
Abstract
Background:
Visual short-term memory (VSTM) and attention were found to modulate neural activity predominantly in a superior parietal lobule. This is thought to be the selective attention importance for encoding and manipulation in VSTM. The major area of investigation mainly rested with the differences in the neural substrates and networks mediating these cognitive processes in near and far cortical structures.
Summary:
Based on previous investigations, the dynamic temporal window route of attention and time locked associated cognitive processes and sub-processes are sketched and its implication in VSTM study is discussed. Imaging cortical structures to isolate closely linked cognitive tasks require circumscribing to certain time-windows in which the paradigm should support to tap time-locked associated processes and sub-processes.
Key Messages:
The neural activities in intraparietal sulcus area 1–2 and angular gyrus during VSTM encoding are beyond the modulatory effects of selective and sustained attention.
The human posterior parietal cortex (PPC) is known to activate during diverse range of cognitive tasks. Among which, the involvement of the PPC in visual attention and working memory remains a hot spot in cognitive neuroscience. Consistent neuroimaging evidence indicates that dorsal parts of the PPC, particularly the superior intraparietal sulcus (IPS), encodes visual attention and subsequent modulation of visual short-term memory (VSTM) processes.1–3 Using multivoxel pattern analysis, human retinotopic areas support active maintenance during working memory encoding.4–6 If a task requires a top-down attention maintenance, selective attention and memory are strongly linked and such links are less obvious when a singleton target, by reducing attention control demand, is presented.7, 8 This suggests that selective modulation of attention needs reorientation at the target phase, which may involve VSTM maintenance. A major challenge in this field is dissociating the spatial and temporal characteristics of these two differing, yet interwoven, cognitive processes and further clarifying how these spatial and temporal differences are accounted for the difference in neuroimaging data. A recent functional magnetic resonance imaging study undertaken by Sheremata et al. 9 added a compliment and extended the evidence that human PPC (retinotopically defined IPS1-2) encodes VSTM and its associated content is dissociable from visuospatial attention related subprocesses.
Briefly, a summary of the work of Sheremata et al. 9 is provided before proceeding to review core properties of visuo-spatial attention, given that active maintenance of VSTM relates to attentional selection. On this account, studies have shown that attentional mechanisms are involved in updating cue dependent trial-by-trial rule-sets. 10 Then previous behavioral and neuroimaging studies aimed at tapping temporal and spatial characteristics of attention and VSTM and how the cognitive task used by Sheremata et al. 9 elucidated the two basic cognitive processes served by the PPC is highlighted. Following this, different views in the theoretical/conceptual underpinning of both attention and VSTM by offering an alternative explanation about how the cognitive demand of the task lead to different neuroimaging results is examined. By converging evidences, the study of Sheremata et al. 9 in tackling the different neural processes associated with attention and VSTM and the implication of this study are discussed.
Sheremata et al. 9 investigated retinotopically defined subcortical structures of PPC and early visual areas by scrutinizing blood oxygen level dependent (BOLD) signal changes, functional connectivity, and hemispheric asymmetries within PPC to identify if memory-specific task demand exists beyond visuospatial attention confounds. Sheremata et al. 9 claim that previous tasks used to study VSTM may contain subprocesses of top-down attention control, like sustained and selective attention, and posed the question whether stimulus-specific delay period, which requires working memory, is dissociable from the ability to sustain attention to an object in space. Their paradigm includes two visually identical tasks with differing rules. By controlling and chasing out sustained attention-related confounds, the authors are able to tap distinctive processes associated to VSTM. Manipulating the time window was the key element in their paradigm, for the time course of VSTM and visual attention, it is distinctively different. A spatial cue was first displayed for 500 ms followed by a series of rapid stimulus presentations, 150 ms each with 150 ms interval, following which a response cue was then presented for a fixed duration of 750 ms. To isolate sustained attention, spatial and response cues capping either end of the task are ignored and attention directed at the orientation of the target shapes within the stimulus presentations. Participants were asked to identify the number of stimuli presentations in which all target shapes are vertically aligned. In a second task designed to induce VSTM mechanisms, participants were asked to determine if the orientation of shapes in the response cue is congruent with the initial “sample” cue. The short-term memory task requires maintenance of visual information over time after the masked delay to compare with a response cue 1500 ms later. On the other hand, the attention task required rapid decoding of object orientation within stimulus presentations. By controlling the possible variance of substructures of the IPS, hemisphere, and the visual field of the stimuli, the authors demonstrated that VSTM maintenance in IPS1-2 is beyond attentional confounds and task difficulty. 9 Functional connectivity analysis of retinotopically defined that the IPS with other dorsal parts of the attention network extend the evidence of modulatory interactions between anterior parts of parietal cortex with task positive and negative networks11, 12 during VSTM tasks. These findings provide novel insight in two ways: VSTM was not confounded with attentional modulation and the network-based connectivity analysis revealed the possible interactive mechanisms within dorsal attention network and modulatory effects of selective attention during VSTM encoding.
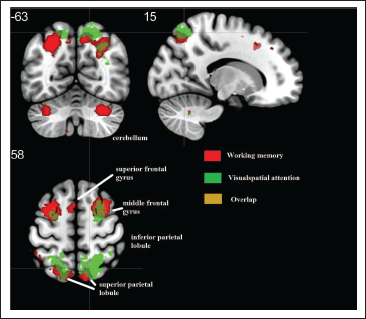
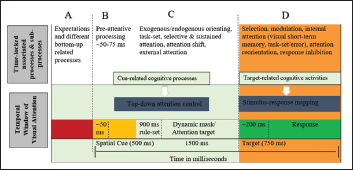
Previous studies examining the behavioral and neural basis of attention and VSTM report a complicated presence of attention that could be directed to specific items encoded in VSTM 13 and/or cognitive tasks with multiple distractors may be confounded with reorienting attention. 14 Top-down attentional modulation with multiple distractors may call-up for resource demand from VSTM14, 15 and response inhibition, which is considered to be part of attention, was argued to be updated from working memory resources. 16 Other behavioral evidences in VSTM show that orienting attention influences internal representations of the encoding process.10, 17 Taken together, these evidences suggest that the late stage of attentional modulation might be time-locked with the early encoding processes of VSTM (see Figure 2, column “D”). Kuo et al. 18 demonstrated that attentional mechanisms that works based on top-down attention processing during task encoding among other competing items are maintained by PPC (particularly posterior IPS) while modulating VSTM. This suggests that the posterior IPS may operate both attention and VSTM flexibly in an interactive and translative manner as suggested by Awh and Jonides. 14 Previous studies lack demonstrating if there is a clear distinction between these two cognitive processes and the role of PPC. PPC has shown an extensive overlap for short-term memory and visuospatial attention tasks (Figure 1). By controlling cue-related attentional processes (indicated in column “C” of Figure 2), Sheremata e al. 9 showed that activities in IPS 1–2 and angular gyrus are distinctive to VSTM encoding.
A Metanalysis of Functional Magnetic Resonance Imaging (fMRI) Studies Using an Online Neuroimaging Database (www.neurosynth.org ) That Involved Visuospatial Attention (Green) and Working Memory (Red) Task. The Overlapping Cortical Structures, Mainly in PPC and Middle Frontal Gyrus (MFG), are Shown in Orange.
The Temporal Window of Visual Attention and Time-Locked Associated Cognitive Processes and Subprocesses. Column A: Prestimulus Bottom-Up Processes, Expectations to the Upcoming Stimulus. Column B: The Visual Cue has Appeared but Task rules are Not Yet Set and is Related to Preattentive Processing (Ward, Duncan, & Shapiro, 1996). Column C: Task Rules are Set and Basic Task Related Processes Started Which is Associated with Multiple Cognitive Sub-Processes and Referred to as Top-Down Attention Control. Column D: The Target Appeared, Initiating the Processes of Stimulus-Response Mapping. Because of the Presence of a Distractor and Top-Down Attention Demand, This Sub-Process Might be Important Because of Links Between VSTM and Visual Attention During Stimulus-Response Mapping.
Even though behavioral experiments have demonstrated that attention task with distractors requires more top-down attention control than with singleton target, some behavioral results from VSTM are rather puzzling. In the VSTM decoding task there was no significant difference in the accuracy rate between target with and without distractors,19, 20 suggesting that VSTM and sustained attention operate relatively different. This might suggest that neural regions maintaining attention are more sensitive to time than VSTM encoding process.
In summary, the evidence from behavioral and neuroimaging studies provided two key insights regarding the “distinctive” process and neural mechanisms of attention and VSTM in that (a) imaging cortical structures to isolate closely linked cognitive tasks require to circumscribe to certain time-windows in which the paradigm should support to tap time-locked associated processes and subprocesses. In this regard, the authors present a clear distinction of VSTM to the remnants of attentional components as a confounder; (b) ensuring the BOLD signal differences in attention and VSTM are not associated with cognitive demand of the task served as a milestone to disentangle the distinctive neural processes of PPC during VSTM encoding.
In the study by Sheremata et al., 9 only cue-related attentional confounds are controlled. Examining stimulus–response mapping processes (as indicated in column “D” of Figure 2) with the same task may add another dimension in attention and VSTM. Trial-by-trial based multivoxel pattern analysis in retinotopically defined that parts of PPC may help to increase control over spatial attention confounds during VSTM encoding and the involvement of VSTM during top-down attention control process and target-related spatial updating as a confound. The latter is particularity appealing to scrutinize as neural processes related to reorienting attention when competing priorities presented at the target phase may require resource allocation from VSTM. By modeling target onset of the BOLD signal, scrutinizing the possible variance of substructures of IPS, hemisphere, and visual field of the stimuli would help to further isolate attentional confounds in VSTM.
Footnotes
Acknowledgments
We acknowledge Professor Chetwyn Chan and lab members of Applied Cognitive Neuroscience Laboratory, Department of Rehabilitation Sciences, The Hong Kong Polytechnic University, Hong Kong for their constructive comments.
Authors’ Contribution
AYD conceptualized drafted and written the manuscript and MAD reviewed and commented on the drafted manuscript.
Statement of Ethics
Not applicable.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.