
Abstract
The natural cycle of hair growth and shedding is continuous. However, an abnormal rate of hair regrowth can signal alopecia, a condition characterized by excessive hair loss and thinning. Alopecia is a common dermatological concern globally, affecting individuals across all ages and sexes, often resulting in psychological distress and diminished quality of life. Despite the availability of various current therapy options such as minoxidil, finasteride, JAK inhibitors, low-level laser, and hair transplantation, these approaches are often limited by variable efficacy or side effects. Consequently, there is growing research interest in regenerative therapies using stem cells as a promising avenue to overcome these shortcomings. Stem cells, characterized by their ability to self-renew and differentiate into multiple cell types, offer the potential to promote hair follicle regeneration. Among the different stem cell types under investigation, hair follicle stem cells (HFSCs), mesenchymal stem cells (MSCs), and induced pluripotent stem cells (iPSCs) have attracted attention due to their regenerative capabilities and immunomodulatory properties. This narrative review explores the underlying causes of alopecia, current and emerging therapies, particularly emphasizing the therapeutic potential of stem cells and their specialized roles in promoting a supportive environment for the hair follicle and scalp. It also examines ongoing clinical and preclinical studies involving stem cell–based therapies for hair loss, aiming to assess their feasibility as future clinical solutions for alopecia.

Keywords
Introduction
“Alopecia” is a medical term for hair loss or baldness. For millions affected by alopecia, the sudden or gradual loss of hair can result in significant emotional and psychological consequences. Although it does not directly impact physical health, hair plays a crucial role in shaping personal identity and confidence 1 . Among the mild to severe signs of psychological issues linked to hair loss are suicidal thoughts and feelings, social disengagement, anxiety, rage, and a decline in confidence. These resemble those that are typically observed in severe, chronic illnesses that pose a serious threat to life 2 . Alopecia is a multifactorial condition influenced by factors including genetic predisposition, immune factors, exposure to toxic agents, hormonal imbalances, nutritional deficiencies, and metabolic dysfunctions3,4. These different elements can disrupt the normal hair growth cycle, which comprises anagen (growth), catagen (regression), telogen (resting), and exogen (shedding) phases. Under normal conditions, most scalp hairs are in the long anagen phase, ensuring continuous growth. In alopecia, this balance is disrupted by the shortened anagen phase, which causes either more hairs to enter catagen or telogen prematurely, or follicles lose their ability to cycle altogether 5 .
As various factors increase the transition from anagen to telogen, inflammation plays a central role in promoting this process, which is closely linked to various forms of hair loss, including androgenetic alopecia (AGA), alopecia areata (AA), stress-induced hair loss, and telogen effluvium (TE) 5 . Histological studies in both male and female alopecia consistently reveal chronic inflammatory infiltrates, particularly lymphocytes, histiocytes, and mast cells around hair follicles. This suggests local inflammation contributes to follicular miniaturization and premature anagen termination 6 . In addition, the immune factors, chemical exposure, and certain drugs can damage hair follicles and induce alopecia. Chemotherapy agents such as alkylating agents, anthracyclines, antimetabolites, vinca alkaloids, and taxanes commonly cause hair loss by targeting rapidly dividing anagen-phase cells 7 .
Hormones impact hair loss through different mechanisms. Multiple hormones influence the hair cycle, particularly dihydrotestosterone (DHT), thyroid hormones, and the estrogen-to-testosterone ratio. DHT is a potent derivative of testosterone. In individuals genetically predisposed to AGA, DHT binds to androgen receptors in hair follicles, shortening the hair growth cycle and causing follicular miniaturization 8 . Both hypothyroidism and hyperthyroidism can cause diffuse, reversible hair loss, often through TE 9 . Thyroid hormone signaling regulates epidermal proliferation and activation of stem cells in the hair bulge. Deficiency impairs stem cell mobilization, while excess stimulation depletes stem cells, meaning both low and high thyroid hormone levels can disrupt the hair cycle 10 . Estrogen has protective effects on hair follicles; elevated during pregnancy, it prolongs anagen, while postpartum or menopausal estrogen decline leads to TE or female-pattern alopecia (FPHL) 11 . It has been suggested that the cessation of ovarian estrogen production and complex interactions with other hormones, growth factors, and cytokines cause changes in hair growth properties 12 .
The metabolic dysfunction, including thyroid dysfunction, insulin resistance, polycystic ovary syndrome (PCOS), and nutritional deficiencies, plays a critical role in AGA 13 . Thyroid dysfunction alters the cycling and structure of hair follicles. Hypothyroidism slows follicular metabolism and delays the telogen-to-anagen transition due to reduced thyroid hormone levels. Whereas hyperthyroidism accelerates metabolism, which leads to oxidative stress, mitochondrial dysfunction, and lipid peroxidation within hair follicle cells, leading to fine, fragile hair and increased shedding 10 . Insulin resistance plays a central role in the development of PCOS and hyperandrogenism in women. Elevated insulin levels stimulate ovarian androgen production and suppress hepatic synthesis of sex hormone-binding globulin (SHBG), resulting in higher levels of free testosterone and other androgens. Increased androgens promote hair follicle miniaturization, contributing to scalp hair loss, particularly in FPHL14,15. Nutritional deficiency can impair hair structure, growth, and pigmentation. While vitamins, minerals, fatty acids, and proteins have been studied for their roles in hair health, the benefits of supplementation remain uncertain, and excessive intake may cause toxicity or worsen hair loss5,16.
However, alopecia is not a one-size-fits-all condition. The severity, progression, and underlying causes vary widely depending on the type. Alopecia can be classified as either non-cicatricial (nonscarring) alopecia, in which hair loss occurs without the destruction of hair follicles, or cicatricial (scarring) alopecia, in which the skin appears atrophic, scaling, and smooth with no hair follicles1,4.
Methods
A narrative literature review was conducted to summarize current and emerging evidence on therapies for hair regeneration, with a specific focus on stem cell–based approaches and their regenerative potential. Relevant studies were identified through searches on electronic databases, including PubMed and Scopus. Keywords such as alopecia, hair regeneration, AGA, AA, minoxidil, finasteride, low-level laser therapy (LLLT), platelet-rich plasma (PRP), hair transplantation, stem cell therapy, HFSC, MSCs, iPSCs, dermal papilla cells (DPCs), regenerative medicine, extracellular vesicles, and tissue engineering were applied. A systematic review of clinical trials was conducted on ClinicalTrials.gov. The keywords “Hair loss” as a condition or disease and “Stem cell therapy” were used. The search included studies published from 2000 to 2026, with searching criteria including clinical trials, systematic reviews, meta-analyses, and original research articles investigating the pathogenesis, pathophysiology, and current and emerging treatments for alopecia, especially stem cell-based. Abstracts of conferences and non-peer-reviewed publications, as well as studies with inadequate statistical rigor, and data redundancy or duplication, were excluded.
Type of non-cicatricial (nonscarring) alopecia
Androgenetic alopecia
AGA, the most common form of hair loss, affects both men and women. This condition affects up to 80% of men and 50% of women 17 . The majority of the men were 20–39 years old, and the women were 60–69. Men typically have an earlier onset of AGA than females 18 . AGA is a complex condition rooted in a combination of genetic predisposition and hormonal influences. Individuals inherit specific genes, most notably variations in the Androgen Receptor (AR) gene, that make their scalp’s hair follicles exceptionally sensitive to androgens. The primary hormone driving the hair loss is DHT, which is converted from testosterone. When DHT binds to these genetically hypersensitive receptors in the hair follicles, it initiates a process called follicular miniaturization, causing the hair to progressively shrink in diameter and length with each growth cycle, ultimately leading to the characteristic pattern of baldness17,19,20.
Alopecia areata
The pathophysiology of AA is rooted in an autoimmune response where the body’s own defense system mistakenly targets its hair follicles. Normally, hair follicles enjoy a state of “immune privilege,” which shields them from attack by immune cells 21 . In AA, after the hair follicle loses its natural immune protection, CD8+ cytotoxic T cells act as the primary aggressors, swarming the hair bulb and releasing inflammatory signals that shut down hair production. CD4+ helper T cells orchestrate this assault and persist because regulatory T cells (Tregs) fail to suppress the rogue immune response, and the follicle prematurely enters a resting state, halting hair production and causing the hair shaft to fall out22,23.
Telogen effluvium
TE is a form of diffuse, nonscarring alopecia characterized by a premature, synchronized shift of a large population of hair follicles from the anagen phase into the telogen phase 24 . TE is a physiological response to a significant systemic trigger, such as major illness, surgery, childbirth, or severe emotional distress. The affected follicles enter a latent telogen phase, typically lasting 2–4 months. The clinical manifestation, the effluvium, or shedding occurs as these resting hairs are subsequently extruded from the scalp 25 .
Type of cicatricial (scarring) alopecia
Frontal fibrosing alopecia
Scarring (cicatricial) alopecia, such as the primary lymphocytic cicatricial alopecia known as frontal fibrosing alopecia (FFA), primarily affects postmenopausal Caucasian women. FFA’s poorly understood etiology may involve hormonal or environmental triggers. It is distinguished by a progressive recession of the frontotemporal hairline, which manifests as a scarring band of hairlessness and is frequently followed by loss of body and eyebrow hair 26 . Recent evidence suggests that epithelial-mesenchymal transition (EMT) plays a central role in its pathogenesis, indicating a disruption in follicular immune privilege and fibrotic remodeling of the scalp. Environmental exposures, particularly to certain cosmetic products, have also been implicated as potential contributing factors 27 . Therapeutically, a systematic review and meta-analysis involving 366 patients demonstrated that oral dutasteride effectively stabilized hair loss in 63% of cases, with 36% experiencing clinical improvement, supporting its role as a viable treatment option for FFA 28 .
Lichen planopilaris
Similarly, lichen planopilaris (LPP) and central centrifugal cicatricial alopecia (CCCA) have drawn attention because of their irreversible nature and complex immune-mediated destruction of hair follicles 29 . LPP is an inflammatory, primary cicatricial alopecia that presents with different hair loss patterns. Its typical clinical presentation includes scaly, erythematous plaques resulting in irreversible alopecia 30 . Between 25 and 70 years old is the typical age at which LPP first appears, and its incidence ranged from 1.15% to 7.59% 31 .
Central centrifugal cicatricial alopecia
In contrast to LPP, CCCA is a unique type of scarring alopecia that is typified by patches of permanent hair loss that appear on the scalp’s vertex or crown and gradually extend outward in a centrifugal trajectory 32 . This illness is characterized by visible inflammation and is more common in African American women, suggesting a genetic predisposition. The treatment modalities include topical and systemic corticosteroids and oral tetracyclines, with a limited response (Fig. 1) 33 .

The causes and major types of alopecia: (a) non-cicatricial (nonscarring) alopecia, including androgenetic alopecia, alopecia areata, and telogen effluvium and (b) cicatricial (scarring) alopecia, including frontal fibrosing alopecia, lichen planopilaris, and central centrifugal cicatricial alopecia. (Created by BioRender.com/Mahidol University.)
Alopecia current therapies
As the understanding of alopecia pathophysiology advances, several treatment options have been developed to manage and potentially reverse hair loss. Current treatment strategies primarily focus on halting hair loss progression, stimulating hair regrowth, and modulating the underlying immune or hormonal mechanisms (Table 1).
Summary of alopecia current therapies.

Topical minoxidil
Among these, minoxidil remains one of the most extensively utilized and FDA-approved topical therapies, particularly for AGA. Minoxidil, commonly applied topically as a solution or foam, is effective in promoting hair regrowth. Although its exact mechanism remains unclear, it is believed to prolong the anagen phase, increase vascular endothelial growth factor (VEGF) expression, and improve blood flow around hair follicles. Available in 2% and 5% concentrations, the 5% formulation is more effective, especially in men with AGA, and is increasingly used in women for its superior results 42 . Recently, low-dose oral minoxidil (0.25–5 mg/day) has shown a favorable safety and efficacy profile for treating AGA. However, when low-dose oral minoxidil is used, the most common adverse effects are hypertrichosis and cardiovascular symptoms (CVS). CVS symptoms included hypotension, edema, premature ventricular contractions, fatigue, tachycardia, and electrocardiogram (ECG) changes 43 . While minoxidil does not address the root causes of alopecia, it remains a key symptomatic treatment. It is often combined with other therapies, such as finasteride, PRP, and stem cell–based approaches, to enhance outcomes in alopecia management44–46.
Oral finasteride
Finasteride is a key pharmacological treatment for AGA, particularly in men, and functions by addressing the hormonal basis of hair loss. It is a 5-alpha reductase inhibitor that selectively blocks the type II isoenzyme, preventing the conversion of testosterone to DHT, a potent androgen that contributes to follicular miniaturization in AGA 47 . Clinical studies have demonstrated that oral finasteride at a dose of 1 mg/day can significantly increase hair count, thicken existing hairs, and slow disease progression over the course of several months to years 48 . It is approved by the FDA for male-pattern hair loss and is commonly used off-label in postmenopausal women with caution, often in combination with other antiandrogens 49 . Finasteride is generally well tolerated, but its use has been linked to rare side effects, including sexual dysfunction, depression, and, in some cases, post-finasteride syndrome (PFS). PFS is a controversial and debated condition involving persistent side effects after discontinuation 50 . Topical finasteride formulations are emerging as an alternative, aiming to reduce systemic exposure while maintaining efficacy 51 . Recent randomized trials and observational studies suggest that topical 0.25% finasteride can decrease scalp DHT levels significantly with fewer systemic effects 52 . As research continues to optimize delivery methods and minimize risks, finasteride remains a cornerstone of evidence-based management for hormonally driven alopecia.
JAK inhibitors
Since 2022, the FDA has approved three JAK inhibitors. Baricitinib became the first FDA-approved systemic treatment for severe AA in June 2022 53 . Ritlecitinib followed this approval in June 2023 and deuruxolitinib in July 2024 54 . The JAK-STAT signaling cascade regulates cytokine-mediated immune responses. In AA, interferon-gamma (IFN-γ) activates the JAK1/2 signaling pathway in hair follicle epithelial cells, leading to the upregulation of interleukin (IL)-15. IL-15 is then presented by hair follicles to CD8+ T cells, activating JAK1/3 signaling and promoting further IFN-γ production. This creates an inflammatory loop that sustains immune-mediated hair follicle damage 55 . Baricitinib acts by selectively inhibiting JAK1/JAK2, thereby interrupting the inflammatory cascade and facilitating hair regrowth 56 . Recent clinical trials, two separate randomized phase III clinical trials (BRAVE-AA1 and BRAVE-AA2), have demonstrated the efficacy of Baricitinib in inducing significant hair regrowth in a proportion of patients over 36 weeks, with some achieving ≥80% scalp coverage 57 . However, its use is tempered by significant limitations: the randomized clinical trial of 654 adults with severe AA, including relapse upon withdrawal, high cost, and unknown long-term risks 58 .
Low-level laser therapy
LLLT, also referred to as photobiomodulation, has gained recognition as a non-invasive and effective treatment for various types of alopecia, especially AGA in both men and women 59 . LLLT devices emit red or near-infrared light (typically 600–950 nm) that penetrates the scalp and stimulates cellular activity in hair follicles 60 . Mechanistically, LLLT promotes mitochondrial ATP production, modulates reactive oxygen species (ROS), and activates transcription factors that enhance the proliferation of follicular keratinocytes, which consequently prolong the anagen phase of the hair cycle. These biological effects support hair regrowth, increased hair density, and improved follicle health 61 . FDA-cleared devices, including laser combs, caps, and helmets, have become widely available for home or clinical use 62 . Although LLLT is not curative and requires long-term maintenance, it is well tolerated with minimal side effects. Its non-pharmacological nature makes it especially suitable for patients seeking drug-free alternatives, or combination therapies such as a combination of LLLT with topical minoxidil and oral finasteride, which may act synergistically to enhance hair growth 59 .
Hair transplantation
Hair transplantation, through follicular-unit grafting, is the preferred surgical method for individuals with specific forms of alopecia, such as AGA 39 . The two primary techniques, follicular unit transplantation (FUT) and follicular unit extraction (FUE), have both evolved with improved precision and outcomes. FUE, in particular, has become the preferred method due to its minimally invasive nature, reduced scarring, and quicker recovery. It involves the extraction of individual follicular units from the donor area (usually the occipital scalp) and their implantation into bald or thinning regions 63 . Advanced robotic-assisted FUE systems and microsurgical tools have further increased graft survival and natural appearance. Modern hair transplantation focuses on aesthetic design, density distribution, and long-term graft viability. Success depends on the surgeon’s expertise, donor site health, and post-operative care 64 . While highly effective, the procedure is costly and not suitable for all types of alopecia, such as diffuse AA. Follicular unit hair transplantation is widely accepted in clinical dermatology as a gold-standard intervention for eligible alopecia patients because it preserves the natural architecture of the hair units and gives natural results 65 .
Alopecia emerging regenerative therapies
Microneedling
Microneedling involves the use of fine needles to create controlled micro-injuries on the skin surface, which initiates a cascade of wound-healing responses. These include the release of platelet-derived growth factor (PDGF), epidermal growth factor (EGF), fibroblast growth factor (FGF), and VEGF, all of which contribute to angiogenesis, extracellular matrix remodeling, and activation of dermal papilla stem cells 66 . Kim et al. 67 applied repeated microneedle stimulation to the dorsal skin of mice and observed significant hair regrowth compared to control groups. Importantly, immunohistochemical staining demonstrated elevated expression of β-catenin in the treated skin areas, indicating that microneedling upregulates the Wnt/β-catenin signaling pathway, which is essential for hair follicle morphogenesis and cycling. Despite promising results, microneedling faces several limitations. Transient erythema and hyperpigmentation following inflammation were the most frequent adverse reactions to microneedle treatment, according to Hou et al. 68 , whereas severe and long-lasting adverse outcomes were uncommon. However, the absence of long-term safety and efficacy research is one of microneedling therapy’s drawbacks.
Platelet-rich plasma
PRP therapy holds a significant, though still adjunctive, role in the treatment of alopecia, particularly AGA and AA69,70. PRP, derived from autologous blood, contains concentrated growth factors that stimulate hair follicle activity, enhance angiogenesis, and extend the anagen phase of the hair cycle. Recent studies continue to demonstrate PRP’s efficacy in promoting angiogenesis, along with vascularization, and promoting hair follicles to enter and extend the anagen phase 71 . PRP used for hair regeneration can be prepared as either activated or non-activated PRP. Activation of PRP is typically achieved by adding agents such as calcium chloride (CaCl2) or thrombin, which trigger the initial blood clotting and fibrin scaffold formation. This results in the induction of intermediate release of growth factors, including PDGF, transforming growth factor-beta (TGF-β), and VEGF, which play important roles in promoting dermal papilla cell proliferation and angiogenesis in hair follicles72,73. In contrast, non-activated PRP relies on physiological activation after administration, where platelets gradually release bioactive molecules upon contact with collagen or thrombin at the target tissue 74 .
Its use is especially effective when combined with conventional treatments such as minoxidil, microneedling, LLLT, and hair transplantation, demonstrating by increasing hair density and hair diameter compared to monotherapy 75 . According to Suo et al., a microneedle system loaded with PRP lysate was designed to deliver GFs transdermally to sites of hair loss. This procedure promoted hair regeneration by enhancing cell proliferation and angiogenesis without inducing significant discomfort 76 . A randomized blinded retrospective study investigating the combined use of microneedling, LLLT, and autologous non-activated PRP demonstrated promising results in patients with androgenic alopecia. The study showed a significant increase in hair density, with average gains of 81 ± 5 and 57 ± 7 hairs/cm2 at 12 and 23 weeks, respectively, compared to baseline. These outcomes were observed using computerized trichograms 77 . While PRP is generally safe and well tolerated, results vary due to non-standardized protocols and patient-specific factors. Despite growing clinical support, PRP therapy remains unapproved by the FDA for alopecia treatment due to limited large-scale randomized trials. Nevertheless, it is widely used off-label in dermatology, reflecting its emerging value as a minimally invasive, regenerative option for hair loss management 78 .
Stem cell therapies
Stem cells, melanocytic cells, and epithelial cells from various developmental origins are among the diverse cell resources found in hair follicles that may continuously renew, differentiate, and regulate skin homeostasis and hair growth 79 . Stem cells in hair follicles play a crucial role in the growth, regeneration, and cycling of hair. These cells are a type of adult stem cell with the ability to self-renew and differentiate into various cell types that make up the hair follicle and complex tissue microenvironment73,80. The hair follicle is a mini-organ that undergoes a hair development cycle. It relies on a coordinated network of stem cells to sustain its function throughout life. These hair follicle–associated stem cells not only orchestrate the continual renewal of hair but also contribute to skin repair and pigmentation81,82. Within the hair follicle, several distinct populations of adult stem cells can be identified, each performing specialized roles.
Hair follicle stem cell–based therapies
Hair follicle stem cells (HFSCs) are multipotent adult stem cells localized in the bulge region of the hair follicle 83 . These cells exhibit the capacity for self-renewal and can differentiate into multiple epithelial lineages of the hair follicle, including outer root sheath cells (ORSCs), inner root sheath cells, and hair shaft progenitors84–86.
A systematic review from Hu et al. 87 found that the activation of HFSCs during the initiation of the anagen phase is regulated by a complex interplay of signaling pathways, including Wnt/β-catenin, bone morphogenetic protein (BMP), Notch, and Sonic Hedgehog (Shh) (Fig. 2). Specifically, Wnt/β-catenin and Sonic Hedgehog act as key activators of HFSCs, while BMP maintains quiescence and Notch contributes to the regulation of HFSC fate and differentiation 88 . Problems with HFSCs function or their surrounding environment (niche) are important factors in the shrinking of hair follicles and the reduced ability to regenerate seen in different types of hair loss; specifically, HFSC niche cells are arranged into groups that help send signals, sense changes, and communicate, enabling HFSCs to adapt their regeneration based on the body’s needs and changes in the environment89–91. AGA is associated with a decline in HFSCs, particularly progenitor cells, leading to miniaturization of hair follicles and hair loss. While the quiescent stem cells remain, there is a defect in their conversion to more proliferative progenitor cells, disrupting the normal hair growth cycle 92 . Similarly, in aging, a decline in HFSC number and a deterioration of the niche contribute to progressive hair thinning. A study from Koester et al. 93 revealed that aging leads to stiffening of the HFSC niche due to alterations in the extracellular matrix. This mechanical change reduces chromatin accessibility at key self-renewal and differentiation genes in HFSCs, impairing their function. Furthermore, Zhang et al. 94 found that, in aged mice, epithelial cells escape from the HFSC compartment to the dermis, contributing to hair follicle miniaturization. In autoimmune conditions such as AA, the immune privilege of the HFSC niche is disrupted, permitting immune surveillance and subsequent cytotoxic T-cell infiltration. However, the long-lived, quiescent HFSCs are strikingly resistant to cytotoxic immune effectors such as T cells and natural killer (NK) cells, even in systems where these cells are artificially directed to attack HFSCs 95 .

The hair cycle and signaling pathway. (a) In the early anagen (growth initiation) phase, growth is initiated by the Wnt/β-catenin signaling pathway, which is the master activator for follicle stem cell proliferation. The key inhibitory pathway, BMP signaling, must be suppressed or low for anagen to proceed. (b) Late anagen (active growth/differentiation) is the phase of rapid cell division and formation of the hair shaft. Both Wnt and Shh signaling remain highly active, driving the intense proliferation of matrix cells and the elongation of the hair shaft. Notch signaling becomes essential, regulating the differentiation of progenitor cells into the various layers of the inner root sheath and the hair shaft itself, ensuring proper structure. (c) Catagen (regression/transition): the controlled self-destruction and shrinking of the lower follicle. BMP signaling reactivates and significantly increases, promoting the programmed cell death (apoptosis) and driving the regression of the follicle. Notch signaling helps synchronize the balance between proliferation and the programmed differentiation/regression events during this transition. (d) The telogen (rest) phase is maintained by the dominance of inhibitory signals, primarily high levels of BMP signaling, which keep the hair follicle stem cells in a quiescent (inactive) state, preparing for the next cycle. (Created by BioRender.com/Mahidol University.)
HFSC transplantation and injection therapies aim to restore hair growth by replenishing or activating the follicular stem cell pool in areas affected by alopecia 96 . In these approaches, HFSCs or progenitor cells are typically harvested from a patient’s own scalp tissue, processed to isolate the stem cell-rich fraction, and then reinjected into balding regions. These transplanted or injected cells can stimulate the regeneration of existing follicles, promote the transition of follicles into the anagen phase, and enhance the overall density and thickness of hair97,98. Some studies have demonstrated that HFSC injections can reactivate dormant follicles by releasing growth factors and signaling molecules that modulate the Wnt/β-catenin and Sonic hedgehog pathways87,98. Although still in the experimental stage, autologous HFSC transplantation represents a promising regenerative approach for AGA and other forms of hair loss, offering potential for follicle rejuvenation without the need for full follicular grafts 99 . However, the clinical translation of HFSC-based therapies is tightly governed by regulatory frameworks, especially the concept of minimal manipulation, which plays a pivotal role in determining the classification, safety, and legal use of these cellular products. Minimal manipulation refers to processing that does not alter the original relevant biological characteristics of the cells or tissue 100 . This technique focuses on the physical disaggregation of tissue, such as mechanical isolation, centrifugation, or filtration, to produce a cell solution, which is permitted within regulations as it does not involve enzymatic digestion or prolonged in vitro expansion to enhance proliferation101,102.
In the context of HFSCs, they fall under the category of more-than-minimally manipulated products when isolated using enzymatic digestion, subjected to cell sorting, or expanded in culture, as these methods enable the expansion of cell numbers, selective enrichment, or functional enhancement of HFSCs103,104. However, they may alter the cell structural and functional properties, exceeding the threshold of minimal manipulation as defined by the US Food and Drug Administration 102 .
Dermal papilla cell–based therapies
Alongside HFSCs, DPCs, which are a specialized mesenchymal population, reside at the base of the follicle and provide essential inductive signals that influence the activity of the cells that generate and regenerate the follicle 105 . Moreover, DPCs remain closely associated with the epithelial progenitor populations of the hair follicle, even as the follicle undergoes dynamic structural changes. DPCs are capable of secreting various signaling factors that initiate hair follicle formation by activating skin epithelial stem cells (Epi-SCs) 106 . Consequently, the combined use of DPCs and Epi-SCs has been shown to promote functional hair follicle regeneration in vivo106,107. Several experimental studies have demonstrated the efficacy of combining DPCs and Epi-SCs in vivo. Notably, injecting a mixture of human-derived epithelial stem cells and DPCs into immunodeficient mice led to the formation of de novo hair follicles with a proper multi-layered epidermis resembling native hair shafts 108 . However, such approaches face clinical application challenges, as both human DPCs and Epi-SCs tend to lose their hair-inductive properties during extended in vitro culture, primarily due to downregulation of critical signaling pathways such as Wnt, BMP, and FGF109,110.
Stem cell therapy involves the isolation of HFSCs, mesenchymal stem cells (MSCs), or induced pluripotent stem cells (iPSCs) to rejuvenate damaged follicles or generate new ones. These cells can differentiate into dermal papilla-like cells. Importantly, transplanted DPCs can induce new hair follicle formation and release trophic factors that promote hair follicle neogenesis and prolong the anagen phase 105 .
Mesenchymal stem cell–based therapies
MSCs are multipotent stromal cells capable of differentiating into mesodermal lineages, such as osteoblasts, adipocytes, and chondrocytes, as well as ectodermal and endodermal lineages. They are commonly harvested from bone marrow, adipose tissue, and umbilical cord111,112.
Bone marrow–derived mesenchymal stem cells
Bone marrow was the first identified source of MSCs and remains a well-characterized population for therapeutic applications 113 . Bone marrow–derived mesenchymal stem cells (BM-MSCs) exhibit strong immunomodulatory effects, largely through the secretion of interleukin-10 (IL-10), TGF-β, and prostaglandin E2 (PGE2), which help suppress T-cell activation and inflammatory cytokine production 114 . In the context of alopecia, BM-MSCs are particularly valuable for treating autoimmune and inflammatory subtypes. Harvesting bone marrow is more invasive than adipose tissue extraction. Byun et al. 115 investigated the therapeutic potential of BM-MSCs in a mouse model of AA and demonstrated that intravenous administration of BM-MSCs effectively prevented the development of AA. The protective effect was attributed to the suppression of key pro-inflammatory cytokines, including IFN-γ, CXCL9, and CXCL10, and a significant reduction in the infiltration of CD3+ and CD8+ NKG2D+ cytotoxic T cells in the affected skin. The results show that the immunomodulatory role of BM-MSCs, particularly their ability to downregulate NKG2D expression, mitigates the autoimmune response associated with AA.
Adipose-derived mesenchymal stem cells
Adipose tissue is one of the richest and most accessible sources of MSCs. The process of using MSCs derived from adipose tissue to treat alopecia is a two-component process: the isolated and cultured pure MSCs are known as adipose-derived stem cells (ADSCs). These cells secrete growth factors that play critical roles in the activation of epidermal stem cells and human dermal papilla cells (hDPCs), such as VEGF, hepatocyte growth factor (HGF), insulin-like growth factor-1 (IGF-1), and PDGF. These factors are essential for encouraging hair growth and improving blood flow in the scalp; and freshly derived primary multipotent MSCs that are a part of the stromal vascular fraction (SVF), also known as adipose-derived stromal vascular cells (ADSVCs) or adipose-derived regenerative cells (ADRCs)116,117. SVF is crucial for the repair and regeneration of tissue that has been injured over time, encourage hair development by increasing dermal papilla’s ability to produce or grow new hair, as a study by Stevens et al. 118 evaluated injections of SVF, which is rich in adipose-derived stromal cells (ASCs) in combination with PRP in the upper scalp as a new autologous treatment option for AGA. This study investigation involved 10 male participants, aged between 25 and 72 years, all diagnosed with moderate AGA (Norwood-Hamilton stages II–III). Each subject received a single autologous intradermal injection of platelet-rich stroma (PRS) into the affected scalp region. Clinical outcomes were assessed at baseline, 6 weeks, and 12 weeks postinjection. Changes in hair density were assessed using ultra-high-resolution photography. Results demonstrated a statistically significant increase in hair density at both 6 weeks (P = 0.013) and 12 weeks (P < 0.001) compared to baseline. Notably, new hair growth was observed from both active and previously dormant follicles, suggesting regenerative activity induced by the PRS formulation.
Umbilical cord–derived mesenchymal stem cells
Umbilical cord tissue, particularly Wharton’s jelly, provides an easily accessible, easy to isolate, and ethically acceptable, they represent a promising source of MSCs for use in regenerative medicine. Umbilical cord–derived MSCs (UC-MSCs) are more primitive, proliferative, and immunosuppressive compared to AD-MSCs and BM-MSCs119,120. UC-MSCs are immunologically immature and exhibit high proliferative capacity, making them ideal for allogeneic use. Several studies have investigated the efficacy of UC-MSCs for hair loss. For example, a case study of Ahn et al. 121 used allogenic, minimally manipulated umbilical cord MSCs for the successful treatment of AA and alopecia universalis. Furthermore, a study by Ni et al. 122 investigated the hair regrowth potential of conditioned medium (CM) derived from human umbilical cord mesenchymal stem cells (HuMSCs) cultured in a three-dimensional gelatin sponge scaffold (GS-HuMSCs-CM). Compared to conventional two-dimensional cultures, the 3D scaffold significantly enhanced the secretion of key growth factors, including VEGF, IGF-1, keratinocyte growth factor (KGF), and HGF. When applied to a C57BL/6J mouse model, GS-HuMSCs-CM presented more enhanced effects in hair length, hair diameter, and growth rate. GS-HuMSCs-CM significantly promoted angiogenesis in the injected skin areas, which might also contribute to faster hair regrowth 122 .
Immune privilege and hair follicle regeneration roles of MSCs in alopecia
MSCs secrete paracrine factors such as EGF, FGF, PDGF, VEGF, TGF-β, HGF, IGF-1, PGE2, stromal cell-derived factor-1 (SDF-1), angiopoietin-1, indoleamine 2,3-dioxygenase (IDO), IL-6, and IL-10 123 . The secretion of soluble paracrine factors has been proposed as an alternative mechanism, and this is termed the “paracrine hypothesis.” A systematic review identifies secreted paracrine factors that could improve the survival of other cell populations and promote skin healing, including anti-inflammatory and immunomodulatory activity, anti-apoptotic activity partly based on Hsp27 upregulation, increased angiogenesis, cell proliferation, and cell migration124,125.
Furthermore, MSCs have strong anti-inflammatory and modulatory effects on innate and adaptive immunity via interactions with immune cells through cell-to-cell contact and paracrine activity involving T cells, B cells, NK cells, macrophages, monocytes, dendritic cells (DCs), and neutrophils 126 . One of the key immunological targets of MSCs is the T lymphocyte population. MSCs inhibit the proliferation of T cells, specifically pro-inflammatory populations of T-helper cells (Th17 and Th1) and CD8+ cytotoxic T cells, and decrease the ratio of Th1/Th2 T-helper cell populations, both of which play central roles in the pathogenesis of AA 127 . Park S-H et al. 128 demonstrated that locally administered MSC therapy effectively improves AA in an IFN-γ-induced mouse model. MSC-treated mice showed better hair regrowth and reduced local skin inflammation, with significant decreases in JAK/STAT-related and pro-inflammatory cytokines (JAK1, JAK2, STAT1, STAT3, IFN-γR, IL-1β, IL-16, IL-17α, and IL-18). MSCs also restored key growth factors (FGF7, FGF2) and reactivated Wnt/β-catenin-related genes (LEF1, β-catenin), supporting normalization of the hair cycle 128 . Also, MSCs facilitate the polarization of macrophages from a pro-inflammatory M1 phenotype to an anti-inflammatory M2 phenotype, thereby attenuating local inflammation and establishing a microenvironment conducive to hair follicle repair and regeneration129,130. In alopecia, M1 macrophages predominate and secrete pro-inflammatory mediators, particularly IL-18 and IL-1β, which collectively induce follicular stress responses, compromise the immune privilege of hair follicles, and accelerate the transition into the catagen phase of the hair cycle 131 . MSCs mitigate these effects by promoting a phenotypic shift toward M2 macrophages, which are characterized by their anti-inflammatory activity and the secretion of growth factors that promote hair growth, such as HGF, IGF-1, and VEGF. These factors contribute to wound-induced hair neogenesis (WIHN), whereas angiogenesis is associated with the active anagen phase132–134.
Safety concerns and regulatory considerations of MSCs in hair regeneration
MSC therapy for hair regeneration is generally considered safe and well tolerated, with most clinical studies reporting only mild local reactions such as redness, swelling, irritation, pain, or headache (Table 2). A primary issue is tumorigenicity, as MSCs possess high proliferative capacity, resistance to apoptosis, and the ability to modulate the tissue environment. In addition, MSCs may promote tumor progression by homing to tumor sites, secreting proangiogenic factors, and differentiating into tumor-associated stromal cells such as carcinoma-associated fibroblasts (CAF) 135 . Immune-related reactions can also occur, particularly with allogeneic MSCs, leading to localized inflammation such as erythema and mild pain at injection sites. Infection risk is another concern, mainly associated with improper cell harvesting, processing, or injection under non-sterile conditions. Additionally, variability in MSC sources and preparation protocols, including differences in adipose-, bone marrow-, or umbilical cord-derived cells as well as xeno-contamination reactions caused by components of culture medium, can affect safety and reproducibility 136 . Genetic instability may arise during in vitro expansion, with prolonged culture leading to chromosomal abnormalities. There is also a low potential risk of unintended differentiation, which could result in fibrosis or abnormal tissue formation 137 . Overall, while adverse effects are typically mild and transient in hair regeneration applications, the lack of standardized procedures and long-term follow-up data remains a significant limitation in fully establishing the safety profile of MSC-based therapies 136 .
Summary of clinical trials investigating mesenchymal stem cells (MSCs) for hair regeneration.

The clinical application of MSCs is strictly regulated to ensure safety, quality, and efficacy. Regulatory frameworks differ across regions but generally emphasize minimal manipulation, homologous use, and compliance with Good Manufacturing Practice (GMP) standards. Furthermore, ethical concerns include informed consent, particularly regarding the experimental nature of MSC therapies, potential risks, and uncertain long-term outcomes. Transparency in reporting clinical results and adverse events is essential. There is also a need for standardization of protocols, including cell sourcing, dosing, delivery methods, and outcome measures, to improve reproducibility and comparability across studies 100 .
An important safety consideration in regenerative therapies for hair loss is the distinction between homologous and non-homologous use, particularly when comparing AD-MSCs and HFSCs. HFSCs, which are tissue-specific epithelial stem cells rather than MSCs, represent a homologous cell source for hair regeneration, as they inherently contribute to follicular cycling and hair shaft formation within their native niche83,84,142. In contrast, AD-MSCs are typically applied in a non-homologous manner, as their primary physiological role in adipose tissue does not involve hair follicle regeneration but rather paracrine support and tissue homeostasis116,117,142. Therefore, HFSC-based approaches may offer a more favorable safety profile due to their homologous use, whereas AD-MSC therapies require careful consideration of the additional risks associated with non-homologous regenerative strategies 102 .
Induced pluripotent stem cell–based therapies
iPSCs are a type of pluripotent stem cell derived from adult somatic cells, such as skin or blood cells, that have been reprogrammed to an embryonic-like pluripotent state, mimicking the properties of embryonic stem cells (ESCs) through the induction of specific genes and factors. iPSCs are similar to ESCs in many aspects, including morphology, proliferation, surface antigens, gene expression, epigenetic status of pluripotent cell-specific genes, and telomerase activity143–145. iPSCs enable the development of an unlimited source of any human cell needed for therapeutic purposes, and they were discovered by Shinya Yamanaka and his team in 2006 from mouse fibroblasts by introducing four factors, Oct3/4, Sox2, c-Myc, and Klf4, under ESCs culture conditions 144 . Later, human iPSCs were first independently produced by Yamanaka’s group through inducing Oct4, Sox2, Klf4, c-Myc (OSKM), and Thomson’s groups through inducing Oct4, Sox2, Lin28, and Nanog from human fibroblasts in 2007 145 , 146 . iPSCs possess three key properties that make them highly valuable for research and clinical applications. They can be derived from any individual’s somatic cells, enabling personalized approaches. They are pluripotent, capable of differentiating into all cell lineages of the three germ layers, such as ectoderm, endoderm, and mesoderm. Additionally, their ability to proliferate indefinitely provides a stable and renewable cell source for long-term and large-scale studies147,148.
The production process of iPSCs involves a sequence of highly controlled steps (Fig. 3): somatic cell isolation and selection, delivery of reprogramming factors, reprogramming culture, colony selection and expansion, and pluripotency validation 149 . The process begins with the selection and isolation of somatic cells, commonly dermal fibroblasts obtained through skin biopsy, mononuclear cells from peripheral (PBMCs) or other easily accessible sources, isolated in non-invasive or minimally invasive such as keratinocytes and urinary cells containing renal tubular epithelial cells and fibroblast-like or urothelial cells 150 . The choice of the somatic source may influence reprogramming efficiency, kinetics, and the epigenetic memory of the resulting iPSCs phenotype and transplantation outcome 151 . Somatic cell reprogramming begins with changes to the transcriptome and chromatin structure, shifting the cell from a differentiated to a pluripotent-like state. This process is primarily regulated by chromatin accessibility, which is influenced by DNA methylation, histone modifications, and ATP-dependent remodeling 147 . Reprogramming transcription factors known as the Yamanaka factors: OSKM bind to pluripotency-related DNA sequences and form an autoregulatory network that activates their own expression and that of other pluripotency-associated genes144,152.

Generation of iPSCs and application in hair follicle regeneration. iPSCs present a promising avenue due to three main features, including the ability to be produced from anyone’s cell, the ability to proliferate almost indefinitely, and the ability to be transformed into cells of various tissues and organs. iPSCs can be directed to differentiate into several key progenitor hair follicle cell types, such as dermal papilla cells, melanocytes, and keratinocytes. (Created by BioRender.com/Mahidol University.)
Delivery of these reprogramming genes can be conducted via various strategies, each with distinct advantages and limitations. iPSC reprogramming methods are categorized into integrative and nonintegrative, as well as viral and nonviral systems153,154. Integrative viral systems like retroviruses and lentiviruses are widely used for iPSC generation, though retroviruses often result in silenced transgene expression and impaired pluripotency due to epigenetic changes 155 . Lentiviruses offer higher efficiency and broader tropism through advanced polycistronic or intron-containing sequences 156 . Although safety has been improved using Cre/loxP and inducible systems, these methods remain limited by potential inefficiency and risks of genomic alterations157,158. Integrative nonviral methods, such as plasmids and transposons like piggyBac and Sleeping Beauty, have been developed to address the safety concerns of viral systems. Although they allow for transient expression and potential transgene excision, their low efficiency and risk of genomic disruption still limit their clinical applicability 159 . Nonintegrative viral vectors such as adenovirus and Sendai virus provide safer reprogramming options by avoiding genomic integration. While adenovirus has very low efficiency, Sendai virus offers improved gene delivery and therapeutic potential, though its reprogramming efficiency still requires further optimization160,161. Nonintegrative nonviral methods like episomal plasmids, minicircle DNA, and mRNA provide safer iPSC generation without genome integration, though their efficiency is generally lower than viral approaches. Emerging strategies using proteins, chemicals, and microRNAs show potential to enhance safety and efficiency, but still face technical challenges and require further optimization for reliable use 162 .
Following the delivery of reprogramming factors, transduced cells are cultured under conditions that support pluripotency. These include defined media such as mTeSR1 or Essential 8, in combination with xeno-free and feeder-free medium formulated for the growth and expansion of human pluripotent stem cells 149 . Over a period of 2–4 weeks, cells undergoing successful reprogramming begin to exhibit morphological changes, forming compact colonies with high nuclear-to-cytoplasmic ratios and prominent nucleoli, which are hallmarks of pluripotent cells 163 .
Once iPSC-like colonies are identified, they are manually or enzymatically picked and expanded under stringent conditions 164 . To confirm successful reprogramming, a series of characterization assays is employed. Expression of common pluripotency markers, such as OCT4, SOX2, and NANOG, is assessed via immunocytochemistry or qRT-PCR. The most common surface markers used for validation of iPSCs are SSEA-4, TRA-1-60, and TRA-1-81. SSEA-4 expression seems to precede the expression of TRA-1–60 and TRA-1–80, which are only detectable at later differentiation stages 165 . In addition, functional assays, including embryoid body formation, directed in vitro differentiation, and teratoma formation in immunodeficient mice, provide evidence of the cell line’s ability to differentiate into derivatives of all three germ layers166,167. Karyotyping and genome integrity analyses are also performed to ensure that the reprogramming process has not introduced harmful mutations or chromosomal abnormalities168,169.
iPSCs as a source of key hair follicular cell types and inducing hair follicle neogenesis in alopecia
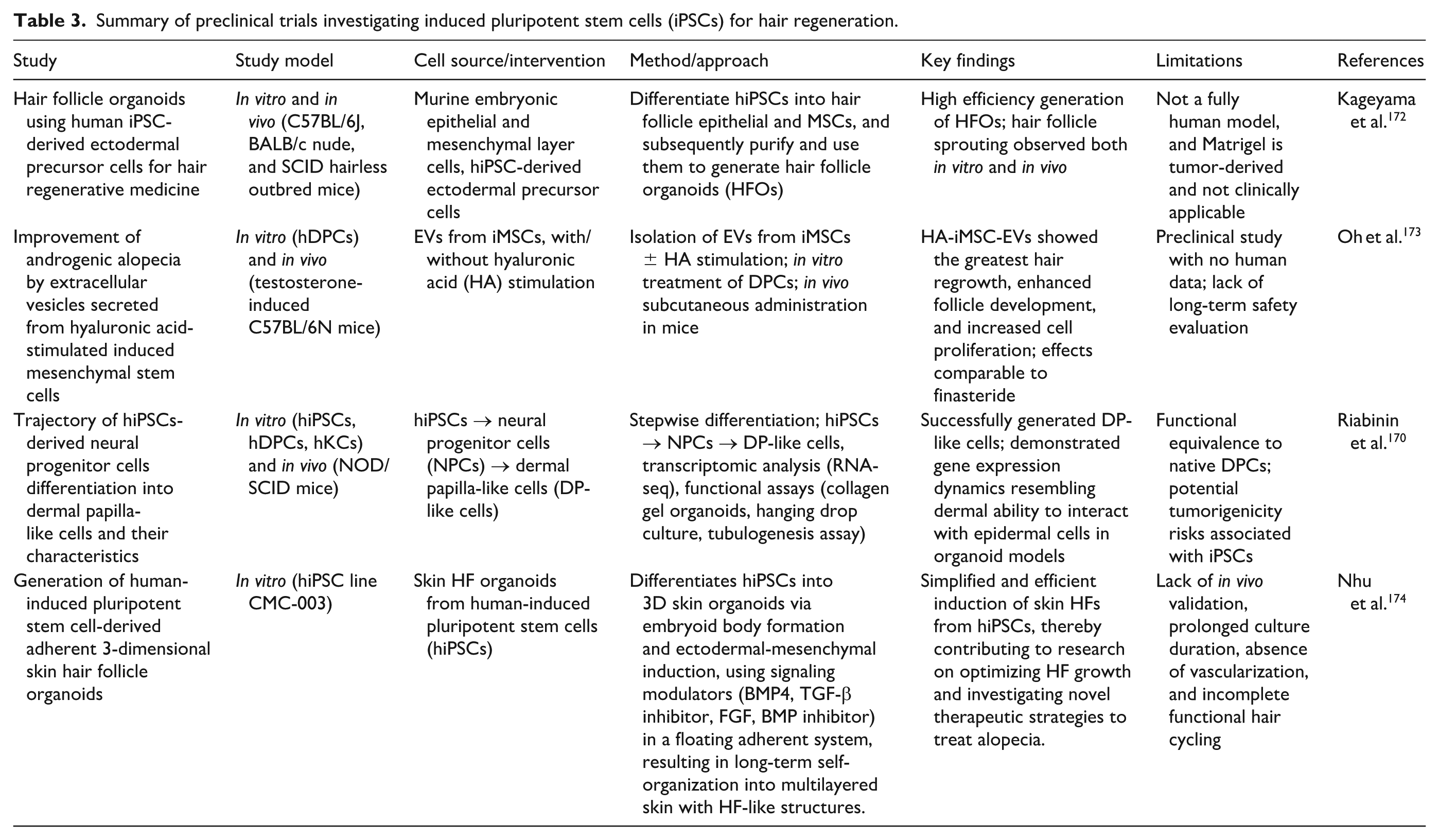
Hair follicle regeneration is a key focus in regenerative medicine, aiming to treat conditions like alopecia and age-related hair loss. iPSCs present a promising avenue by enabling personalized, scalable, and ethically acceptable therapies for hair restoration. iPSCs can be directed to differentiate into several key progenitor cell types, including those essential for the development and regeneration of hair follicles, such as DPCs, melanocytes, and keratinocytes (Fig. 3) 107 . iPSCs can give rise to DPCs, which are crucial for hair follicle development, growth, and cycling. They act as a signaling center, interacting with other cells in the follicle to regulate hair growth, shape, size, and even color 105 . Riabinin et al. 170 established a stepwise protocol to differentiate hiPSCs into dermal papilla-like cells through a neural progenitor cell (NPC) stage. The differentiated cells exhibited key dermal papilla markers, including versican (VCAN), α-smooth muscle actin (α-SMA), fibronectin (FN1), vimentin (VIM), and CD133, which also demonstrated functional hair-inductive properties in 3D epithelial–mesenchymal co-culture system 170 . Several studies have demonstrated that iPSC-derived epithelial and mesenchymal cells can reconstitute functional hair follicles when transplanted into animal models (Table 3). In particular, when iPSC-derived DP-like cells are combined with epidermal progenitors and implanted into the dermis of immunodeficient mice, they can induce de novo hair follicle neogenesis 171 .
Summary of preclinical trials investigating induced pluripotent stem cells (iPSCs) for hair regeneration.
iPSCs have the potential to generate melanocytes, the pigment-producing cells responsible for hair coloration, by synthesizing melanin within organelles called melanosomes and transferring these keratinocytes, epithelial stem cells, which iPSCs can also differentiate into keratinocytes that comprise the hair shaft itself, as well as the encircling inner and outer root sheaths 175 . Liu et al. (2019) demonstrated that human iPSC-derived melanocytes (hiMels) have long-term functionality in restoring pigmented hair follicles and establishing both mature melanocytes and stem cell populations, supporting their potential as a personalized therapy for hair and skin depigmentation. A modified in vivo hair follicle reconstitution assay showed that hiMels localized to the hair bulb and epidermis, producing melanin for up to 7 weeks post-transplantation 176 . Sah et al. 177 developed a feeder-free, efficient protocol to differentiate human iPSCs into keratinocyte progenitors in the presence of factors like retinoic acid and bone morphogenetic protein 4 (BMP4), followed by their maturation into keratinocytes upon calcium addition. The method offers a reliable source of functional keratinocytes for regenerative medicine and skin disease modeling 177 . These differentiation capabilities make iPSCs a versatile and promising source of multipotent progenitor cells for engineering fully functional hair follicles in regenerative medicine.
Safety concerns and regulatory considerations of iPSCs in hair regeneration
A primary safety concern associated with iPSCs is their inherent tumorigenic potential. Residual undifferentiated iPSCs within the transplanted cell population can give rise to teratoma formation, even when present in small quantities. The risk is further compounded by genetic abnormalities that may arise during the reprogramming process or be prolonged in vitro expansion 178 . The use of integrating viral vectors and oncogenic transcription factors, such as c-MYC, may also increase the likelihood of insertional mutagenesis and malignant transformation 179 . Insertional mutagenesis is a functional genomics approach that uses the integration of exogenous DNA elements, such as retroviruses or transposons, into the genome to identify cancer-related genes. The integration of the transgene into the host genome can disrupt tumor suppressor genes or activate proto-oncogenes, thereby increasing oncogenic potential. The risk is further compounded by the proliferative capacity and pluripotency of iPSCs, which make them more susceptible to tumor formation 180 . In addition to tumorigenicity, genomic instability and heterogeneity pose substantial risks. iPSCs can accumulate chromosomal aberrations and mutations over time, potentially altering their differentiation capacity and functional behavior. Incomplete or aberrant differentiation may lead to a mixed cell population or unintended lineage commitment, increasing the risk of ectopic tissue formation and reduced therapeutic efficacy 181 . Although autologous iPSC-derived cells are expected to minimize immune rejection, immunogenic responses have been reported due to altered antigen expression following reprogramming and differentiation 181 .
From a regulatory perspective, iPSC-based therapies are typically classified as advanced therapy medicinal products (ATMPs), as they involve more-than-minimal manipulation and complex manufacturing processes 182 . As such, they are subject to rigorous oversight by regulatory authorities. Compliance with the GMP standard is essential to ensure product consistency, sterility, and quality, with critical quality attributes including cell identity, purity, viability, and the absence of undifferentiated pluripotent cells. In particular, validated assays for tumorigenicity and genomic stability are required before clinical application 183 . Safety concerns, including tumorigenicity, genetic instability, and immunogenicity, further necessitate stringent quality control and thorough characterization of final cell products. Additionally, the elimination of undifferentiated cells and the maintenance of stable functional phenotypes remain critical for ensuring clinical safety. Beyond these technical barriers, high production costs and complex supply chains continue to limit large-scale implementation and commercialization181,184.
Recent advances in stem cell–based regenerative procedures for hair regeneration
Tissue engineering and bioprinting
Recent advances in tissue engineering and 3D bioprinting have opened new frontiers for hair follicle regeneration. Using iPSC-derived cells, researchers are now able to fabricate skin-like constructs embedded with functional hair follicles. Castro and Logarinho 185 reviewed tissue engineering approaches for regenerating human hair follicles, highlighting key challenges such as maintaining the hair-inducing capacity of DPCs, sourcing competent epithelial stem cells, and replicating the complex follicle microenvironment. Reciprocal epithelial-mesenchymal (EM) interactions that regulate embryonic organogenesis and postnatal hair follicle cyclic growth are the foundation of hair follicle bioengineering techniques 185 . Lee et al. 186 developed a 3D organoid system using mouse pluripotent stem cells that self-organize into skin-like structures capable of forming hair follicles. The organoids recapitulate key stages of embryonic hair follicle development, including placode formation, dermal condensate aggregation, and early follicle morphogenesis. Molecular and structural analyses confirmed similarities to in vivo development. This model offers a valuable platform for studying hair follicle biology, disease modeling, and regenerative medicine, though full follicle maturation and cycling remain challenges for future work73,186.
The effective hair follicle neogenesis requires not only iPSCs but also supportive mesenchymal components and a biomimetic microenvironment. MSCs enhance regeneration through paracrine signaling, immunomodulation, and secretion of bioactive factors123,126. Incorporation into 3D scaffolds further improves outcomes by mimicking the extracellular matrix and promoting essential epithelial-mesenchymal interactions 122 . Supporting this concept, a recent study on the synergistic effects of mesenchymal stem cell secretome and Wnt10b on hair follicle regeneration in a 3D bioengineering model demonstrated that a 3D GelMA hydrogel system combining DPCs and epithelial cells with placenta-derived MSC secretome significantly enhanced hair follicle formation both ex vivo and in vivo. This effect is amplified by Wnt/β-catenin activation, particularly by Wnt10b, which promotes follicular induction and anagen transition 187 .
Stem cell–derived extracellular vesicles or exosomes
Extracellular vesicles (EVs) are nanobiological membrane-bound vesicles released by most cell types into the extracellular space of body fluids and cell culture media, which is the most common intercellular communication pathway188,189. The content, or cargo, of EVs consists of proteins, lipids, and nucleic acids, which influence numerous signaling pathways in cellular communication190,191. EVs are categorized into small EVs and large EVs based on their size, molecular composition, and subcellular origins 192 . Based on the diameter of the separated particles, small EVs are often described as <200 nm in diameter, including exosomes and microvesicles (ectosome), and large EVs are often described as >200 nm, such as apoptotic bodies and microvesicles193,194. EVs are emerging as critical mediators of intercellular communication and harbor potent therapeutic potential, thereby underscoring their significance in facilitating the rapid transition toward cell-free therapeutic strategies in hair regeneration research 195 . EVs, particularly exosomes derived from MSCs and DPCs, encapsulate a rich cargo of bioactive molecules (proteins, growth factors, and microRNAs) that stimulate dormant hair follicles and modulate the hair growth cycle188,196.
Current research is focused on characterizing the regenerative potential of EVs, with MSC-derived exosomes, sourced from various tissues, including adipose tissue, placenta, hair follicles, bone marrow, foreskin, and umbilical cord, which have shown promising results in treating hair loss, primarily in AGA. Across the studies analyzed, all demonstrated improvements in at least one hair parameter. Notably, these exosome treatments led to substantial increases in both hair density, ranging from 9.5 to 35 hairs/cm2, and hair thickness, with gains reported up to 13.01 µm, and no significant side effects were observed 197 . The retrospective study by Sasaki 198 used a single injection of BM-MSC EVs (XoFlo) in 31 patients with hair loss, which led to an 11.1%–24.2% increase in hair density over 6 months. The treatment was considered safe, with no severe adverse events, and resulted in high patient satisfaction 198 . Furthermore, the study shows that EVs derived from DPCs carry the essential instructions to promote and regulate the hair growth cycle, directly activating HFSCs and promoting proliferation by initiating the critical Wnt/β-catenin signaling pathway via transferred molecules such as Wnt3a and miRNAs73,199. They can reprogram other stem cells (like adipose-derived stem cells) to acquire dermal papilla-like properties, providing a potential source of functional, hair-inducing cells for therapy 196 .
The multifaceted role of EVs in hair follicle regeneration
Delivery of bioactive cargo and hair cycle modulation
Exosomes are critical mediators of cell-to-cell signaling, transporting bioactive molecules to their target sites via mechanisms such as membrane fusion and endocytosis (Fig. 4). This delivery system consequently potentiates intracellular signaling and facilitates the transfer of substances across cell membranes 200 . The roles of EVs, particularly exosomes, in hair regeneration center on their function as biological messengers that coordinate the hair growth cycle and optimize the scalp microenvironment 197 . They effectively transport and deliver a repertoire of essential growth factors (GFs) such as PDGF, VEGF, FGF, KGF, HGF, and IGF-1. These GFs directly stimulate the proliferation and migration of both DPCs and ORSCs122,188. Finally, exosomes significantly improve the scalp microenvironment by promoting angiogenesis (neovascularization) via VEGF delivery, thereby ensuring optimal blood flow and nutrient supply to the follicles201,202. Exosomes derived from regenerative cell sources, such as DPCs and MSCs, are critical for accelerating the critical shift from the quiescent telogen phase to the active anagen phase. Rajendran et al. 188 indicated that MSC-EVs can induce earlier conversion of the hair cycle and stimulate hair growth in a murine model. The quantified results revealed that the MSC-EVs groups showed a significant increase (P > 0.05) in the hair growth, and a higher increase (P > 0.001) was measured at day 27. Sections of the dorsal skin after 27 days demonstrated that the application of MSC-EVs significantly promoted conversion from telogen to anagen compared to the control group, suggesting that MSC-EVs promote hair regrowth by activating conversion from telogen to anagen of hair follicles 188 .

Extracellular vesicle roles in hair regeneration. EVs, mainly exosomes from dermal papilla or mesenchymal stem cells, act as messengers delivering a cargo of growth factors and regulatory molecules. These EVs trigger hair regeneration by activating the signaling pathway in hair follicle stem cells, thereby promoting cell proliferation and creating a favorable regenerative environment, and promoting an anti-inflammatory role, primarily from MSC-EVs. EVs deliver cargo (IL-10) that reprograms the local immune environment, shifting it from pro-inflammatory to anti-inflammatory. (Created by BioRender.com/Mahidol University.)
Activation of regenerative signaling pathways
Exosomal delivery is essential for activating key molecular pathways central to hair follicle development and homeostasis. Hair regeneration is fundamentally controlled by a dynamic balance between activating and inhibitory signals, including Wnt/β-catenin, Shh, Notch, and BMP signaling 88 . Zhou et al. 203 provided evidence that DPCs regulate hair growth by releasing exosomes, which stimulate ORSCs proliferation and migration via β-catenin and Shh signaling, and accelerate the onset of hair follicle anagen and delay catagen in mice. Furthermore, the study of Shang et al. 204 showed that treatment with dermal papilla cell-derived exosomes (DPC-Exos) increases the expression levels of key Wnt/β-catenin pathway transcription factors (β-catenin and Lef1) in fibroblasts and in the dermis of skin wounds, and also induces hair follicle neogenesis in nude mice when combined with neonatal mice epidermal cells. The transfer of specific microRNAs and regulatory proteins via exosomes directly targets and represses the expression of the bone morphogenetic protein 2 (BMP2). The study from Chen et al. 205 found that DP-EVs (from low-passage, P3) are highly enriched with a specific microRNA called miR-140-5p, which is delivered to recipient hair follicle cells. By downregulating BMP2, the DP-EVs effectively release the inhibitory brake on the hair cycle, allowing HFSCs and matrix cells to increase their proliferation, thus stimulating hair growth and accelerating hair follicle elongation 205 .
Immunomodulatory and anti-inflammatory mechanisms
The progressive miniaturization and subsequent loss of the hair follicle, characteristic of conditions like AGA and AA, are critically linked to chronic, low-grade perifollicular inflammation within the scalp microenvironment206,207. EVs, particularly MSC-derived exosomes, represent a promising therapeutic frontier due to their anti-inflammatory and immune-regulating effects, anti-aging properties, and they promote wound healing, stimulate hair growth, and repair the skin barrier 208 . MSC-EVs facilitate a critical shift in the immune microenvironment by promoting the polarization of macrophages from the pro-inflammatory M1 phenotype toward the restorative M2 phenotype 209 . The M1 phenotype is associated with chronic inflammation and tissue damage, and its promotion is essential, particularly in mitigating conditions such as stress-induced hair loss via IL-18 and IL-1β 131 . By contrast, the induced M2 macrophages play a beneficial role in tissue homeostasis and repair. These cells secrete vascular growth factors such as VEGF, IGF-1, TGF-β, TNF-α, IL-6, IL-8, and IL-10 210 .
Conclusion
In conclusion, the investigation into stem cell-based treatments obviously establishes them as the most compelling and potentially transformative approach to addressing extensive hair loss conditions, notably AGA. This review has demonstrated the theoretical and preclinical power of utilizing various stem cell lineages, particularly HFSCs, MSCs, iPSCs, and their derivatives, including extracellular vesicles, which serve as the biological agents capable of modulating the hair cycle and primarily focusing on the hair follicle regenerative capacity, thus highlighting their substantial role in follicular neogenesis. Furthermore, extracellular vesicles, particularly exosomes derived from these stem cells, are established as the principal mediators of the paracrine effect. They encapsulate and deliver a concentrated bioactive cargo that directly regulates critical signaling pathways, promoting angiogenesis and prolonging the anagen phase in neighboring follicular cells. The shift toward EV-based, cell-free therapy offers significant advantages over direct cell transplantation, including lower immunogenicity, easier scalability, and enhanced stability.
Despite the highly encouraging preclinical and early-phase clinical data, the clinical translation of these modalities is currently restricted by several critical challenges, including a significant lack of standardized protocols for the isolation and characterization of therapeutic, the challenge of establishing GMP-compliant, large-scale production, and the need for optimized dosing and delivery methods. Therefore, the successful translation of stem cell-based therapies will depend on the integration of robust manufacturing platforms, comprehensive safety assessments, and strict regulatory compliance, supported by coordinated efforts across scientific, industrial, and regulatory domains.
The future perspective of hair follicle regeneration is therefore centered on innovation in bioengineering. Efforts are concentrating on developing complex 3D culture systems and biodegradable scaffolds that successfully mimic the in vivo niche, thereby preserving the critical signaling properties of the stem cells. As regulatory pathways mature and techniques such as microinjection and cell delivery become standardized, these therapies will inevitably become scalable and accessible. Ultimately, stem cell therapy is not merely an improvement upon current surgical and pharmacological options. It is the inevitable direction of regenerative medicine in dermatology, poised to revolutionize how we understand and treat the weakening effects of hair loss.
Footnotes
Acknowledgements
This research project was partially supported by Scholarships for PhD Student from Mahidol University. This research project is supported by Mahidol University (Fundamental Fund: fiscal year 2023 by National Science Research and Innovation Fund (NSRF)).
Ethical considerations
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Author contributions
Pitsunan Paphawannasri: Writing – original draft, Visualization. Tulyapruek Tawonsawatruk: Writing – review and editing. Jitrada Phetfong: Writing – review and editing. Aungkura Supokawej: Conceptualization, Writing – review and editing, Supervision.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research project was partially supported by Scholarships for PhD Student from Mahidol University. This research project is supported by Mahidol University (Fundamental Fund: fiscal year 2023 by National Science Research and Innovation Fund (NSRF)).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
Not applicable.
Statement of human and animal rights
This article does not contain any studies with human or animal subjects.
Statement of informed consent
There are no human subjects in this article and informed consent is not applicable.