
Abstract
Background: Exposure to adverse environmental conditions such as toxic chemicals, viral infections, and even stress during pregnancy or early life may disrupt the development of normal brain and its functioning leading to incidence of neurodevelopmental disorders at later stages of life. Recently, we reported that poly (I:C) exposure altered synaptic plasticity protein level and impaired memory through activation of microglia cells.
Purpose: As epigenetic modifications are involved in memory formation, we have studied methylation of DNA and acetylation of histone at promoters of synaptic plasticity genes in the brain of rats exposed to poly (I:C) during early life.
Methods: One dose of poly (I:C) (5 mg/kg bw) was intraperitoneally injected to rat pups on postnatal seventh day. A set of pups exposed to vehicle was included as control. In order to assess methylation of DNA and acetylation of histone at synaptic plasticity gene promoter, we performed qPCR after methylated DNA immunoprecipitation and chromatin immunoprecipitation.
Results: Poly (I:C) exposure reduced the level of 5-methylcytosine (5mC) at synaptic plasticity gene (bdnf, arc, and egr1) promoters in the frontal cortex (FC) and hippocampus of 3-week rats, although increased it later in both regions of 12-week rats as compared to respective controls. On contrary, poly (I:C) exposure enhanced acetylation of histone H3K9 (H3K9Ac) at promoters of these genes in both regions of 3-week rats but decreased in 12-week rats.
Conclusion: Poly (I:C) exposure altered 5mC and H3K9Ac at synaptic plasticity gene promoters resulting in memory impairment of rats at later life.
Keywords
Abbreviation
AD, Alzheimer’s disease; ARC, activity-regulated cytoskeleton-associated protein; BDNF, brain derived neurotrophic factor; ChIP, chromatin immunoprecipitation; DNMTs, DNA methyltransferases; EGR1, early growth response 1; FC, frontal cortex; HATs, histone acetyl transferases; HDACs, histone deacetylases; HP, hippocampus 5-mC 5-methyl cytosine; MeDIP, methylated DNA immunoprecipitation; NARP, neuro activity regulated pentaxin; PND, postnatal day; Poly (I:C), polyinosinic-polycytidylic acid; qPCR, quantitative polymerase chain reaction
Introduction
Brain development is a tightly regulated programmed process, which includes cellular events like proliferation, migration, differentiation, and synapse formation. 1 Prenatal exposure to adverse environmental conditions like toxic chemicals, viral infections, and even maternal stress during pregnancy negatively affects brain development and its function leading to neurodevelopmental disorders such as autism and schizophrenia in late life. 2 Our previous study showed impairment of learning and memory through activation of microglia and alteration in the level of expression of synaptic plasticity related proteins (BDNF, Arc, EGR1) in the frontal cortex (FC) and hippocampus (HP) of rats exposed to poly (I:C). 3 Highly activated microglia produce inflammatory cytokines and induce neuronal apoptosis. 4 In addition, poly (I:C) exposure triggers the apoptosis signaling leading to degeneration of neurons and subsequently deficits in cognitive function including memory. 5 On the other hand, it is reported that viral infection alters 5mC and histone acetylation, which in turn affects gene expression leading to neurodevelopmental disorders.6–9 Singh and Thakur 10 have also shown that variation in epigenetic modifications changes the synaptic plasticity gene expression in cerebral cortex as well as HP of aged mice.
Methylation of DNA and acetylation of histone play a major role in regulation of chromatin remodeling, gene expression, and memory formation.11–13 They have also important functions in normal aging and associated neurodegenerative diseases. 14 For DNA methylation, DNA methyltransferases (DNMTs) transfer methyl groups to fifth carbon of cytosine at CpG sites. In case of histone acetylation, histone transferases (HATs) transfer acetyl groups to histone tails and histone deacetylases (HDACs) remove them. 15 These epigenetic modification machineries are sensitive to environmental stimulus. 16 Earlier reports have shown that poly (I:C) alters methylation of DNA and acetylation of histones in brain as well as immune cells.17, 18
These evidences advocate that poly (I:C) exposure during early life might alter the expression of synaptic genes through modulation of epigenetic modifications and lead to memory deficits. This has prompted us to analyze methylation of DNA and acetylation of histone at synaptic plasticity gene promoters in the FC as well as HP of 3-, 6-, and 12-week rats.
Methods
Animals and Poly (I:C) Injection
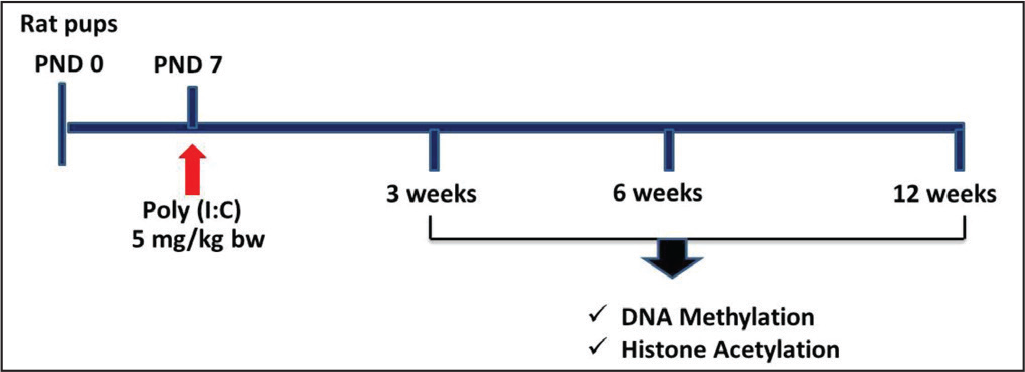
Wistar rats were maintained in animal colony at School of Studies in Neuroscience, Jiwaji University, Gwalior. Poly (I:C) was administered to rat pups on postnatal day (PND) seven as described previously. 3 All the experiments were performed according to the instructions of Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), and approval of the Institutional Animal Ethical Committee, Jiwaji University, Gwalior.
The schematic timeline for drug administration and other molecular experiments is presented in Figure 1.
Analysis of Epigenetic Modifications
Following poly (I:C) administration, rats were sacrificed at different age of 3-, 6-, and 12-week and FC and HP were isolated and used for the analysis of epigenetic modifications.
Methylated DNA Immunoprecipitation (MeDIP)
The levels of 5mC at bdnf, arc, and egr1 promoters were analyzed by MeDIP. Initially, we isolated DNA from FC and HP of control and poly (I:C) exposed rats by the phenol-chloroform-isoamyl alcohol (PCI) method. Then DNA was fragmented (300–1,000bp) by sonication and quantified using a spectrophotometer. A small fraction (20%) of fragmented DNA was used as input DNA control. Thereafter 4 μg fragmented DNA was diluted in 1× immunoprecipitation medium and incubated overnight with 1 μg of 5mC antibody (A-1014; Epigentek, USA) for immunoprecipitation on ice with shaking. Then, 50 μl protein A-bead was mixed and incubated on ice with shaking for 2 h. Further, the precipitated complex bound with beads was setteled down by centrifugation at 3,500 ×g at 4°C for 10 min. After careful removal of supernatant, the pellet was washed with 1× TE buffer atleast three times followed by centrifugation. Finally, 200 μl elution buffer (100 mM NaHCO3 and 1% SDS) was added to the precipitated complex, DNA was precipitated, purified, and further dissolved in equal 20 μl TE buffer for each experimental group. Thereafter, qPCR was performed using eluted and input DNA (for normalization) as template and promoter specific primers to amplify specific promoter regions, 134 nucleotides of bdnf (F-5’-TGATCATCACTCACGACCACG-3’, R-5’-CAGCCTCTCTGAGCCAGTTACG-3’), 19 91 nucleotides of arc (F-5’-GGGTGGCTCTGAAGAATATTGG-3’, R-5’-CACCGAGCCCTGTTTGAACT-3’), 20 and 120 nucleotides of egr1 (F-5’-ACCACCCAACATCAGCTCTC-3’, R-5’-GTGGGTGAGTGAGGAAAGGA-3’). 21
Chromatin Immunoprecipitation (ChIP)
ChIP was performed to analyze H3K9 acetylation at histone bound to gene promoter sequence. Briefly, the FC and HP tissues were removed from the brain of experimental and control rats. Then tissues were chopped into small pieces in ice cold 1 × PBS and crosslinked with 1% formaldehyde at 25°C for 15 min. Thereafter, crosslinked tissues were homogenized in lysis buffer (5 mM KOH pH 8.0, 85 mM KCl, 0.5% NP-40 with 1 mM protease inhibitors), kept at 4°C for 5 min, and spun at 1,000 × g at 4°C for 5 min. After decanting supernatant, the pellet was dissolved in nuclear lysis buffer (50 mM Tris-Cl pH 8.0, 10 mM EDTA pH 8.0, 1% SDS with 1 mM protease inhibitors) followed by incubation at 4°C for 20 min and centrifugation at 12,000 × g for 15 min to settle the debris. The chromatin complex present in supernatant was stored in a fresh tube and protein was estimated by the Bradford method. 22 Furthermore, we diluted 200 μg chromatin in buffer containing 100 mM NaCl, 20 mM Tris-Cl pH 8.0, 2 mM EDTA, pH 8.0, 1% Triton X-100 with 1 mM protease inhibitors. The diluted chromatin complex was incubated with protein A-sepharose beads for 2 h for preclearing step to remove non-specific binding with beads and spun at 3,500 × g for 10 min. The precleared supernatant was transferred to a fresh tube and divided into input (20%) and immunoprecipitation fraction (80%). The immunoprecipitation fraction was incubated with H3K9Ac antibody (1 μg) overnight on ice. Then, protein A bead (50 μl) was added to it and kept for further incubation for 2 h on ice followed by centrifugation at 3,500 ×g for 10 min at 4°C. The supernatant was removed carefully, and pellet was washed sequentially with different solutions (low salt, high salt, LiCl, and TE buffer) followed by centrifugation. Thereafter, the precipitated complex was eluted with buffer (100 mM NaHCO3 and 1% SDS), mixed with 200 mM NaCl and incubated at 65°C for 4 h to reverse the cross-linking of protein-DNA complex. Then DNA was isolated from the precipitated and input fraction by the PCI method. Finally, these eluted and input DNA served as template for PCR amplification of gene promoter region using specific primers as described above.
Statistical Analysis
The data obtained from MeDIP and ChIP qPCR assay were statistically analyzed by SPSS 16.0 (Statistical Products and Service Solutions, IBM Corporation, Armonk, NY, USA) to check the significant mean difference between control and experimental groups. The p < .05 was considered as statistically significant. Histogram was plotted with mean ± SEM values to display the difference between poly (I:C) exposed experimental and control rats.
Results
Effect on DNA Methylation
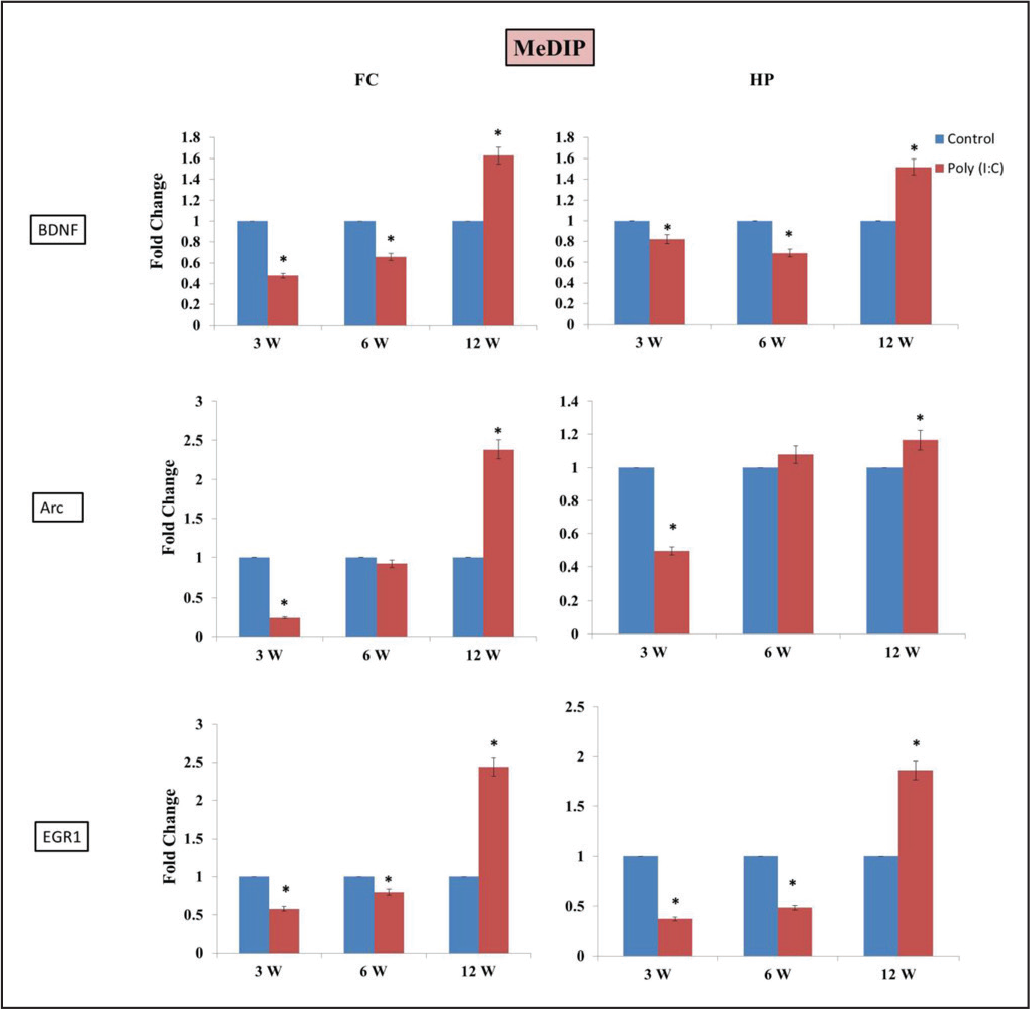
Alteration in 5mC level at gene promoter region tightly regulates its expression. To check this, we performed MeDIP-PCR of promoter sequence of bdnf, arc, and egr1 genes. As compared to vehicle control, a significant decrease was noted in 5mC level at the promoters of these genes in FC and HP regions of poly (I:C) exposed 3- and 6-week rats; except the arc promoter of 6-week rats which showed no difference in methylation level. However, in poly (I:C) exposed 12-week rats, 5mC level was increased in both regions as compared to control (Figure 2).
Effect on Histone Acetylation
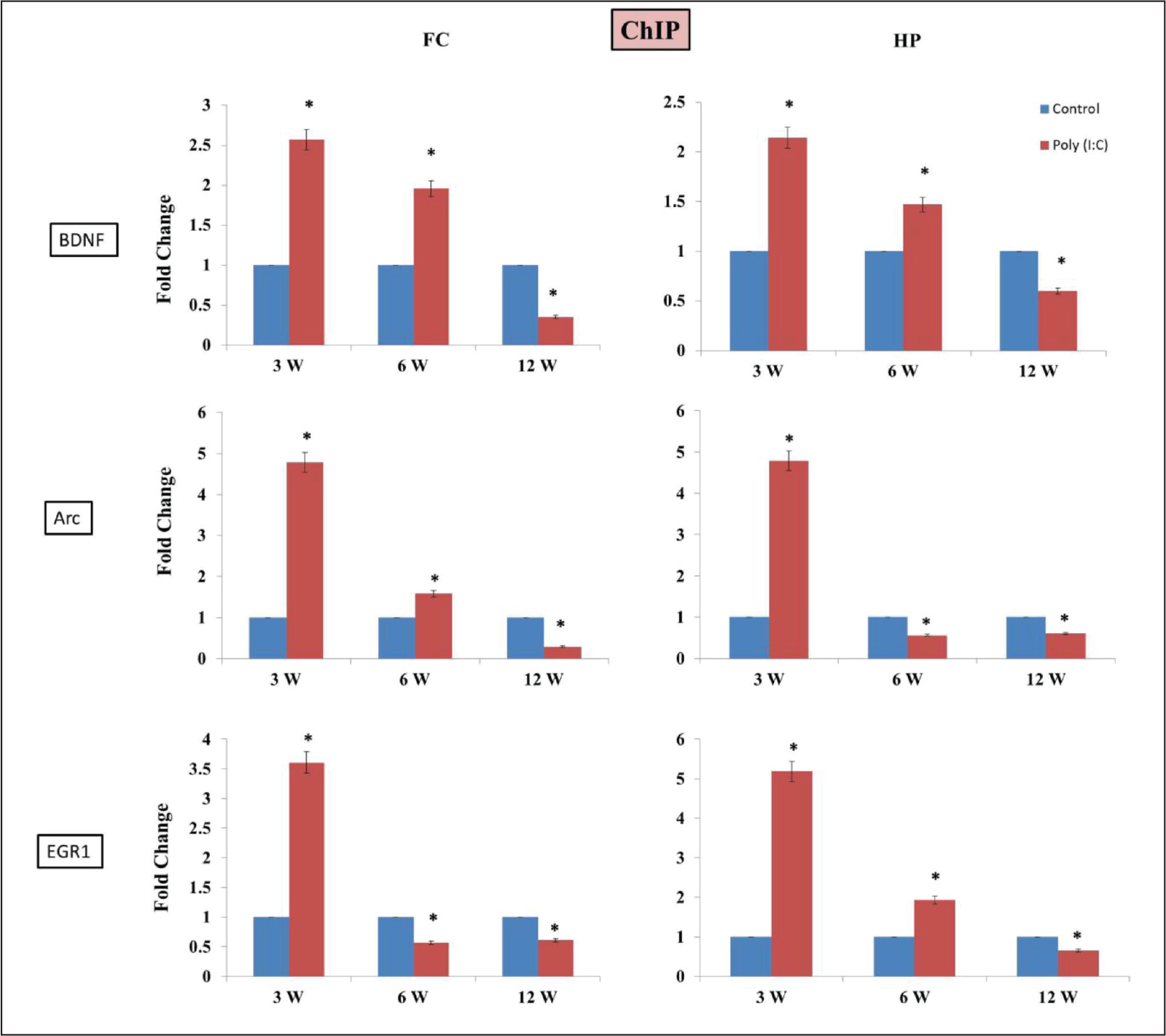
We have also analyzed histone H3K9Ac status at the promoter of these genes. The exposure to poly (I:C) significantly increased the H3K9Ac level at bdnf, arc, and egr1 promoter in both regions of 3-week rats. However, in 6-week, H3K9Ac level was increased at bdnf promoter in both regions but increased at arc promoter in FC and decreased in HP. Similarly, H3K9Ac at promoter of egr1 decreased in FC and increased in HP of poly (I:C) exposed 6-week rats. Further, it decreased at the promoter of these genes in both regions of poly (I:C) exposed 12-week rats (Figure 3).
Discussion
In the current investigation, we have analyzed epigenetic regulation of differentially expressed synaptic plasticity genes during postnatal exposure of poly (I:C) to developing rats. Poly (I:C) has been reported to cause cognitive deficits and alterations in epigenetic modifications leading to neurological disorders.6, 8, 9 Therefore, it was likely that poly (I:C) affects the epigenetic regulation of synaptic plasticity genes and consequently impairs memory. In epigenetic machinery, DNA methylation is involved in the development of neural system, cognitive functions, neurodevelopmental disorders and neurodegenerative diseases.23–25 Alteration in 5mC level is one of the causal factors for aging and related neurodegenerative diseases like AD. 26
We have found differential methylation pattern at the promoter of bdnf, arc, and egr1 genes in FC as well as HP of poly (I:C) exposed rats. It was initially increased in both brain regions of 3- and 6-week but later decreased in 12-week rats. In our previous publication, we reported that poly (I:C) injection activated and enhanced TNF-α positive microglia cells after 12 h of exposure in hippocampus (HP) and it persisted in 3- and 12-week rats. 3 Interestingly, TNF-α positive cells were less in number in 12-week rats as compared to early age (12 h, 3-week). Moreover, Zhao et al 27 have shown that the treatment of TNF-α decreases methylation level at IL-32 promoter and upregulates its expression in cell lines. This decreased methylation continues upto 30 days after the withdrawal of treatment, but thereafter it increases leading to decrease in the expression of IL-32 expression. Thus, TNF-α level might be playing a crucial role in decreasing DNA methylation initially in 3- and 6-week when the TNF-α level is high but increasing methylation in 12-week rats when the TNF-α level is relatively low.
Methylation pattern also corroborates with the expression pattern of these genes. 3 Similarly, alterations in DNA methylation and histone acetylation at bdnf, arc, egr1, homer1, and narp promoters influence their expression in the brain of aged and scopolamine-induced amnesic mice. 10 DNA methylation recruits the suppressor complex at promoter and prevents the interaction of transcription factors, thus reducing gene transcription. DNA methylation and demethylation events are important for synaptic plasticity and memory consolidation. 27 The inhibition of DNMT enzyme altered the level of DNA methylation and promoted the expression of synaptic plasticity reelin and bdnf genes in adult brain. 28 Hypermethylation of bdnf promoter resulted in its downregulation in schizophrenic patients. 29 In addition, Penner et al 30 have shown that alterations in arc expression and its promoter methylation are associated with impairment in spatial memory. Our results suggest that differential methylation status at promoter affects bdnf, arc, and egr1 expression in poly (I:C) induced infection. The balanced expression of BDNF is critical for synaptic plasticity, synaptogenesis, and formation of memory. 31 Arc is an important activity dependent cytoskeleton associated protein, which helps in actin polymerization, synaptogenesis, long term potentiation, dendritic arborization, and memory consolidation. 32 EGR1 is a member of immediate early gene and transcription factor, which regulates expression of different synaptic plasticity genes. 33 Any imbalance in the expression of these genes impairs their associated functions leading to abnormal behavior and cognitive functions including memory. 3 Moreover, poly (I:C) exposure to pregnant females reduces dendritic complexity and spine density leading to cognitive impairment. 34
Methylated DNA recruits HDAC2 repressor complex, which removes acetyl group and regulates H3K9Ac level. 35 Histone acetylation is crucial for memory acquisition and consolidation. Therefore, we have analyzed H3K9 acetylation level and found that in the promoter of bdnf, arc, and egr1 genes, H3K9 acetylation increased in the beginning at 3-week and later decreased at 12-week in FC and HP of poly (I:C) exposed rats. Acetylation level positively correlates with the expression pattern of these genes. 3 Poly (I:C) also alters histone acetylation by enhancing HDAC level. 17 Moreover, Fischer et al 36 have demonstrated increase in the acetylation of H3K9 and H3K14 with enrichment of environmental factors and recovery of memory in neurodegenerative disease model. H3 and H4 acetylation also decreases at egr1, c-fos, and bdnf promoter in AD.37–39
Conclusion
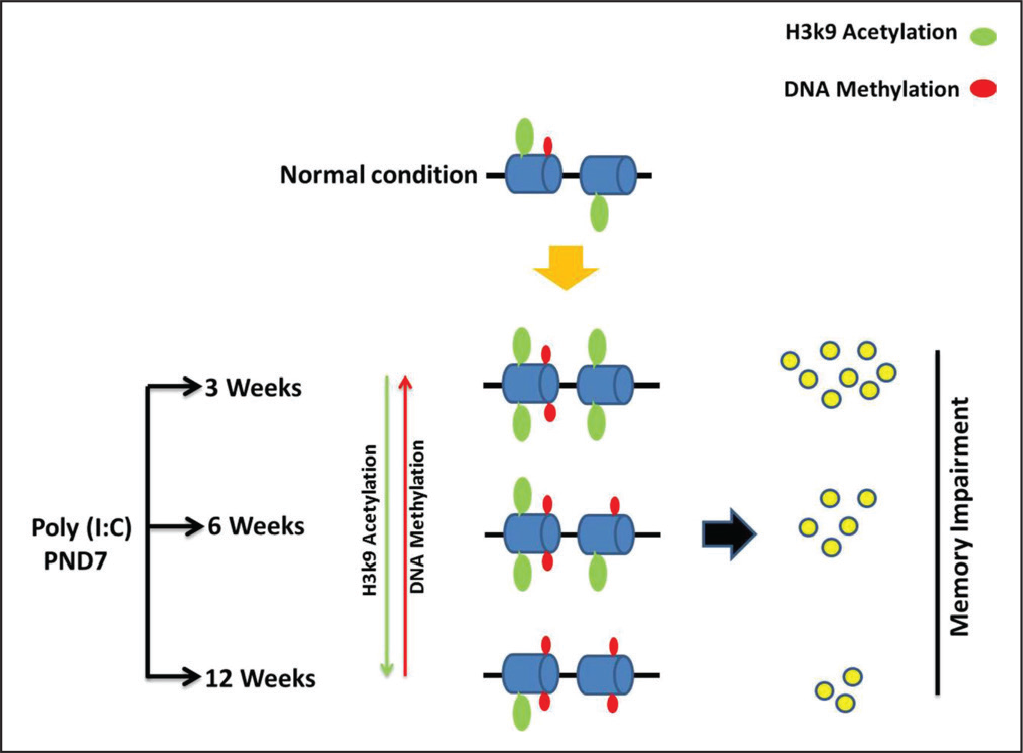
Our findings show that the influence of poly (I:C) exposure during early life also persists in later stages. The poly (I:C) exposure alters methylation of DNA and acetylation of histones at synaptic plasticity gene promoters and subsequently affects gene expression in FC and HP regions of rats even at later ages leading to impaired memory (Figure 4).
Footnotes
Acknowledgements
This work was conducted by three research centers under a collaborative project supported by the Department of Biotechnology (DBT), Ministry of Science and Technology, Government of India (BT/PR14311/MED/30/470/2010). We acknowledge UGC-UPE FA-II, and Interdisciplinary School of Life Sciences for qPCR facility; UGC-CAS in Zoology, Banaras Hindu University, Varanasi for SRF to MSB, and DBT-BET JRF program for DBT-SRF to BS.
Author Contributions
MS Baghel, N Patro, VK Khanna, IK Patro, and MK Thakur: planned and designed the experiments.
B Singh: Conducted poly (I:C) treatment and collected tissue samples.
MS Baghel: Carried out experiments, analysed data and drafted the manuscript.
VK Khanna, IK Patro, and MK Thakur: Edited the manuscript and corresponded with the journal.
Ethical Statement
The maintenance and experimental protocols were approved by the Institutional Animal Ethical Committee, Jiwaji University, Gwalior (Ref. no. IAEC/JU/2010/02) and carried out according to the instructions of Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for
the research, authorship and/or publication of this article: Department of Biotechnology (DBT), Ministry of Science and Technology, Government of India (BT/PR14311/MED/30/470/2010).