related decrease in DMN activity. 17
Among various forms of meditation, sound-based practices, especially those invoking the sacred syllable ‘AUM’, hold a foundational role in Vedic and yogic traditions.18–21 Neuroscientific studies on AUM chanting have demonstrated diverse EEG signatures depending on the practice context and measurement timing. Some studies report increased theta and alpha power following AUM chanting,22, 23 while others show increased theta relative power with decreased theta frequency during OM chanting sessions. 24 Notably, fMRI studies reveal that AUM chanting produces significant deactivation of limbic regions, including the amygdala, orbitofrontal cortex, anterior cingulate, and parahippocampal gyri, suggesting vagal nerve stimulation and parasympathetic activation.23, 25
Mantra-based practices more broadly show context-dependent neural signatures. Studies on Gayatri mantra chanting and other repetitive religious chanting have demonstrated modulation of late-stage brain responses to emotional stimuli and changes in autonomic balance. 25 The effects appear influenced by factors including chanting duration, audibility (silent vs. vocalised), participant experience level, and the specific measurement approach employed. This heterogeneity underscores the need to characterise sound-based meditation practices with well-controlled experimental designs.
The current study examines rhythmic sound meditation (RSM), a structured practice rooted in the Nāda Bindu Upanishad, a seminal ancient text describing progressive inward auditory concentration on increasingly subtle inner sounds, culminating in silence and transcendental awareness.26, 27 We aimed to characterise the neurophysiological signatures of RSM in meditation-naïve University students compared to a resting baseline.
Based on the theoretical framework outlined above, we hypothesised that RSM would produce distinct neurophysiological signatures compared to resting state (RS). Specifically, we predicted: (a) reduced EEG power across frequency bands, particularly in prefrontal regions associated with self-referential processing, based on studies demonstrating power suppression during ‘mental emptiness’ states and task-related neural efficiency13, 17; (b) enhanced subjective alertness despite reduced oscillatory activity, consistent with evidence that meditation-induced power suppression reflects active attentional engagement rather than drowsiness15, 28; and (c) frontocentral topographic distribution reflecting the cognitive demands of auditory attention and breath awareness inherent in RSM. These predictions contrast with conventional meditation literature emphasising power increases but align with emerging evidence that different meditation phenomenologies engage distinct neural mechanisms.29, 30
Materials and Methods
Participants
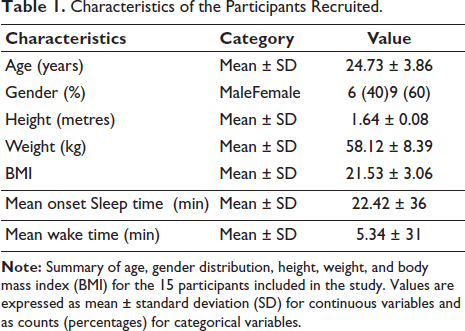
Fifteen healthy adult participants (nine females, six males; age range: 17–33 years; mean ± SD = 24.8 ± 3.6) with no prior meditation experience were recruited for the study. This sample size was determined based on feasibility constraints and the exploratory pilot nature of this investigation. The within-subject repeated-measures design enhances statistical power, as each participant serves as their own control, reducing inter-individual variability. Our sample size of N = 15 is comparable to other published pilot studies examining meditation-related EEG changes.10, 23, 31 We acknowledge this as a preliminary investigation aimed at establishing feasibility and generating hypotheses for future adequately powered confirmatory studies.
All the participants were fluent in English and capable of comprehending and responding to standardised self-report measures. The recruitment was conducted via pamphlet-based advertisements at the Central University of Rajasthan (CURAJ) in India. Participant characteristics, including age, gender, height, weight, BMI, sleep and waking time, are detailed in Table 1. Prior to inclusion, each individual underwent a brief clinical screening conducted by a certified medical officer at the University to ensure the absence of any neurological, psychiatric, or chronic medical conditions. None of the participants reported current or past use of psychoactive medications, recreational drugs, or any ongoing medical treatment. During screening, participants were asked to report any history of participation in meditation retreats, workshops, or courses, regular meditation or mindfulness practice, yoga practices involving meditation components, and use of meditation apps or guided practices. Only individuals reporting no such experiences were included. The study received approval from the Institutional Ethics Committee of CURAJ (CURAJ/H-IEC/24/12/008). Written informed consent was obtained from all the participants following a detailed explanation of the research objectives before their involvement in the study.
Characteristics of the Participants Recruited.
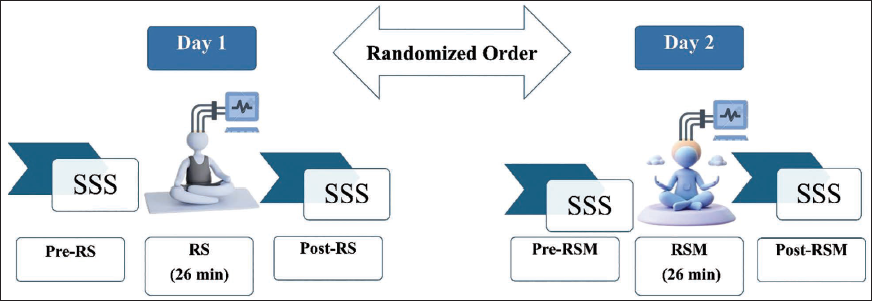
Experimental Design
Each participant completed two experimental sessions, RSM and RS, in separate sessions held on different days. In addition, all the participants were asked to report to the Laboratory for acclimatisation before the actual data acquisition of RSM and RS. Participants were randomly assigned to one of two session order sequences using a computer-generated randomisation list: (a) RSM followed by RS, or (b) RS followed by RSM, with sessions separated by at least 48 hours (range: 2–7 days) to minimise carryover effects. The randomisation was implemented by the study coordinator (not involved in data analysis). Seven participants received RSM first (RSM-RS sequence), and eight participants received RS first (RS-RSM sequence). The study employed a single-blind design wherein participants were naïve to the specific hypotheses regarding EEG power changes and directional predictions. They were informed only that the study aimed to ‘compare brain activity during different mental states’. Participants were not told which condition was expected to produce particular effects, minimising expectancy effects on subjective measures. However, complete blinding was not feasible given the distinct nature of the interventions (guided meditation vs. quiet rest). The EEG technician who prepared participants and acquired data was not involved in data preprocessing or analysis. Both sessions were matched in duration, with each session lasting 26 minutes, during which continuous EEG data were recorded. In the RS condition, participants were instructed to remain relaxed with eyes closed, allowing thoughts to flow naturally without attempting to focus on any particular object or suppress emerging thoughts. In RSM, participants practised a meditation protocol, a 26-minute guided auditory meditation centred around structured attention to the sound of AUM. This meditative session was similarly conducted with eyes closed in a quiet environment. To assess immediate changes in subjective alertness, the Stanford Sleepiness Scale (SSS) 32 was administered before and after both sessions. Figure 1 schematically illustrates the procedural flow of the experimental design.
RSM Intervention
The RSM is a 26-minute structured auditory-based meditative practice rooted in the Nāda Bindu Upanishad.26, 27 It guides the practitioner through a progression of inner auditory awareness, focusing on the syllable AUM, and transitioning from external sound perception to internal silence. The technique comprises seven sequential stages: (a) Starting Prayer, (b) Kapalbhati (high-frequency breathing with active exhalation followed by passive inhalation), (c) Breath Awareness, (d) Contemplation on one’s own emotion, (e) Sound Concentration on AUM, (f) Silence with automatic repetition of the AUM (Ajapa japa), and (g) Closing prayer.
EEG Data Acquisition
EEG signals were acquired using a 64-channel actiCHamp system (Brain Products GmbH, Germany) following the international 10–20 montage. The reference electrode was placed at FCz, and the ground at AFz. The data were sampled at 1000 Hz, and electrode impedance was maintained below 30–40 kΩ throughout the recording as recommended by the vendor. Instructional stimuli were presented using E-Prime 2.0 software for stimulus delivery (Psychology Software Tools, Inc., Sharpsburg, PA, USA). Participants were positioned at a fixed distance of 90 cm from a 34 cm × 27 cm LCD monitor, which continuously displayed task instructions throughout the experimental protocol.
All recordings took place in a sound-attenuated, electromagnetically shielded chamber, with an ambient temperature of 25°C and humidity controlled between 40% and 60%. To ensure high-quality EEG recordings, participants were instructed to wash their hair and refrain from consuming coffee, tea, or any other beverages known to affect EEG signals prior to the session. During both the resting and meditation conditions, participants were seated comfortably on a cushioned chair with their arms and feet resting, their eyes closed, and were instructed to remain as still as possible to minimise motion-related artefacts.
Subjective Sleepiness Assessment
To assess participants’ momentary subjective alertness, the SSS32, 33 was administered before and after each experimental condition. The SSS is a single-item, 7-point Likert scale ranging from 1 (‘Feeling active, vital, alert, or wide awake’) to 7 (‘No longer fighting sleep, sleep onset soon; having dream-like thoughts’). We selected the SSS for its brevity (<30 seconds), which minimises state changes between the meditation session and EEG recording, and its demonstrated use in previous meditation research. 34 In the present study, a categorical scoring scheme was applied to enhance interpretability. Specifically, responses from categories 1 and 2 were grouped into an ‘Alert’ classification, while selections from categories 3 through 7 were grouped as ‘Not Alert’, following prior convention in cognitive alertness research. This binary classification was used to compare the impact of RS versus RSM on subjective alertness.
EEG Preprocessing
Preprocessing was conducted using EEGLAB v2024.2 in MATLAB R2024a. 35 The EEG data were first resampled from 1,000 Hz to 250 Hz and re-referenced to the average reference. Raw continuous data were subjected to a zero-phase finite impulse response (FIR) bandpass filter (0.5–45 Hz, filter) to attenuate slow drifts and high-frequency noise while preserving physiological signals of interest. Channels with excessive noise, flat signals, or poor contact (indicated by abnormally high or low variance relative to neighbouring channels) were identified through visual inspection and statistical outlier detection. Artefact-contaminated channels and segments were automatically identified and removed using the Artefact Subspace Reconstruction (ASR) method (threshold = 5 SD). Subsequently, independent component analysis (ICA) with the ‘infomax’ algorithm was performed, and components corresponding to scalp muscle, ocular, ECG, and respiration artefacts were rejected using the ICALABEL plug-in (90% confidence threshold). Previously identified bad channels were interpolated using spherical spline interpolation to restore the complete 64-channel montage for topographic analysis. A trained researcher visually inspected these components to verify accurate classification, after which they were excluded to maintain data integrity. The remaining components were backprojected to reconstruct the cleaned EEG signal for further analysis.
EEG Spectral Analysis
Power spectral density (PSD) was calculated for each EEG channel using MATLAB’s band power function, employing a Hamming window and a frequency resolution of 0.5 Hz. PSD values were extracted for the canonical EEG frequency bands: delta (1–4 Hz), theta (4–8 Hz), alpha (8–12 Hz), beta (13–30 Hz), and gamma (30–40 Hz). To normalise the distribution of power values and reduce skewness, log-transformed power values were computed for each electrode within each frequency band, yielding robust features for subsequent statistical testing.
Statistical Analysis
Statistical comparisons between the RS and RSM were conducted using a robust Yuen’s paired t-test, implemented in Python. For each frequency band, electrode-wise trimmed mean differences were computed, and t-statistics were calculated on 20% trimmed samples to enhance robustness against outliers. Cohen’s d was calculated as a measure of effect size. To control for multiple comparisons across 64 electrodes, false discovery rate (FDR) correction was applied using the Benjamini-Hochberg procedure (α < 0.05). Electrodes showing significant differences after FDR correction were visualised using topographic plots generated with the MNE-Python library.
Results
Frequency-specific Power Modulation Across Cortical Regions
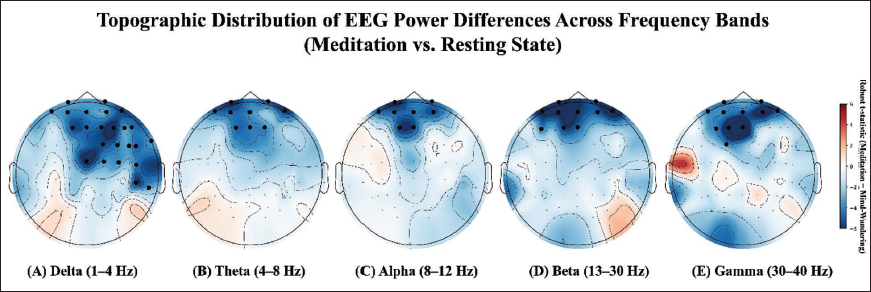
RSM significantly reduced EEG power across all frequency bands, primarily in prefrontal and frontocentral regions, compared to the RS condition (Figure 2). Significant power reductions were observed across delta, theta, alpha, beta, and gamma bands, with effects most pronounced in frontal and frontocentral cortices.
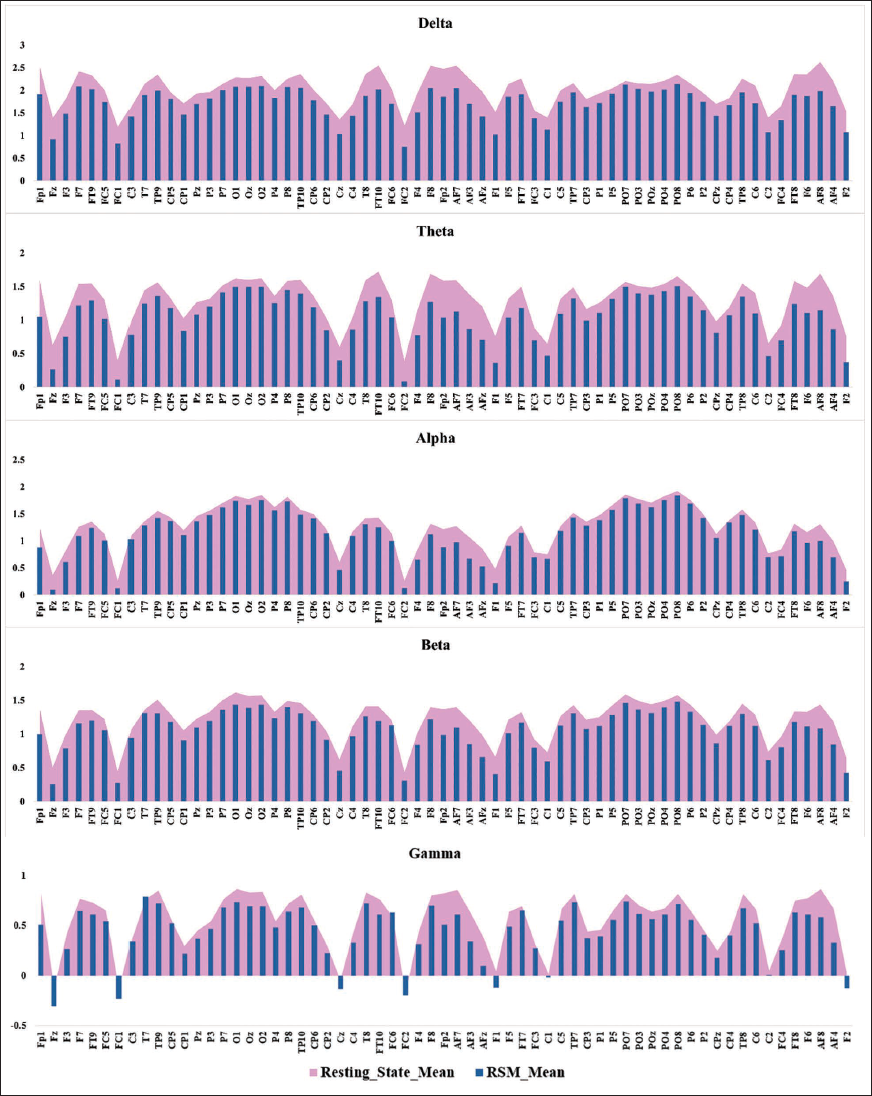
In the delta band (1–4 Hz), RSM led to widespread power suppression, particularly in prefrontal (Fp1, Fp2, Fz, AF3, AF7, AFz), frontocentral (FC2, FC4, FC6, Cz, C2, C4), and temporoparietal (TP8, TP10, T8, FT8, FT10) regions. Posterior and parietal areas showed negligible changes. Theta band (4–8 Hz) power was significantly lower during RSM in anterior prefrontal electrodes (Fp1, Fp2, Fz, AF3, AF7, AFz, F1, F2, AF8). Alpha band (8–12 Hz) power decreased in prefrontal regions (Fp1, Fp2, Fz, AF3, AF7, AFz, F1, AF4). Beta band (13–30 Hz) reductions were localised to prefrontal electrodes (Fp1, Fp2, Fz, F1, F5, AF3, AF7, AFz, AF8, AF4). Gamma band (30–40 Hz) power also decreased significantly in prefrontal and frontocentral areas (Fp1, Fp2, Fz, F3, FC1, AF3, AF7, AFz, F1, AF8, AF4). All comparisons used Yuen’s paired t-test with 20% trimming, with significance determined after FDR correction (p < .05). Figure 3 illustrates the topographical distribution of these power differences across RS and RSM sessions. Detailed statistical results for PSD across all channels and five frequency bands are provided in Supplementary Table S1.
Subjective Alertness Based on the SSS
Analysis of subjective alertness using the SSS revealed distinct changes between conditions. Prior to the RS, 93.3% of participants reported feeling alert, indicated by SSS scores of 1–2. This proportion declined to 73.3% following the session, reflecting a reduction in wakefulness. However, all the subjects reported that they were not sleepy during the RS. In contrast, prior to RSM, 86.7% of participants were alert, increasing to 93.3% post-meditation. Figure 4 presents these changes in self-reported alertness.
Discussion
Employing a within-subject approach to compare RSM with the RS condition, we observed consistent and frequency-specific decreases in EEG power throughout delta, theta, alpha, beta, and gamma bands during meditation, especially in the prefrontal, frontocentral, and fronto-lateral regions of the brain. Additionally, the increase in subjective alertness after meditation and its decrease following the RS condition suggest that RSM may foster altered arousal states.
While many previous EEG studies on meditation reported an increase in theta and alpha wave activity, particularly during unstructured or open monitoring meditation,8, 9, 12, 31 more recent research demonstrates heterogeneous patterns depending on meditation type and practitioner experience. Notably, sound-based practices show variable findings: some studies report increased theta and alpha power following AUM chanting.22, 31 While others document power decreases during meditation states characterised by ‘mental emptiness’ or focused attention.13, 14 Our observed reductions across delta, theta, alpha, beta, and gamma bands, particularly in prefrontal and frontocentral regions, align with the latter pattern and extend these findings to RSM in meditation-naïve participants. The functional significance of EEG power suppression during meditation admits multiple interpretations. Most parsimoniously, our pattern suggests active attentional engagement rather than cortical hypoactivity, supported by three converging lines of evidence: (a) the concurrent increase in subjective alertness, distinguishing our findings from drowsiness-related power reductions 36 ; (b) the frontal topographic emphasis, implicating executive control regions rather than global deactivation; and (c) consistency with task-related power decreases reflecting neural efficiency and reduced DMN activity.17, 37 The widespread power reduction may reflect enhanced neural efficiency through reduced synchronised population activity and decreased self-referential processing, consistent with meditation’s effects on reducing mind-wandering. 6 Nevertheless, we acknowledge that power suppression has been associated with both enhanced task engagement and reduced cortical activation in different contexts, and definitive mechanistic conclusions require complementary methods such as fMRI for anatomical localisation and connectivity analysis for network-level characterisation.
The lowering of delta activity during meditation is consistent with previous research, which reported that delta oscillations are primarily observed during sleep, deep relaxation, and may also decline during heightened alertness. 38 While some studies are associated with increased delta activity to mind-wandering at a trait level,39, 40 our observation of a notable lowering of delta power during meditation indicates a state of increased alertness and reduced cortical idling. Our findings also support the lower delta activity during eyes-closed transcendental meditation. 41 Furthermore, delta power was suppressed during short mindfulness meditation sessions in young adults 42 and in the experienced breath-focused meditators, 43 suggesting less cortical idling and greater neural efficiency. Collectively, these findings support the idea that a reduction in delta power could serve as a consistent electrophysiological marker of heightened meditative engagement and extended internal focus. This interpretation is further supported by the concurrent increase in subjective alertness in our sample, distinguishing meditation-related power suppression from hypoarousal states. Likewise, theta power is attributed to internal focus, emotional processing, and the depth of meditation.44, 45 In contrast to the increased theta activity observed during passive or emotional practices, the concentrated auditory focus required in RSM may lower frontal-midline theta, potentially reflecting altered cognitive processing. This perspective aligns with findings reported a decline in frontal theta and overall EEG power during states characterised by meditative emptiness, and during silent trance-like absorption, compared to mind-wandering.13, 30 These observations imply that states of focused auditory or introspective meditation may inhibit frontal-midline theta and thus may reflect changes in attentional processing. However, alpha suppression during meditation, especially in the prefrontal cortex, is increasingly interpreted as a marker of reduced DMN activity and enhanced attentional control.17, 36, 46 While relaxation-based and open monitoring meditation typically shows alpha enhancement,31, 37 and some sound-based practices report alpha increases post-chanting, 31 our findings of reduced alpha amplitude align with studies on focused attention and mindfulness meditation in novices.14, 29 However, the reductions in alpha amplitude during mindfulness meditation can be interpreted as an indication of reduced mind-wandering and are consistent with altered attentional states. This pattern suggests reduced mind-wandering and enhanced attentional focus, potentially reflecting the structured, attention-demanding nature of RSM’s sequential stages compared to passive chanting alone. Furthermore, beta activity indicates engagement in cognitive tasks, planning motor actions, and processing sensory information. 47 In the meditation context, a reduction in beta power is associated with a state of mental stillness and sensory detachment. Our findings showed diminished beta power across prefrontal and frontocentral electrodes, aligning with previous studies that observed beta suppression during meditation, attributed to lower cognitive effort and reduced demands for integrating information.15, 48, 49 Moreover, gamma rhythms are frequently associated with complex integrative functions and conscious awareness. 50 Though increased gamma activity has been demonstrated in experienced meditators engaged in compassion-based meditative techniques, 51 and subsequently other studies have also observed a decrease in gamma during concentrated, mantra-oriented meditation.15, 52 Our results are in line with this decreased gamma activity during focused auditory meditation. While we interpret this as reflecting reduced cognitive elaboration, we acknowledge that without source localisation or connectivity analysis, definitive conclusions about the underlying neural mechanisms remain tentative. 53
The overall decrease observed in all significant EEG frequency bands, especially in the prefrontal, frontocentral, fronto-lateral, and central areas, is suggestive of a widespread lowering of neural oscillations related to the DMN. These findings are consistent with neuroimaging studies showing that meditative states correspond to altered DMN activity,6, 50 though our EEG findings cannot directly confirm DMN involvement without source localisation analysis.
The increase in alertness reported by participants after RSM, contrasted with decreased alertness following RS, provides important behavioural context for interpreting the observed EEG power reductions. This divergence between reduced oscillatory power and enhanced subjective alertness supports the interpretation that power suppression reflects active attentional engagement rather than drowsiness or cortical hypoactivity. 54 The alignment between EEG suppression patterns and heightened wakefulness suggests that RSM may promote sustained alertness through reduced self-referential processing and enhanced attentional control. However, the reliance on subjective self-report (SSS) without objective behavioural measures limits strong conclusions about functional cognitive benefits, highlighting the need for future studies incorporating performance-based vigilance tasks.
Conceptual Framework: Potential Neural Mechanisms of RSM
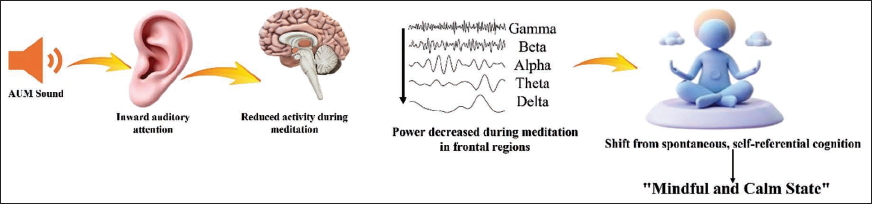
Figure 5 illustrates a conceptual framework for potential mechanisms by which RSM might influence brain function. This hypothetical model integrates our observed scalp-level EEG findings (widespread power reduction across frequency bands, particularly in frontal regions) with theoretical constructs from the broader meditation neuroscience literature. We propose that systematic auditory focus on AUM, gradually transitioning to internal silence, may stabilise attention and reduce self-referential processing, as potentially reflected in the observed decreases in neural oscillations across delta, theta, alpha, beta, and gamma bands. Important caveats apply to this conceptual model. First, our scalp-level EEG recordings cannot definitively localise these effects to specific deep brain structures or networks. While we hypothesise potential involvement of DMN regions based on topographic patterns and convergent fMRI literature on meditation,6, 17 confirmation would require source localisation methods such as LORETA, beamforming, or simultaneous EEG-fMRI. Second, we did not measure functional or effective connectivity between brain regions, precluding conclusions about directional information flow or network-level interactions depicted by the arrows in Figure 5. The illustrated pathways represent hypothesised relationships requiring validation through connectivity analyses (e.g., Granger causality, dynamic causal modelling). Collectively, while the observed oscillatory patterns are consistent with a potential transition from self-referential cognitive processing to present-centred awareness, the model should be interpreted as hypothesis-generating rather than established causal mechanisms. Future research employing source reconstruction, connectivity analysis, and complementary neuroimaging modalities is essential to test the specific mechanistic pathways proposed in this framework.
Limitations and Future Directions
Several limitations qualify the interpretation of these findings. First, with N = 15 participants, this exploratory pilot investigation has limited statistical power and generalizability. The within-subject design partially mitigates power concerns, but replication in larger samples is essential to establish reliability. Second, the study lacked an active control condition to isolate meditation-specific effects from nonspecific factors such as expectancy, placebo response, or general relaxation. Third, we relied on the SSS, a single-item subjective measure, without objective behavioural assessments of attention, vigilance, or cognitive performance, limiting conclusions about functional significance. Future research should incorporate performance-based measures (e.g., Psychomotor Vigilance Task, executive function tasks) to validate neurophysiological findings. Fourth, as an exploratory investigation with extensive statistical testing despite FDR correction, some effects may represent false positives requiring confirmation through preregistered, hypothesis-driven designs. Additionally, our scalp-level EEG recordings cannot definitively localise effects to specific brain networks without source reconstruction or connectivity analysis. Interpretations regarding DMN involvement remain speculative. Despite these limitations, this pilot study provides preliminary evidence that RSM produces distinct neurophysiological signatures in meditation-naïve participants, establishing feasibility and generating hypotheses for future adequately powered, rigorously-controlled confirmatory investigations.
Conclusion
This exploratory pilot study found that RSM produced widespread reductions in delta, theta, alpha, beta, and gamma power, predominantly in frontal regions potentially associated with attention, cognitive control, and self-referential processing. Coupled with increased subjective alertness, these preliminary findings suggest that RSM may facilitate an alert yet calm meditative state characterised by reduced cortical oscillations and mental clarity. While replication in larger samples with rigorous controls is essential, this study provides initial neurophysiological evidence that sound-based meditation practices may engage distinct neural mechanisms compared to other meditation forms, contributing to our understanding of meditation’s heterogeneous effects on brain function and generating hypotheses for future confirmatory research.
Footnotes
Acknowledgements
We thank the volunteers who participated in this study for their time and commitment. We also appreciate the medical officer who performed participant screenings and the Central University of Rajasthan for providing access to CIL EEG facilities.
Authors’ Contribution
Km Megha: Conceptualisation, methodology, software, validation, formal analysis, investigation, writing original draft, writing - review & editing, visualisation, project administration. Ankita Mishra: Literature search and manuscript formatting. Raksha Sharma: Review of manuscript content, literature search and analysis of the table contents. Pallabi Pal: Literature search, manuscript formatting and analysis of the table contents. Vijay Shanker Yadav: Literature search and writing - review & editing. Arjun Ram Roj: Validation, resources, data curation, writing - review & editing, visualisation. Arun Sasidharan and Gulshan Kumar: Software, validation, resources, formal analysis, writing - review & editing, visualisation. Sanjib Patra: Conceptualisation, resources, writing - review & editing, supervision.
Consent for Publication
Not applicable.
Consent to Participate
Written informed consent was obtained from all the participants following a detailed explanation of the research objectives before their involvement in the study.
Data Availability
The datasets used and/or analysed during the current study are available from the corresponding author upon reasonable request. Detailed statistical results are provided in Supplementary Table S1.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
Origin of the Work
The work originated from the Neurophysiological Laboratory, Department of Yoga, Central University of Rajasthan, Ajmer, Rajasthan, India CIL EEG facilities.
Statement of Ethics
The study received approval from the Institutional Ethics Committee of CURAJ (CURAJ/H-IEC/24/12/008).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.