
Abstract
Background
Suicidal behaviour (SB) is one of the most complex neuropsychiatric conditions, with an etiopathogenesis that remains elusive despite a myriad of studies revealing its multifaceted nature, influenced by various biological, psychological, socioeconomic and cultural factors, usually with complex reciprocal and synergistic interactions. Emerging evidence has recently suggested a potential involvement of pathogens and host-tissue derived microbial species in contributing to SB. This review aims to provide a concise synthesis of clinical evidence focusing on the presence of microbial alterations in subjects with SB and in those exhibiting risk factors for SB, thereby exploring a possible new perspective in suicidology.
Summary
We conducted a surveillance of the literature in the PUBMED database, from its inception to 10 May 2025, to identify associations between infection/microbial alterations with SB and some of its risk factors (by focusing on two pathologic examples of psychiatric and somatic origins, respectively: depression and chronic pain, two often closely related conditions). Major clinical findings were selected and synthesised into a hypothetical framework to support the presence of a microbe-related origin of SB. We found that SB was associated with selected infections, such as Toxoplasma gondii and Cytomegalovirus. Dysbiosis, including changes in diversity and/or abundance of selected microbial species, in the oral cavity, gastrointestinal tract, and other mucosal tissues, was observed in subjects with SB and individuals with psychiatric (depression) and somatic (chronic pain) risk factors of SB.
Key Message
Microbial dysbiosis might contribute to SB etiopathogenesis. Further studies in this emerging field of research are expected to provide additional mechanistic insights for an improved understanding, prevention, and therapeutic management of this neuropsychiatric condition.
Keywords
Background
Suicidal behaviour (SB), which includes suicidal attempts (SA) and completed suicides, is one of the leading causes of death and disability worldwide, with substantial healthcare and socioeconomic burdens. Current treatments for SB primarily focus on pharmacological and psychotherapeutic interventions targeting neuropsychiatric symptoms. 1 Since prevention approaches still show limited efficacy, further research into the complex pathophysiology of SB is needed to facilitate advances in clinical care for affected individuals.
SB etiopathogenesis is a multifaceted phenomenon, influenced by various biological, psychological, socioeconomic, and cultural factors, usually with complex reciprocal and synergistic interactions. In particular, growing evidence suggests links between increased suicide risk and immunological inflammation, 2 neurometabolic dysregulation 3 and various neuropsychiatric conditions, 4 warranting additional research to further elucidate the mechanisms that contribute to these putative pathological pathways of SB. Additionally, recent discoveries have shown how microbial species can modulate and interact with the immune system, 5 metabolic tissues 6 and neurobiological processes 7 via different molecular and cellular pathways. These findings have ignited scientific and clinical interests in exploring the possible role of microbes in SB pathogenesis.
Many types of microorganisms have developed a symbiotic relationship with humans. These symbionts, including bacteria, viruses, fungi, and protozoa, reside on the mucosal surfaces of the human body, such as the skin, nails, hair, oral cavity, and gastrointestinal/respiratory tracts. Together, they constitute the host microbiome, normally present in an eubiotic state. If there is an imbalance in the relative abundance/composition of microbial species and/or changes in their distribution (quantitative/qualitative changes), the host microbiome is no longer in homeostasis, a condition termed dysbiosis.
Dysbiosis has been shown to contribute to the development of various somatic illnesses, 8 ranging from immune-mediated conditions, such as allergy and asthma, to metabolic disorders, such as gastrointestinal diseases and obesity. Furthermore, the recent discovery of a gut-brain connectome suggests that dysbiosis may be implicated in neurodegenerative and neuropsychiatric illnesses, including Alzheimer’s and Parkinson’s disease, depression, and potentially SB. 9
This critical review summarises major clinical evidence for the hypothesis that microbial alterations exist in subjects with SB and in those exhibiting risk factors for SB, by focusing on depression and chronic pain (two closely related conditions of psychiatric and somatic origins, respectively).
Methods
In this critical review, we selected representative clinical studies that could highlight the emerging importance of microbial alterations in the pathophysiology of SB. To achieve this goal, we conducted a surveillance of the literature related to the main topic of this review (i.e., the association between different microbes and SB) in the PUBMED database, using the following keywords: [‘microbe’ OR ‘microbiota’ OR ‘microbiome’ OR ‘dysbiosis’ OR ‘infection’] AND [‘suicidal behavior’ OR ‘suicide’ OR ‘suicidal attempt’ OR ‘suicidal ideation’ OR ‘chronic pain’ OR ‘depression’] from the inception of the database until 10 May 2025. Preclinical studies (those employing animal models of psychiatric illnesses) or those reported in languages other than English were excluded. The content of all studies with human subjects was screened, and major findings from relevant studies were categorically summarised in the sections below to highlight the potential association between infection/microbial dysbiosis with SB and its risk factors.
Infections and SB
Infection-induced neurochemical changes in the central nervous system and inflammatory responses that elevate suicide susceptibility may provide a potential mechanistic explanation for the association between infection and elevated suicide risk. 2 In this regard, one of the earliest indications of a possible association between microbes and SB was suggested by a correlation between increased suicide risk and Toxoplasma gondii infection in the general population across 20 European countries. 10 A follow-up study revealed that T. gondii seropositivity was significantly associated with increased suicide rates in women, particularly among those at 60–74 years of age. 11 In a different study of more than 45,000 participants, violent SA risk and self-directed violence risk among infected child-bearing women were, respectively, 1.81 and 1.53 fold higher than those of non-infected counterparts, with a positive correlation between self-directed violence risk and T. gondii IgG titres. 12 Seropositivity for T. gondii was also associated with suicidal ideation (SI) in women attending primary care clinics. 13 In this study, T. gondii IgG antibody detection rate was 11.9% in individuals with an SA history compared to a control rate of 8.0%. Furthermore, T. gondii seroprevalence (anti-T. gondii IgG titre >150 IU/mL) was associated with the presence and number (>3) of SA in this patient cohort. 13
In schizophrenia patients, T. gondii infection has also been linked to SA and SI. For example, T. gondii, but not cytomegalovirus (CMV) or herpes simplex virus 1, seropositivity was significantly associated with SB in a subset of schizophrenics who were younger than 38 years of age. 14 In another study that controlled for age and sex, T. gondii seroprevalence was significantly higher in schizophrenia patients with SA while T. gondii seropositivity was not significantly associated with SA. 15 Furthermore, T. gondii infection and plasma level of kynurenine, a metabolite previously implicated in the pathogenesis of SB, 16 were suggested to have cumulative effects on the risk of non-fatal suicidal self-directed violence among patients with schizophrenia. 17 Of note, among patients with T. gondii seropositivity, those with kynurenine levels of the upper 25th percentile showed a 1.95 adjusted odds ratio for the association between non-fatal suicide history and plasma kynurenine concentration. 17
In patients with depression, latent toxoplasmosis (IgG, but not IgM, seropositivity for T. gondii) was associated with SB. In this regard, participants with T. gondii IgG seropositivity showed a 2.9-fold increase in depression odds and a 6.2-fold increase in odds of having an SA history compared to seronegative counterparts. 18 In another study, T. gondii IgG titre was significantly higher in depressed subjects with SI. 19 Higher T. gondii IgM level and IgM seropositivity for T. gondii (an indicator of acute infection) were also observed in suicide attempters with active depressive symptoms. 20 In a post-mortem study of suicide victims, the presence of neuroinvasion by T. gondii (8% in a study of 87 subjects) was associated with a history of depression. 21 Lastly, higher toxoplasmosis titres were also suggested as an indicator for increased susceptibility to SA among older adolescents receiving antidepressant treatment with selective serotonin reuptake inhibitors. 22
Infectious agents other than T. gondii have also been implicated in the pathogenesis of SB. For example, CMV seropositivity was reportedly associated with SA in a large-scale study of subjects with various psychiatric illnesses (n > 2,500). 23 Similarly, CMV IgG levels were higher among depressed suicide attempters. 20 Interestingly, this study also revealed a similar association between CMV seropositivity and SA, which had an intriguing additive effect on the association of T. gondii with SA. Infection during the hospitalisation period was also linked to an increase in suicide risk in a cohort study of more than 7 million subjects. 24 Importantly, a dose-response relationship between the number of hospital-related infections and the length of infection-related treatment with suicide risk was observed, with an incidence-rate ratio of 2.9 and 2.38 for having more than 7 infections and having been hospitalised for infection-related treatment for more than 3 months, respectively. Most recently, reports of increased neuropsychiatric complications, including SB, have been described in SARS-CoV-2-infected patients. 25 In a study of 115 long COVID-19 patients from France, a significant increase in suicide prevalence was observed at 4 months after intensive-care unit admission. 26 In another study of 506 adults with a history of COVID-19 diagnosis from the United States, higher SI and SB scores were documented up to one year after infection. 27
It is worth noting that some contradictory findings emerged in other studies regarding the association between infections and elevated suicide risk. For example, a recent study demonstrated that rates of T. gondii seropositivity (including IgG) and seroprevalence were similar between controls and suicide attempters.28, 29 A lower rate of SA among schizophrenia patients with Toxoplasma IgG seropositivity as compared to seronegative counterparts was also reported. 30 Furthermore, a 15-year longitudinal study found no association between general infection status and the risk of suicidal death. 31 These discrepancies may be attributed to differences in sample sizes and patient selection criteria, as well as the geographical areas and detection methodologies used among the aforementioned studies. Nevertheless, two recent systematic analyses confirmed a potential association between T. gondii infection and SB.32, 33 Lastly, some studies suggested that anti-infectives could be associated with an elevated risk of suicide. For example, in a longitudinal study of more than 15,000 Danish participants with documented self-harm behaviours, anti-infective treatments were linked to an increased risk of self-harm with a hazard rate ratio of 1.8. 34 The use of the anti-malaria drug hydroxychloroquine/chloroquine has also been linked to increased risk of suicide, possibly via its cardiac and neuroendocrine toxicities. 35 A summary of key findings from the primary studies in this section is provided in Table 1.
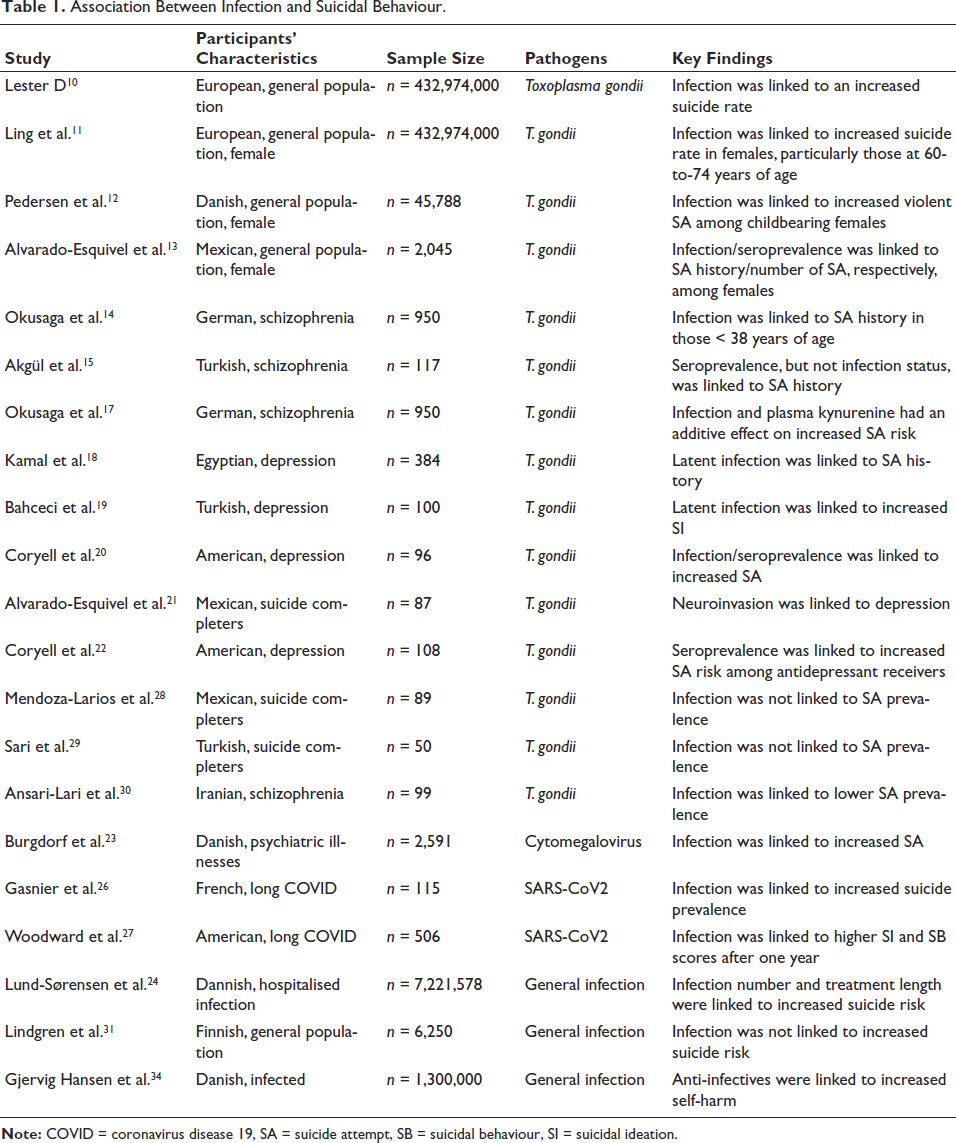
Association Between Infection and Suicidal Behaviour.
Microbial Dysbiosis in Subjects with SB
There is already some, yet limited, evidence in the literature regarding the presence of dysbiosis during SB development, which is primarily characterised by spatial alterations of microbiome in these subjects (Table 2). The following section will detail the changes detected in various microbiome compartments in subjects with SB.
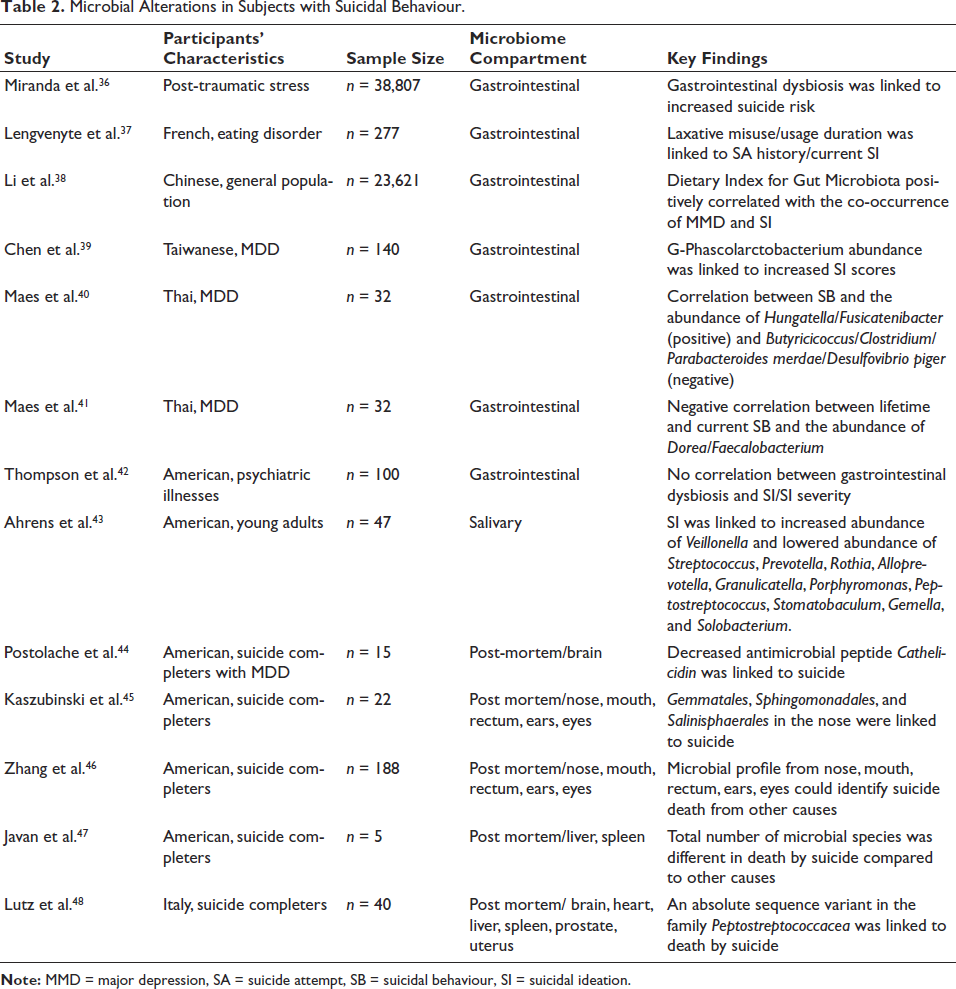
Microbial Alterations in Subjects with Suicidal Behaviour.
Gastrointestinal and oral Microbiomes
The gastrointestinal tract contains the largest number of commensals in the human body. Notably, gastrointestinal dysbiosis has recently been linked to the development of SB in specific vulnerable patient populations. For example, a case-control study examining the electronic medical records of over 30,000 patients with post-traumatic stress disorder found that changes in gastrointestinal bacteria were a significant predictor of the risk of suicide-related events, including both SB and completed suicides. 36 Similarly, in a cohort study of patients with eating disorders (n = 277), those with a history of SA exhibited a cumulative increase in laxative misuse, which had previously been linked to a disruption of gastrointestinal microflora. 37 Notably, duration of laxative usage in the past 4 weeks was also significantly associated with current SI in these subjects. 37
Besides these ‘proof of concept’ clinical studies, alterations in the gastrointestinal microbiome have been implicated in suicide development. In this regard, a study of more than 20,000 participants revealed a striking dose-response relationship between the Dietary Index for Gut Microbiota (a measurement of gastrointestinal microbial diversity) with SI in subjects experiencing major depression, 38 supporting the influence of gastrointestinal commensals on the evolution of SI. SI has also been linked to the alteration of specific microbial species in the gastrointestinal tract. In this regard, assessment of faecal microbiome in a study of 140 participants from Taiwan revealed a significant positive correlation between the Beck Suicidal Ideation Scale score with the abundance of g-Phascolarctobacterium, particularly among patients with depressive symptoms. 39 Similarly, unique gastrointestinal microbial profiles have been identified in subjects with SB. For example, in patients with major depressive disorder (n = 32), 6 microbial taxa were identified as a putative enterotype with clinical association with SB. 40 Specifically, Hungatella and Fusicatenibacter were positively associated with SB, while Butyricicoccus, Clostridium, Parabacteroides merdae, and Desulfovibrio piger showed an inverse association. Of note, adverse childhood experiences and the recurrence of depressive symptoms served as predictors of this microbial enterotype. Further, the prediction of current SB could be refined by the presence of this enterotype and of a high-density lipoprotein cholesterol level, the latter being predicted by a unique gastrointestinal microbial profile. In a follow-up study of the same research group, SB were confirmed to be related to a dysbiotic phenotype of the gastrointestinal microbiome among depressed participants. 41 Interestingly, lifetime and current SB showed a negative correlation with pathogenic Dorea and Faecalobacterium. Additionally, these SB features could be strongly predicted (63.5%) by the combinatorial usage of this microbial profile along with immunological (Th1 polarisation) and growth factor (increased vascular endothelial growth factor/stem cell growth factor ratio) alterations. It is also worth noting that no alteration in gastrointestinal microbiome was also reported in a study of 100 psychiatric patients, 42 which might be attributed to variations in demographic and environmental factors with a strong influence on gastrointestinal microbiome analysis.
To date, only one study has shown an association between SI and dysregulated microbial composition in the oral microbiome, the second largest microbial community in the body after the gastrointestinal tract. 43 Since the oral microbiome is influenced by sex, dietary, genetic, and sleep factors, propensity score matching was used to examine potential alterations in oral microbes in more than 30 university students with SI versus control subjects. Salivary microbial analysis showed that SI was associated with alterations in microbial abundance. Specifically, an increased abundance of Veillonella and a lowered abundance of Streptococcus, Prevotella, Rothia, Alloprevotella, Granulicatella, Porphyromonas, Peptostreptococcus, Stomatobaculum, Gemella, and Solobacterium was observed in subjects experiencing SI compared to controls. At the species level, Streptococcus oralis, Prevotella melaninogenica, Fusobacterium periodoncitum, Rothia mucilagniosa, Graulicatella adiacens, Streptococcus parasanguinis, Prevotella nanciensis, Solobacterium moorei, Dialister pneumosintes, Peptoanaerobacter stomatis, and Actinomyces naeslundii were more abundant in controls, while Bacteroides dorei and Fusobacterium naviforme were more abundant in subjects with SI. Furthermore, microbial diversity indices were reportedly reduced in subjects with the cooccurrence of SI and sleep disturbance.
Post-mortem Microbiome
While the previous sections discussed the impact of microbial alterations on living subjects with SB, some studies investigated the post-mortem microbiome in completed suicides. Among one of the first implications of microbial changes in post-mortem tissues of completed suicides was the observation that in the brain samples of depressed subjects who completed suicide, reduced level of the antimicrobial peptide cathelicidin (an indicator of changes in microbiome) was observed. 44 Subsequently, in an investigation of potential associations between the cause and manner of death with microbial communities from nose, mouth, rectum, ears, and eyes of more than 180 police records involving various causes of death, 45 three microbial orders, Gemmatales, Sphingomonadales, and Salinisphaerales, from the nose bacterial community were significantly associated with suicidal death. Interestingly, gun-shot suicides were distinguishable from homicides by 10 microbial orders, including Tepidisphaerales, Clostridiales, Rhodobacterales, Pirellulales, Corynebacteriales, Bacteroidales, Reyranellales, Cytophagales, Clostridiales, and Micrococcales. In support of these findings, another study demonstrated that the combinatorial use of post-mortem information from the five aforementioned microbial communities could predict the manner of death (accident vs. natural, homicide, or suicide) with >88% accuracy. 46 However, unlike the previous analysis, this study showed that death by suicide was associated with a higher abundance of Actinomyces species, likely due to potential changes in microbial communities during different time intervals after death. Besides the tissues examined in the studies above, the Chao1 index of microbial richness (the total number of species) in the liver and the spleen of suicide completers was reported to be significantly different from those of subjects who died by other causes. 47 In a more extensive investigation of microbial changes in the brain, heart, liver, spleen, prostate, uterus of suicide completers, death by suicide showed a significant inverse correlation with an absolute sequence variant in the family Peptostreptococcaceae. 48
Microbial Dysbiosis in Subjects with Chronic Pain and Depression
Modulating somatic and psychiatric risk factors of SB might represent a potential pathway by which host microbial alterations could contribute to SB development. This section provides an overview of how dysbiosis might contribute to SB development by focusing on chronic pain and depression, two representative somatic and psychiatric risk factors of SB, respectively.
Chronic Pain
Chronic pain has been defined as ‘an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage’. 49 Importantly, patients with chronic pain not only showed increased rates of SI, SA, and completed suicide 50 but also experienced various changes in their microbial repertoires. In this regard, some of the most notable observations of dysbiosis in subjects with chronic pain were reported in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS), an etiologically complex disease with a high prevalence of chronic visceral pain co-occurrence (>90%) 51 (Table 3). In ME/CFS, lower diversity of the gastrointestinal prokaryotic, but not eukaryotic, microbes was observed along with the expansion of inflammatory species and involution of anti-inflammatory species.52, 53 Specifically, increased gastrointestinal abundance of proinflammatory microbes, such as lactic-acid producing Enterococcus/Streptococcus spp, 54 Gram-positive anaerobic Lactonifactor and Alistipes, 55 and Gram-negative Bacteroides and Phascolarctobacterium, 56 were reported. Decreased abundance of several anti-inflammatory Firmicutes and Lachnospiraceae populations was also observed in ME/CFS patients from three independent studies.52, 55, 57 Furthermore, increased pain in ME/CFS patients was associated with decreased abundance of beneficial short-chain fatty acid (SCFA)-producing Faecalibacterium species. 58 Along with the implication of gastrointestinal dysbiosis in ME/CFS, one study also revealed an increased salivary abundance of Rothia dentocariosa, 56 a species that has been linked to inflammation and endocarditis. 59
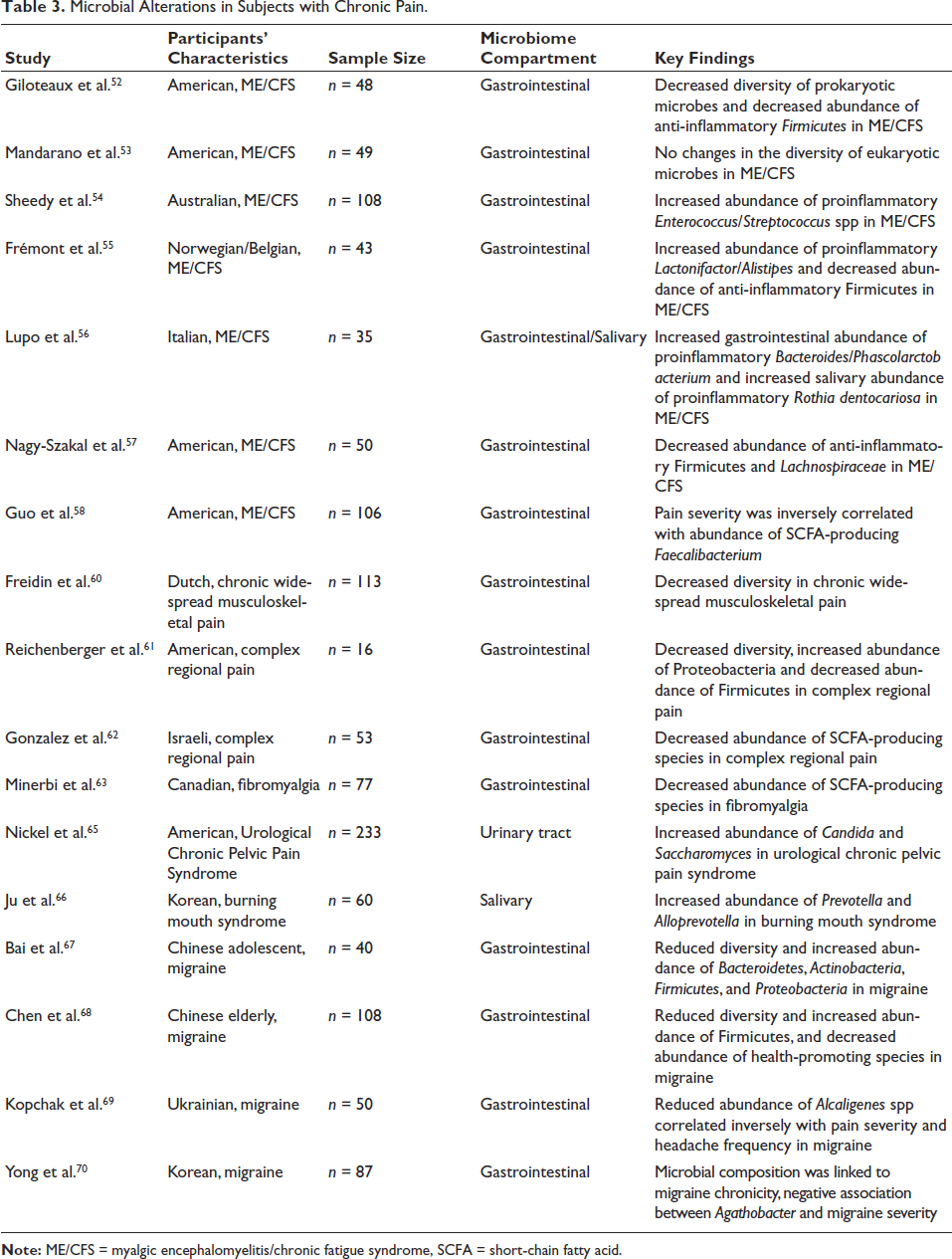
Microbial Alterations in Subjects with Chronic Pain.
Besides ME/CFS, dysbiosis has been observed in other diseases of chronic visceral pain. For example, patients with chronic widespread musculoskeletal pain and complex regional pain syndrome showed reduced gastrointestinal microbial diversity.60, 61 In the latter condition, increased gastrointestinal abundance of Proteobacteria and decreased abundance of Firmicutes and other SCFA-producing species were also observed.61, 62 Similarly, decreased gastrointestinal abundance of several SCFA-producing bacterial species, including Faecalibacterium prausnitzii, Blautia faecis, Haemophilus parainfluenza, Prevotella copri, and Bacteroides uniformis, was reported in patients with fibromyalgia. 63 This dysbiosis profile of fibromyalgia was further supported by urine metabolomic analysis of microbial metabolites. 64 Of note, changes in microbial compartments other than the gastrointestinal tract are present in diseases of chronic visceral pain. In this regard, a recent study of chronic pelvic pain revealed urinary-tract accumulation of the fungal species Candida and Saccharomyces. 65 In burning mouth syndrome, higher relative salivary abundance of bacterial species was observed compared to controls, with Prevotella and Alloprevotella as the most abundant genera in the patient group. 66
Similar to visceral chronic pain, recent findings have implicated a role of microbial dysbiosis in neuropathic chronic pain, such as migraine. In a study of adolescents with this chronic pain condition, reduced gastrointestinal microbial diversity and increased abundance of various phyla (Bacteroidetes, Actinobacteria, Firmicutes, and Proteobacteria) were noted. 67 Similarly, elderly subjects with migraine also exhibited reduced gastrointestinal microbial diversity, with increased abundance of Firmicutes and depletion of health-promoting species (such as Faecalibacterium prausnitzii, Bifidobacterium adolescentis, and Methanobrevibacter smithii). 68 Furthermore, gastrointestinal Alcaligenes spp was reportedly reduced in migraine patients compared to controls, and its abundance showed a negative correlation with Visual Analogue Scale pain score and headache frequency. 69 Interestingly, gastrointestinal microbial composition, but not diversity, was shown to be related to the chronicity of migraine, with a negative association between Agathobacter and migraine severity. 70 Collectively, these diverse alterations of microbial composition and abundance at different anatomical locations in patients with various chronic pain conditions might support the potential presence of tissue-specific roles of microbes in chronic pain.71, 72
Depression
Most studies suggesting an indirect connection between microbial alterations and SB risk have been conducted in subjects with depression. In this regard, depression-associated dysbiosis is characterised by spatial changes in various microbiome compartments in the human body, most frequently documented in the gastrointestinal microbiome. Comprehensive reviews and detailed descriptions of gastrointestinal dysbiosis in depression have been extensively described elsewhere73–75 and summarised in Table 4a of this review. In this regard, one of the largest recent systematic reviews (44 studies) showed that the majority of analyses carried out in these studies (68 out of 86 analyses) found no difference in alpha-diversity (i.e., overall diversity of the gastrointestinal microbiome within a sample) between patients with depression and controls, while most of the beta-diversity examinations (13 out of 15 analyses) documented dissimilarity in the gastrointestinal microbiome between the two aforementioned study groups. 75 In terms of microbial abundance, at the phylum level, the most consistent changes were increases in Proteobacteria (5 out of 6 analyses) and Fusobacteria (3 out of 4 analyses). At the family level, increases in Rikenellaceae (6 out of 7 analyses) and Enterobacteriaceae (4 out of 4 analyses) and a decrease in Ruminococcaceae (9 out of 11 analyses) were commonly observed. At the genus level, major alterations included increases in Eggerthella (7 out of 7 analyses), Alistipes (8 out of 10 analyses), Parabacteroides (9 out of 10 analyses), Enterococcus (3 out of 3 analyses), Lactobacillus (3 out of 4 analyses), Veillonella (4 out of 4 analyses), Flavonifractor (6 out of 6 analyses), and Streptococcus (6 out of 6 analyses) as well as a decrease in Faecalibacterium (11 out of 12 analyses). Besides this phylogenetic investigation, functional consequences of gastrointestinal dysbiosis have also been quantified (9 analyses). The most notable changes were enrichment in glycan metabolism/synthesis, lipopolysaccharide and protein synthesis, and transport/catabolism pathways, along with a reduction in cell motility/secretion and membrane transport in patients with depression as compared to controls.
Several emerging studies also indicated alterations in microbes from tissues other than the gastrointestinal tract in patients with depression (Table 4b). For example, in adolescents with depression, increased abundance, but not overall diversity, of the salivary microbiome was associated with higher depressive symptoms.76, 77 In contrast, reduced oral microbial diversity was observed in another study of treatment-naïve adolescents, 78 suggesting a possible influence of antidepressants on oral microbial profile. Furthermore, in these studies, increased abundance of selected bacterial populations (including the phylum Spirochaetes and its order Spirochaetales; and the genus Neisseria and Prevotella nigrescens)76, 77 as well as decreased abundance of various other taxa (including the genera Prevotella, Rothia, Treponema, Schaalia, Neisseria, Solobacterium, Lepotrichia, Fusobacterium, Veillonella, Streptococcus, and Hemophilus)77, 78 were observed in subjects with higher depressive symptoms.
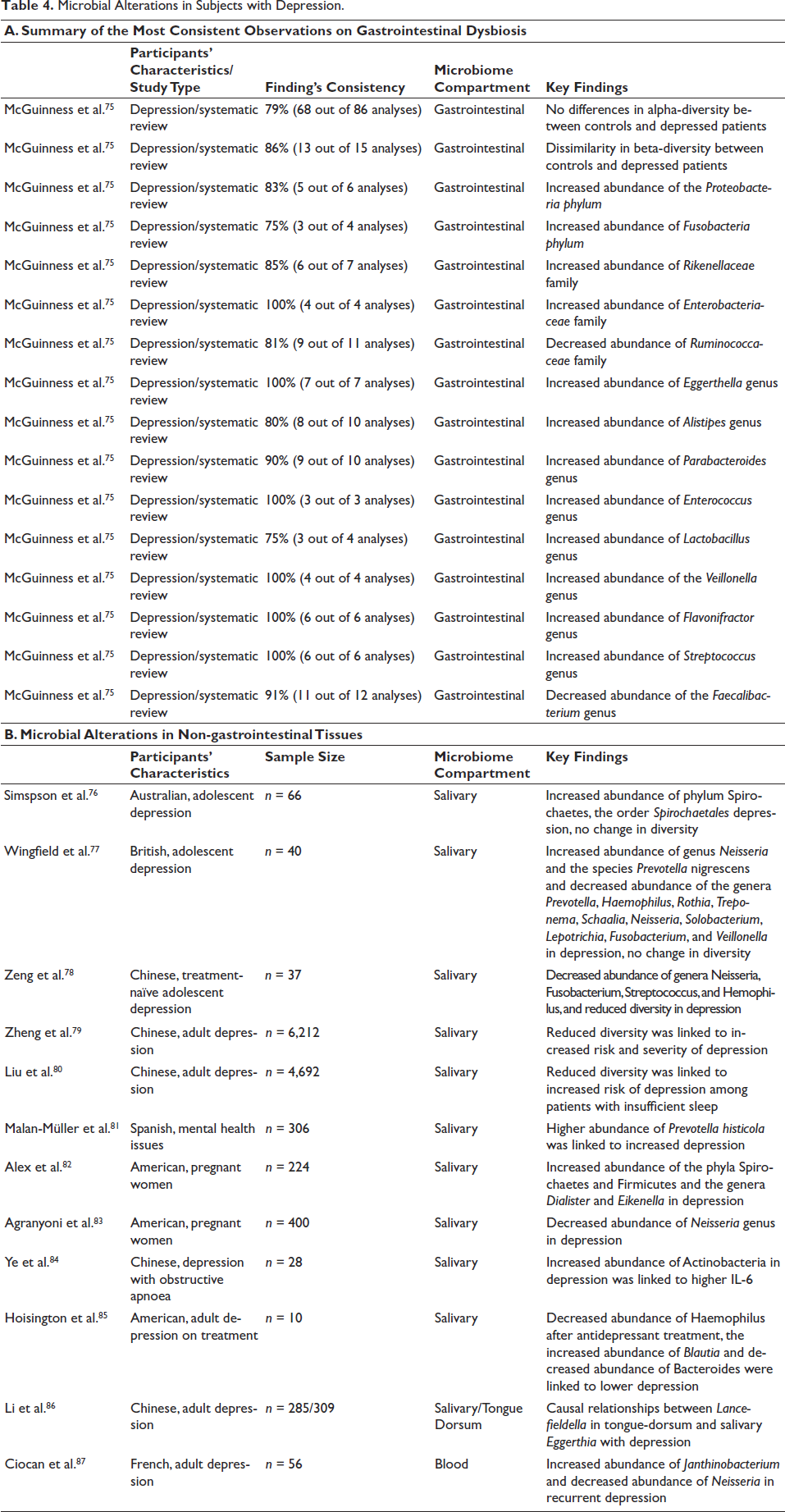
Microbial Alterations in Subjects with Depression.
In adults with depression, reduced oral microbial diversity has been linked to increased risk and severity of depressive symptoms, 79 particularly in those with insufficient sleep. 80 Regarding specific alterations of oral microbial populations, a positive association between oral abundance of Prevotella histicola and depressive scores was observed. 81 In pregnant women with depressive symptoms, lower oral abundance of several bacterial taxa, including the Neisseria genus, and higher oral abundance of the phyla Spirochaetes and Firmicutes, as well as the genera Dialister and Eikenella, have been reported.82, 83 In subjects with comorbid obstructive apnoea, increased salivary abundance of Actinobacteria was also observed, and might be modulated by higher circulating level of the inflammatory cytokine IL-6. 84 Lastly, antidepressant treatment was linked to a decrease in salivary abundance of Haemophilus, with additional associations between decreased depression scores and the abundance of Blautia (negative correlation) and Bacteroides (positive correlation). 85
Besides these association studies, a two-sample Mendelian randomisation analysis of salivary and tongue dorsum microbiomes (n > 280) suggested potential causal relationships between Lancefieldella in tongue-dorsum microbiome and Eggerthia in salivary microbiome with depression, with the latter as a potential common risk factor for depression and anxiety. 86 Lastly, changes in the blood microbiome have been reported recently in patients with recurrent depressive disorder (n = 56). 87 Compared to controls, patient blood samples exhibited higher and lower proportions of Janthinobacterium and Neisseria, respectively.
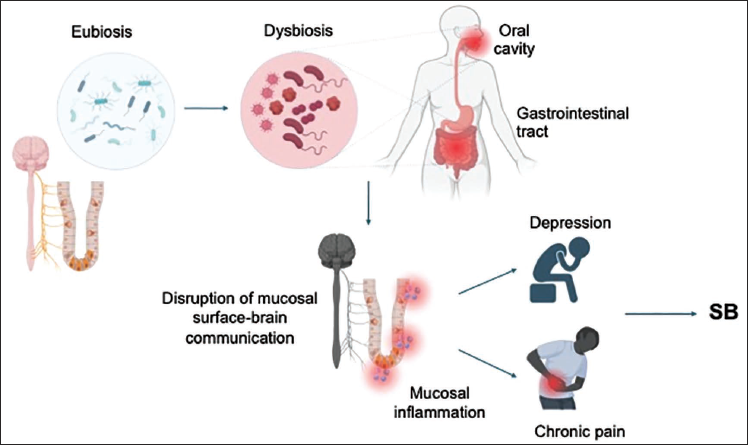
Combined with findings of dysbiosis in subjects with chronic pain, these observations of microbial alterations in depression suggest that dysregulation of multiple microbiome compartments might be intimately involved in the development of somatic and neuropsychiatric risk factors of SB and may predispose affected individuals to SB. Although the mechanisms by which microbes modulate the development of neuropsychiatric illnesses were not principally explored in these studies, the diverse findings on specific microbial profiles of subjects with chronic pain, depression and SB are consistent with the shift from health-promoting anti-inflammatory microbial species towards a pathogenic pro-inflammatory microbial state. We speculate that this spatial dysbiosis might result in compromised barrier integrity and subsequent dysregulated mucosal immunity and/or aberrant mucosal surface-brain communication, ultimately resulting in abnormal regulation of signalling cascades that have been implicated in SB development, including inflammatory and/or neurological pathways that promote selected somatic and psychiatric risk factors of SB (Figure 1).
Dysbiosis as a Putative Contributor to the Development of Suicidal Behaviour. Dysbiosis might contribute to suicidal behaviour by disrupting homeostatic interactions between the mucosal surface and other physiological systems (such as the immune and nervous systems), thereby augmenting various risk factors of this condition, including those of immunological (inflammation) and neurobiological (depression and chronic pain) origins.
Therapeutic Implications
Various therapeutic modalities, including prebiotics/probiotics (microorganisms that, upon consumption, restore beneficial microflora), microbe-associated metabolites, and microbial transplantation, have yielded promising clinical outcomes in the treatment of a wide range of human diseases. In this regard, a probiotic blend of B. lactis Bi-07, L. paracasei Lpc-37, Lactobacillus acidophilus NCFM, and B. lactis Bl-04 with omega-3 fatty acid supplementation could reduce circulating high-sensitivity C-reactive protein, a marker of systemic inflammation, in elderly participants. 88 Similarly, the combinatorial supplementation of Lactobacillus, Bifidobacterium, omega-3 fatty acid, and vitamin D resulted in a significant suppression of the proinflammatory cytokine IL-6 in obese subjects with chronic low-grade inflammation. 89 As such, targeting the inflammatory pathway, a biological risk factor of SB, by modifying the microbiome may represent a promising preventative approach.
Similarly, probiotic therapies and faecal transplantation were shown to have a positive impact on chronic pain in subjects with fibromyalgia.90, 91 Specifically, probiotic supplementation significantly decreased the Beck Depression and Anxiety Indices as well as the Pittsburgh Sleep Quality Index, while prebiotic supplementation only significantly improved the Pittsburgh Sleep Quality Index. 90 Several randomised trials also showed that microbiome-targeting therapies provide some psychological benefits for depressed patients.92, 93 For instance, a probiotic blend consisting of L. acidophilus, Lactobacillus casei, and Bifidobacterium bifidum provided a significant reduction in Beck Depression Inventory scores for clinically depressed patients. 94 Additionally, prebiotic intervention with 4G-beta-D-Galactosylsucrose has been reported to improve microbiome diversity and the Global Self-Efficacy Scale scores for patients with depressive symptoms. 95 Therefore, modulating chronic pain and/or depressive symptoms with microbiome-targeting therapies may provide additional preventative benefits to reduce SB risk.
Limitations
A limitation of this review includes the absence of a systematic approach to assess the quality, strength, or consistency of the findings related to SB and its risk factors, owing to the scarcity of studies in this emerging field of research as well as the lack of standardisation methods/difficulties in evaluating human microbiome in subjects with many SB-related conditions. We anticipate that with further efforts invested in this scientific topic of important public health concern, a more comprehensive systematic review will materialise in the near future. Additionally, while implying that microbe-associated inflammation might contribute to SB development, this review did not provide a detailed proposal on the roles of specific microbial populations in the development of SB. We anticipate that future development of methods to delineate the impact of specific microbes on various physiological processes of the host will clarify their role in SB pathobiology.
Conclusions
Dysbiosis in different compartments of the microbiome may indicate a developing pathology in subjects with SB and in individuals with various risk factors for this condition, such as chronic pain and depression. This concise collection of clinically relevant findings aims to serve as a basis for further research into how microbes modulate the biological and psychological processes involved in SB pathogenesis. By gaining new mechanistic insights into the potential dysregulation of crosstalk between microbes and SB-related pathological pathways, we may be able to develop more effective preventive and treatment approaches for this condition.
Footnotes
Abbreviations
CMV: Cytomegalovirus
COVID: Coronavirus Disease 19
ME/CFS: myalgic encephalomyelitis/chronic fatigue syndrome
MMD: Major Depressive Disorder
SA: Suicidal Attempts
SB: Suicidal behaviour
SCFA: short-chain fatty acid
SI: Suicidal Ideation
Acknowledgements
We would like to express our sincere gratitude to Dr Vasileios Chytas, from the Psychiatric Liaison Department of the Geneva University Hospital (HUG), for his insightful contribution to the revision of this manuscript.
Authors Contribution
Conceptualization: KDN, AC, LM.
Writing—original draft: AC, JA, LM, DS, KDN.
Writing—review & editing: AAm, AAg, AP.
Supervision: GS, MA.
Availability of Data and Materials
Not applicable.
Consent for Publication
Not applicable.
Statement of Ethics
Not applicable.
Declaration of Conflicting Interests
KDN is the scientific founder of Tranquis Therapeutics, a biotechnology company that develops novel treatments for neuro-inflammatory and neurodegenerative diseases. KDN is also a scientific advisor for Tochikunda, a biotechnology company that develops SARS-CoV-2 diagnostic devices. All other authors declare no conflict of interest and that they have no commercial associations (e.g., consultancies, stock ownership, equity interest, patent/licensing arrangement, etc.) that might pose a conflict of interest in connection with this work.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.